
Abstract
Therapeutic genome editing is currently reshaping and transforming the development of advanced therapies as more ex vivo and in vivo gene editing-based technologies are used to treat a broad range of debilitating and complex disorders. With first-generation gene editing modalities (notably those based on ZFNs, TALENs and CRISPR/Cas9), comes a new second-generation of gene editing-based therapeutics including base editing, prime editing and other nuclease-free genome editing modalities. Such ground-breaking innovative products warrant careful considerations from a product development and regulatory perspective, that take into account not only the common development considerations that apply to standard gene and cell therapy products, but also other specific considerations linked with the technology being used. This article sheds light into specific considerations for developing safe and effective in vivo and ex vivo genome editing medicines that will continue to push barriers even further for the cell and gene therapy field.
The past 10
Molecular tools and modes of therapeutic editing for current and second-generation genome editing technologies
Listed examples of target clinical indications in development as of November 30, 2022.
Terminated (NCT02695160; NCT02702115; NCT03041324).
AAV, adeno-associated virus; ABEs, adenine base editors; ADARs, adenosine deaminases acting on RNA; APOBEC, apolipoprotein B mRNA editing enzyme, catalytic polypeptide-like; A-to-G, adenine-to-guanine; A-to-I, adenosine-to-inosine; CBEs, cytidine base editors; CRISPR-Cas9, clustered regularly interspaced short palindromic repeat-associated Cas9 nuclease; C-to-T, cytosine-to-thymine; C-to-U, cytidine-to-uracil; G-to-A, guanine-to-adenine; MGN, meganuclease; mRNA, messenger RNA; pegRNA, prime editing guide RNA; TALEN, transcription activator-like effector nuclease; T-to-C, thymine-to-cytosine; ZFN, zinc finger nuclease.
This was soon followed by the first in vivo adeno-associated virus-based ZFN-mediated gene editing product for lysosomal storage diseases to be used in the clinic in 20183 and the first in vivo nonviral-based clustered regularly interspaced short palindromic repeat-associated Cas9 nuclease (CRISPR/Cas9)-mediated gene editing product for transthyretin amyloidosis. 4 Beyond cancer, lysosomal storage diseases and pathologies caused by accumulation of misfolded proteins, ex vivo genome editing using CRISPR/Cas9 has shown remarkable clinical results in hemoglobinopathies, which could pave the way to the anticipated marketing approval of the first genome editing product for sickle-cell disease in 2023. 5
