
Abstract
The cardiac autonomic nervous system (CANS) is intimately connected to the regulation of electrophysiology and arrhythmogenesis in cardiac systems. This work aimed at investigating whether interleukin-10 (IL-10) could effectively modulate CANS and suppress ischemia-induced ventricular arrhythmia (VA) through chronically acting on the cardiac sympathetic ganglion (CSG). Using an adeno-associated virus (AAV), we achieved local chronic overproduction of IL-10 in the CSG, left stellate ganglion (LSG). As a result, in the IL-10 group, we observed a decreased number of tyrosine hydroxylase-positive (TH+) cells in the LSG. IL-10 markedly downregulated the nerve growth factor, synaptophysin, as well as growth-associated protein 43 expression. In vivo, results from ambulatory electrocardiography showed that IL-10 overexpression significantly inhibited the cardiac sympathetic nervous system activity and improved heart rate variability. Meanwhile, we observed decreased LSG function as well as prolonged ventricular effective refractory period and suppressed VA after myocardial infarction (MI) in the IL-10 group. In addition, IL-10 overexpression attenuated inflammation and decreased norepinephrine levels in the myocardium after acute MI. In conclusion, our data suggest that chronic IL-10 overexpression modulates cardiac sympathetic nerve remodeling and suppresses VA induced by MI. Neuromodulation through AAV-mediated IL-10 overexpression may have the characteristics of and advantages as a potential neuroimmunotherapy for preventing MI-induced VAs.
INTRODUCTION
Ventricular arrhythmia (VA) is the main cause of sudden cardiac death (SCD) secondary to myocardial infarction (MI). It is, and will remain, a critical international health issue. 1 Because cardiac autonomic nervous system (CANS) dysfunction is closely associated with arrhythmia, 2 rebalancing the CANS is considered an important therapeutic approach to reduce the burden of SCD in patients with MI. Studies have shown that the overexpression of numerous neuroimmune factors, especially pro-inflammatory cytokines, in the left stellate ganglion (LSG) microenvironment could activate CANS, resulting in the initiation and maintenance of ventricular arrythmia. 3
Moreover, contrary to that observed in pro-inflammatory signaling, some studies have demonstrated that attenuating neuroinflammation in the cardiac sympathetic ganglion (CSG) could regulate neuroimmune homeostasis and suppress cardiac arrhythmias. 4 Therefore, CSG microenvironment-based neuro-immune therapy is a feasible method for VAs treatment. The effective modulation of neurogenic inflammation, and neuronal excitability has become a key challenge in neuroimmunotherapy.
Interleukin-10 (IL-10), originally discovered as an inhibiting cytokine produced by T helper 2 cells, is an important anti-inflammatory cytokine. 5 In vivo, it plays an essential role in controlling inflammation. 6 Aside from its role in regulating inflammation, IL-10 has been described to play a protective role in neurodegenerative diseases, infection, and cancer. 7,8 IL-10 has emerged as an attractive therapeutic target in neuroimmune-related diseases.
Previous studies have demonstrated that IL-10 can limit neuronal damage during inflammatory processes as well as modulate homeostatic processes. 9 Generally, IL-10-based neuroimmune related therapies have shown a fair curative effect on both pathology and symptoms relevant to many neuroimmune diseases. 10,11 Therefore, exploring potential treatment strategies for IL-10-based therapy for neuroimmune-related diseases has been proposed in recent years. 12
Cardiovascular diseases, including but not limited to Vas, are associated with neuroimmune dysfunction. Because sympathetic activation is closely linked to VAs, and it is well known that IL-10 has the potential to modulate neuroimmune homeostasis, it is plausible to consider modulating the CANS through IL-10 as a potential therapeutic approach for VAs.
In this study, we used adeno-associated virus (AAV) to express IL-10 in the sympathetic ganglion, the LSG, in a canine model. We investigated whether IL-10 overproduction could play a cardioprotective role in preventing VAs through modulating the CSG.
MATERIALS AND METHODS
Animal preparation
All the experimental male beagles (body weight ranging from 8 to 12 kg) were obtained from Marshall (Beijing) and were housed at the Center of Experimental Animals, Renmin Hospital, Wuhan University. The Animal Care and Use Committees of Renmin Hospital approved all animal experiments (Approval No. WDRM20210909), and the research was conducted following the criteria of the National Institutes of Health. To implement animal anesthesia according to experimental requirements, 3% pentobarbital sodium was used, and under anesthesia, a ventilator was employed to assist in breathing (with an oxygen flow rate 4–6 L/min and a tidal volume 20 mL/kg).