With the first-generation nuclease-dependent gene editing technologies (ZFNs, TALENs, meganucleases, and CRISPR/Cas9) comes a second-generation of gene editing-based therapeutics notably base editing (cytidine base editors and adenine base editors) and prime editing; as well as other nuclease-free genome editing modalities and epigenetic editing (the tuning of gene outputs) (Table 1; Fig. 1).
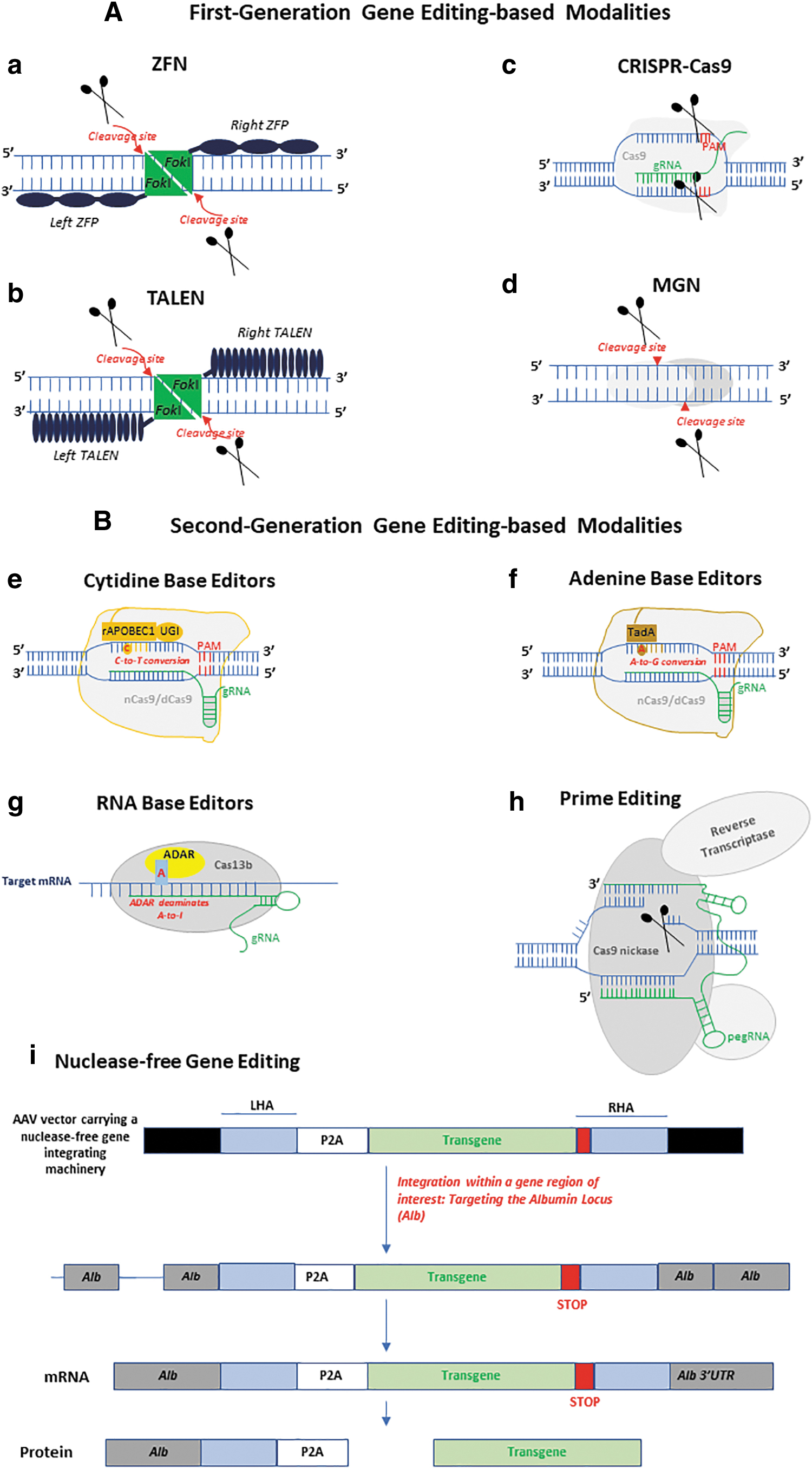
Schematic representation of first- and second-generation gene editing-based approaches. First-generation gene editing-based modalities
The next 12 months may be groundbreaking for the field, with the first ex vivo CRISPR-based gene editing product for sickle-cell disease anticipated to reach the market in 20235; and initial clinical data arising from the first pivotal registration-enabling Phase 2 study using allogeneic TALEN-based gene-edited anti-CD19 CAR-T cells for relapsed/refractory non-Hodgkin lymphoma. 6
As new second-generation gene editing systems move toward first-in-human studies, a shift toward using such technologies for in vivo administration and in a much younger target population to address specific unmet medical needs will likely increase. This will potentially push barriers even further, paving the way to potentially more complex applications notably xenotransplantation 7 and possibly other unprecedented therapeutic interventions.
Nevertheless, such groundbreaking applications warrant careful considerations from a technical scientific, clinical, and regulatory perspective, backed up with a robust risk-based science-driven approach to drive the development of such sophisticated products.
From a product development perspective, these novel technologies share some common technical considerations as those that typically apply to standard viral vectors, nonviral systems (such as lipid nanoparticles), and cell-based products. Other specific considerations are linked to the process itself and any other product-specific characteristics associated with the technology being used. In the case of gene-edited cell-based products, these, for instance, include the number of gene editing events the processed cells have been subjected to, the conditions under which the gene editing was conducted (including the quality of the genome editing components/materials used) as well as the cell banking system used to generate the final cell product.
For products with a “high-risk” profile, and where limited safety information can be gathered from preclinical animal models, a thorough product characterization to evaluate off-target gene editing effects, based on an orthogonal testing approach and latest state of the art sensitive methods, encompassing at least one unbiased genome-wide analysis, should be considered. Table 2 highlights some examples of unbiased analytical assays used to detect off-target and unwanted on-target gene editing events.
Some examples of unbiased methods used for the detection of off-target and unwanted on-target gene editing events
AMP, anchored multiplex PCR sequencing; BLESS, direct in situ breaks labeling, enrichment on streptavidin and next-generation sequencing; CHANGE-Seq, ccircularization for high-throughput analysis of nuclease genome-wide effects by sequencing; ChIP-Seq, chromatin immunoprecipitation sequencing; CIRCLE-Seq, circularization for in vitro reporting of cleavage effects by sequencing; DIG-Seq, digenome-seq using cell-free chromatin DNA; Digenome-Seq, in vitro nuclease-digested genome sequencing; DISCOVER-Seq, discovery of in situ Cas off-targets and verification by sequencing; DSB, double-strand break; dsODN, oligodeoxynucleotide; GE, gene editing; GUIDE-Seq, genome-wide, unbiased identification of DSBs enabled by sequencing; HTGTS, high-throughput genome-wide translocation sequencing; IDAA, indel detection by amplicon analysis; IDLV, integrase-defective lentiviral vector; iPSCs, induced pluripotent stem cells; LAM-PCR HTGTS, linear-amplification-mediated-polymerase chain reaction high-throughput genome-wide translocation sequencing; MRE11, meiotic recombination 11 homolog 1; MRN complex, MRE11-RAD50-NBS1 complex; NHEJ, nonhomologous end joining; SITE-Seq, selective enrichment and identification of tagged genomic DNA ends by sequencing; sgRNA, single-guide RNA; VIVO, verification of in vivo off-targets; WGS, whole-genome sequencing.
For in vivo genome editing products, an additional layer of safety applies, notably with regard to products developed for systemic administration, where the risk of uptake into nontarget organs/tissues (including germline cells) should be considered and addressed to rule out any risk of accidental germline modification/transmission.
Although unbiased testing methods are proving beneficial from a product de-risking point of view, one has to account for the “human factor” and target population. The impact of genetic variation/diversity on on-target but also off-target gene editing, within the target patient population should be accounted for, in light of recent evidence suggesting possible off-target gene editing effects linked with genetic variants, which can potentially predispose certain individuals to treatment failures or even potential adverse events. 27
The impact of ethnic genetic variations on off-target gene editing mediated by therapeutic guide RNAs that have been used as part of an ex vivo CRISPR-Cas9-gene editing approach for sickle-cell disease and other genetic disorders (notably severe combined immunodeficiency, chronic granulomatous disease, X-linked hyper IgM syndrome, acquired immunodeficiency syndrome, and Leber congenital amaurosis) have been investigated. The findings revealed an additional 234 off-targets observed when variants from different ethnic groups were introduced.
These additional off-targets were not seen when the reference genome (hg38) was analyzed. 28 These results correlate with previous reports identifying those of African ancestry heritage as being the most heterogeneous for single nucleotide polymorphisms and structural variants, 29,30 which calls for caution particularly in those disease areas with a high prevalence in specific subsets of the population. Hence the importance of risk assessing the impact of such variations within the target disease population, when considering treatment with genome editing.
With the majority of conditions affecting the very young patient population, genome editing can potentially transform the way some debilitating notably orphan or ultra-orphan genetic diseases (for which no current or insufficient standard of care treatment is available) are treated. This could potentially offer a one-off single treatment approach that could be initiated at an early age, potentially resulting in a stable permanent therapeutic transgene expression over time as compared with conventional complementary DNA-based/nonintegrating gene therapies, which (depending on the organ being targeted and the age of patients being treated) carry the risk of transgene dilution over time and hence subsequent loss of optimal therapeutic activity.
Although therapeutic genome editing is still at its early clinical infancy, caution should be exercised, when it comes to evaluating the potential impact of on-target and off-target gene editing and the clinical significance of these. For this, collective efforts should be made to collect and understand the clinical significance of data that have been acquired/are to be acquired from adults and adolescents who have already received/will be receiving gene therapy products incorporating genome editing components, before infants, toddlers, and children are to be treated. These cumulative data are crucial to push the therapeutic genome editing field toward reaching its optimal potential in addressing unmet medical needs, including the possibility to safely intervene and treat at a much early disease onset.
As the field advances, and as we gain more direct experience correlating product-specific characteristics with clinical outcome (for ex vivo modified gene-edited cellular products), as well as understanding the long-term clinical significance/impact of in vivo genome editing activity from liver-targeted viral and lipid nanoparticle-based in vivo genome editing products, evidence-based and platform-based learning should be leveraged to continually tailor product-related quality and safety attributes to achieve the optimal benefit to risk ratio for the target disease population being treated.
Although such learning might provide some initial answers from a technical scientific and preliminary clinical safety perspective, more efforts are needed to develop sophisticated viral and nonviral vectors that are capable of transferring the gene editing machinery specifically to target tissues of interest, after systemic administration, to widen the therapeutic application of genome editing beyond liver-targeted indications for in vivo genome editing-based products.
Footnotes
AUTHOR DISCLOSURE
The author is an employee of Precision for Medicine and does not have any conflict of interest to declare. Precision for Medicine is a global precision medicine clinical research services organization, purpose-built to improve the clinical research and development process of new therapeutics. Precision for Medicine also provides consultancy in the field of cell and gene therapies.
FUNDING INFORMATION
No funding was received for this article.