An intravenous indwelling needle was inserted to establish a channel for rehydration, and arterial pressure was monitored by catheterization of the left femoral artery. Lead 7000, a computer-based laboratory system provided by Jinjiang Electronics (Chengdu, China), was used to collect data on blood pressure and body surface electrocardiography.
Experimental protocol
Eighteen experimental beagles were randomly allocated into three groups: the Control group (n = 6, receiving a microinjection of the same amount of saline + acute myocardial infarction [AMI]), the AAV group (n = 6, receiving a microinjection of empty AAV vector + AMI), and the IL-10 group (n = 6, receiving a microinjection of AAV vector with IL-10 microinjection + AMI).
Ambulatory electrocardiography (AECG) was recorded for all dogs to assess CANS activity. AAV vector (10 μL) or the same amount of saline was microinjected into the LSG. Thirty days later, AECG was recorded again. To evaluate the LSG function and detect ventricular electrophysiological parameters, a left thoracotomy was performed under anesthesia in all experimental animals. Subsequently, an acute ischemia model was established, and VAs events were recorded over 60 min. Finally, samples of both LSG and myocardial tissue were collected for further analysis (Supplementary Fig. S1A).
Statistical analysis
All data presented as the mean ± standard deviation (SD). One-way analysis of variance (ANOVA) was used to detect group differences. Two-way repeated-measures ANOVA was used to test the effect of LSG micro-injection on the CANS. Tukey's or Sidak's multiple-comparison test was used as a post hoc test to compare differences between individual groups. Unpaired Student's t-test was employed to analyze the differences in enhanced green fluorescence protein (EGFP) fluorescence intensity in the AAV and IL-10 groups. Data analysis was performed using GraphPad Prism software 9.1 (GraphPad Software, La Jolla, CA, USA). p < 0.05 was considered statistically significant.
For additional details and information on materials and methods, please refer to the Supplementary Data S1.
RESULTS
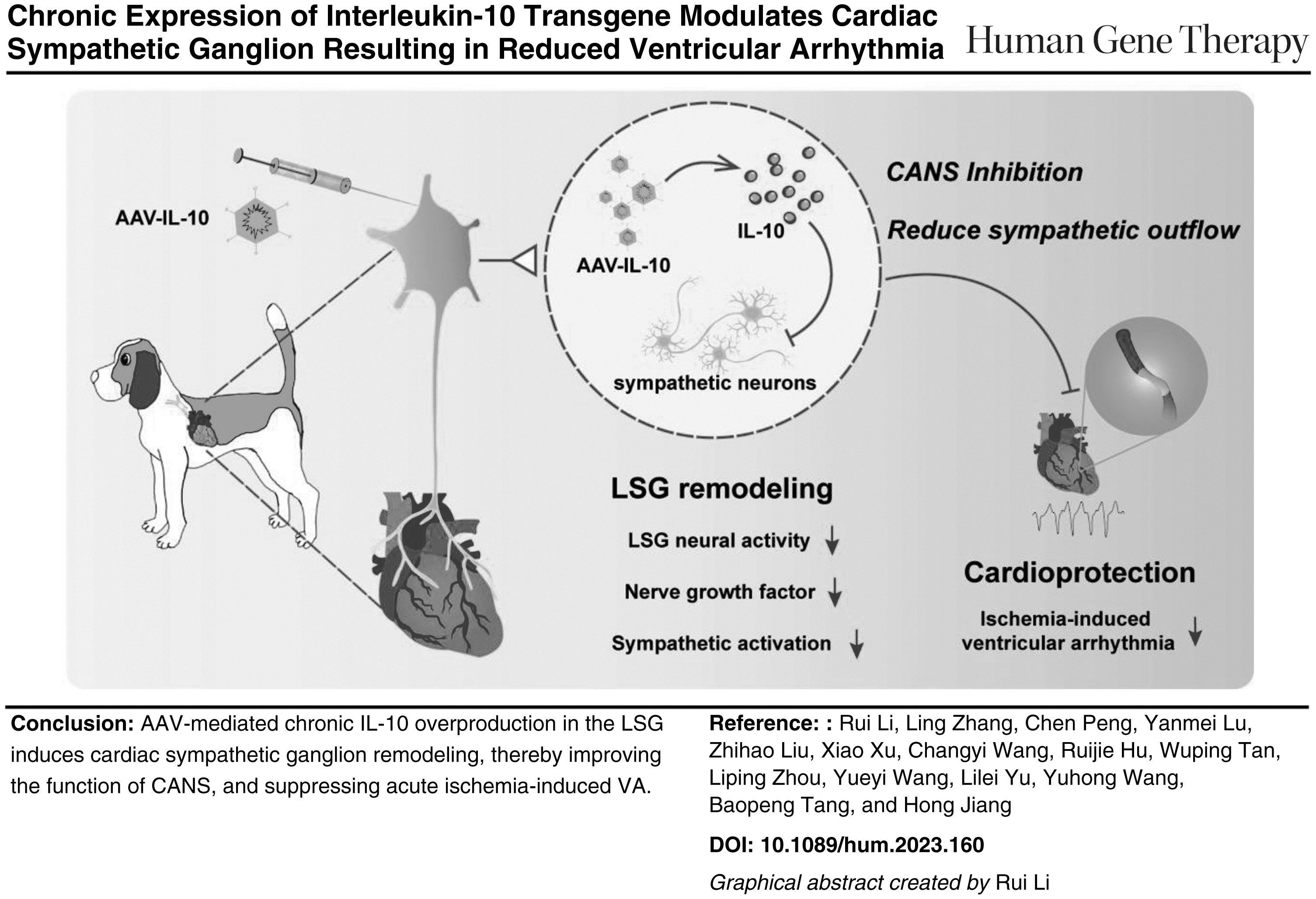
AAV-IL-10 microinjection into LSG leads to an increase in the local IL-10 expression
Immunofluorescence results of EGFP verified the successful transduction of the AAV vector into the LSG (Fig. 1A), and the quantitative results showed no difference between the AAV and IL-10 groups (Supplementary Fig. S1B, C). Subsequently, we measured the local IL-10 expression levels in all groups. IL-10 mRNA expression levels in the IL-10 group were significantly increased (Fig. 1B), whereas the other two groups exhibited similar expression levels.
IL-10 chronic overexpression in LSG tissue.
Immunofluorescent results demonstrated much stronger IL-10 immunoreactive staining in the IL-10 group, indicating an overproduction of IL-10 in the LSG (Fig. 1C, D). The IL-10 expression level in the control and the AAV groups in the LSG was comparable. Despite IL-10 overproduction within the LSG in the IL-10 group, no increase in serum levels was detected as compared with that in the other two groups (Supplementary Fig. S1D, E).
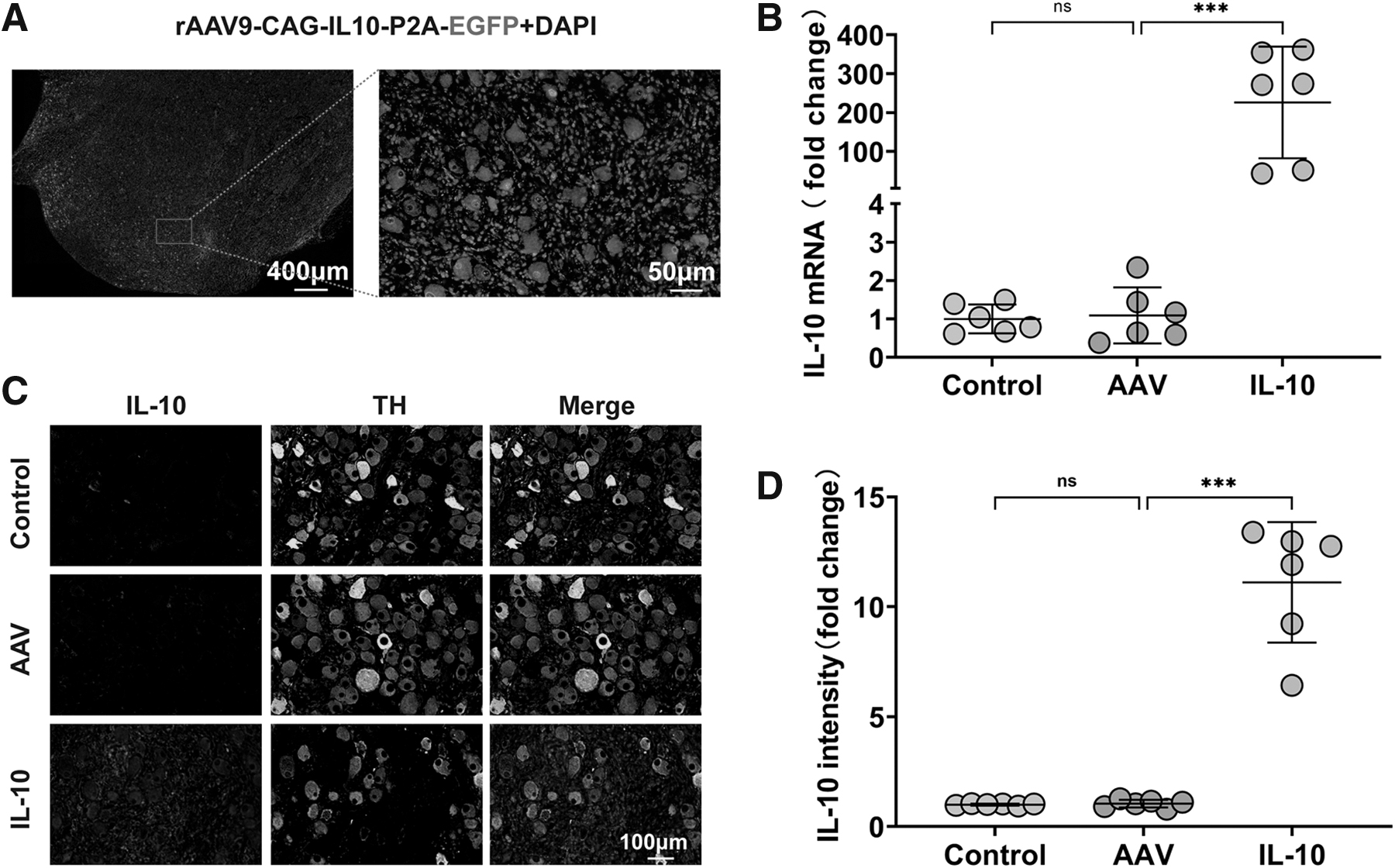
IL-10 overexpression mediated the modification of the sympathetic nervous system
Chronic IL-10 overproduction through AAV vector delivery caused modification of the cardiac sympathetic nerves. Immunofluorescence staining revealed a decrease in the number of TH-positive neurons in the IL-10 group compared with the AAV group, as well as a reduction in the NGF fluorescence intensity (Fig. 2A–C), indicating the inhibition of the cardiac sympathetic nervous system (SNS). Further, the distinct changes in numbers were accompanied by a decrease in SYN and GAP43 expression in the IL-10 group (Fig. 2D–F).
Effects of IL-10 overexpression on sympathetic modification in the LSG.
Immunoreactivity in the control and AAV groups showed no differences. In addition, we evaluated the expression of STAT3 (Fig. 2G) and DDIT4 (Fig. 2H), and the results showed that their expression levels were significantly up-regulated in the IL-10 group. Meanwhile, mammalian targets of rapamycin (mTOR) expression levels were decreased (Fig. 2I). These results suggest that IL-10 suppresses mTOR activity through the IL-10-DDIT4-mTOR axis. Consequently, IL-10 overexpression-induced stimulation of apoptosis and autophagy may have caused the inhibition of the cardiac SNS.
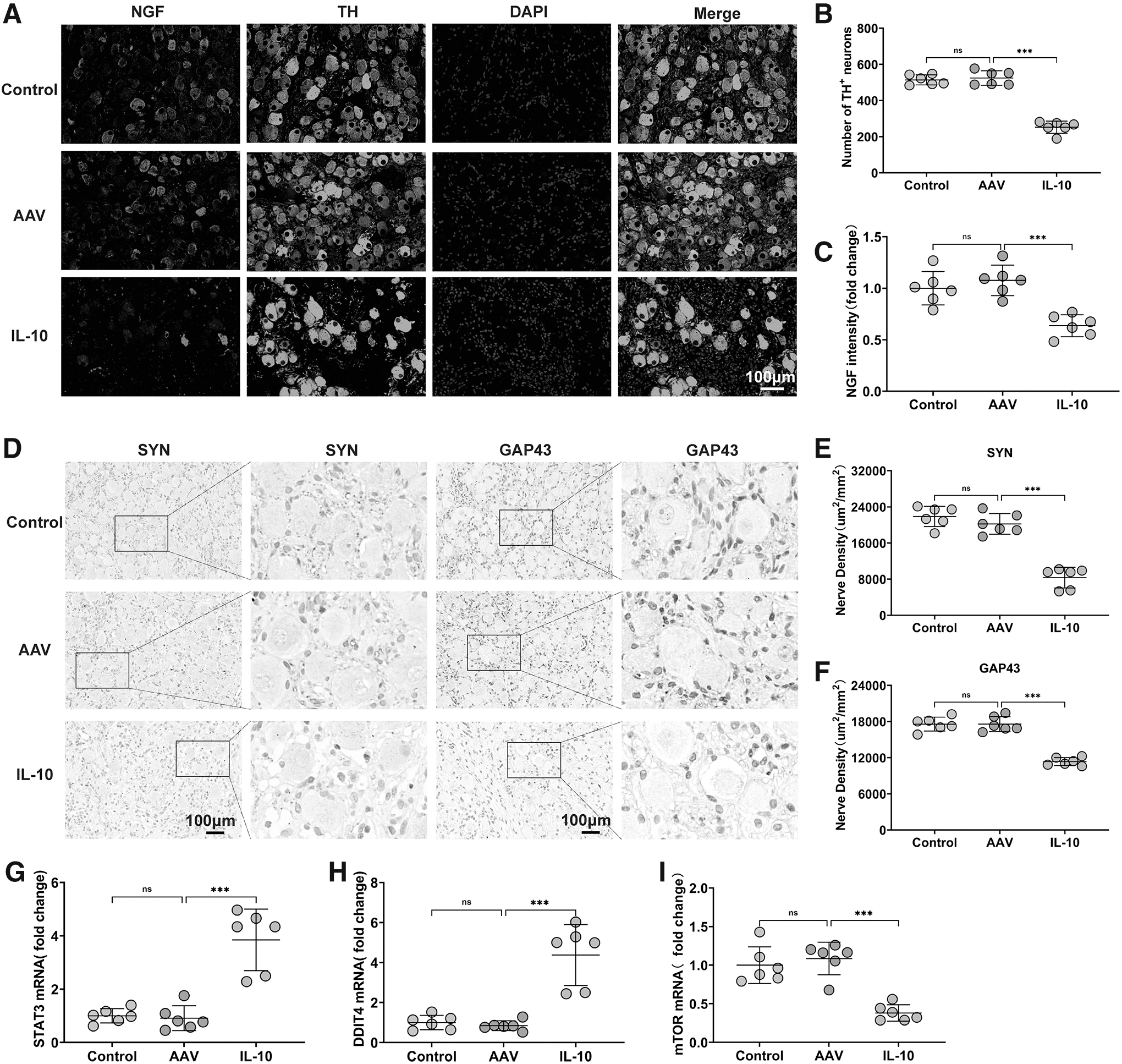
IL-10 overexpression modulated the CANS and decreased LSG function
The activity of the CANS can be noninvasively evaluated using AECG. To investigate the effects of IL-10 overexpression on the CANS, we collected AECG data from all dogs on both days 0 and 30 (Supplementary Table S1). No obvious differences were observed in the baseline characteristics of all these groups.
Thirty days after injection, no significant difference was observed in standard deviation of the N–N intervals (SDNN), standard deviation of the averages of N–N intervals (SDANN), deceleration capacity of rate (DC), low frequency (LF), high frequency (HF), and the ratio of LF to HF (LF/HF) between the control and AAV groups.
However, compared with the AAV group, we observed increases in SDNN, SDANN, and DC (Fig. 3A–C). IL-10 overexpression also resulted in lower LF and LF/HF values and higher HF values (Fig. 3D–F), reflecting decreased sympathetic activity and increased vagal activity. These results indicate that IL-10 overexpression modulates CANS activity and plays a protective role against arrhythmia.
Effects of IL-10 overexpression on the CANS. SDNN, SDANN, DC, LF, HF, and LF/HF at days 0 and 30 are shown in
The elevated blood pressure changes in response to LSG stimulation were considered to represent LSG function. Left femoral artery systolic blood pressure (SBP) changes were collected (Supplementary Table S2), and we used maximal SBP change to evaluate LSG function. As shown in Fig. 4A–E, the maximal SBP increased equally after LSG electrical stimulation at five different voltage levels in the control and AAV groups; however, in the group in which IL-10 level differed, the maximal SBP changes were lower.
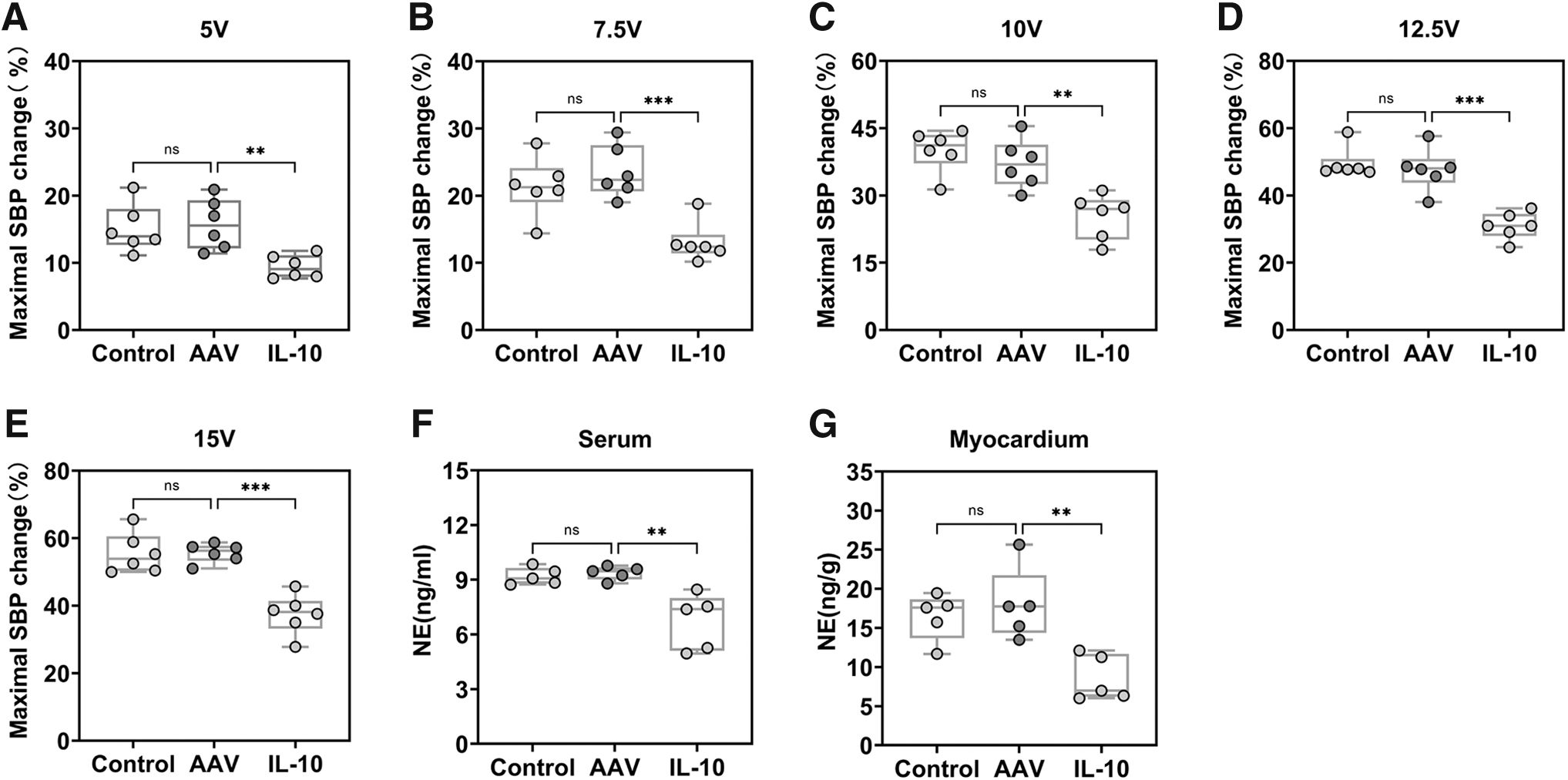
Effect of IL-10 overexpression on LSG function and NE.
Further, the neurotransmitter norepinephrine (NE) (Fig. 4F, G) in both the serum and myocardium was measured to confirm the inhibiting effects of IL-10 overproduction on the CANS. Chronic IL-10 intervention decreased NE levels in both the serum and myocardium.
IL-10 overexpression improved the ventricular effective refractory period and decreased the VA scores after AMI with attenuating post-MI myocardium inflammation
Ventricular effective refractory period (ERP) is commonly used to evaluate the electrophysiological properties of ventricles. ERP at three different sites of the left ventricle, including the left ventricular base, the left ventricular median area, and left ventricular apical area, was evaluated in all experimental dogs (Supplementary Table S3). IL-10 overproduction in the LSG prolonged the ERP in the IL-10 group (Fig. 5A–C).
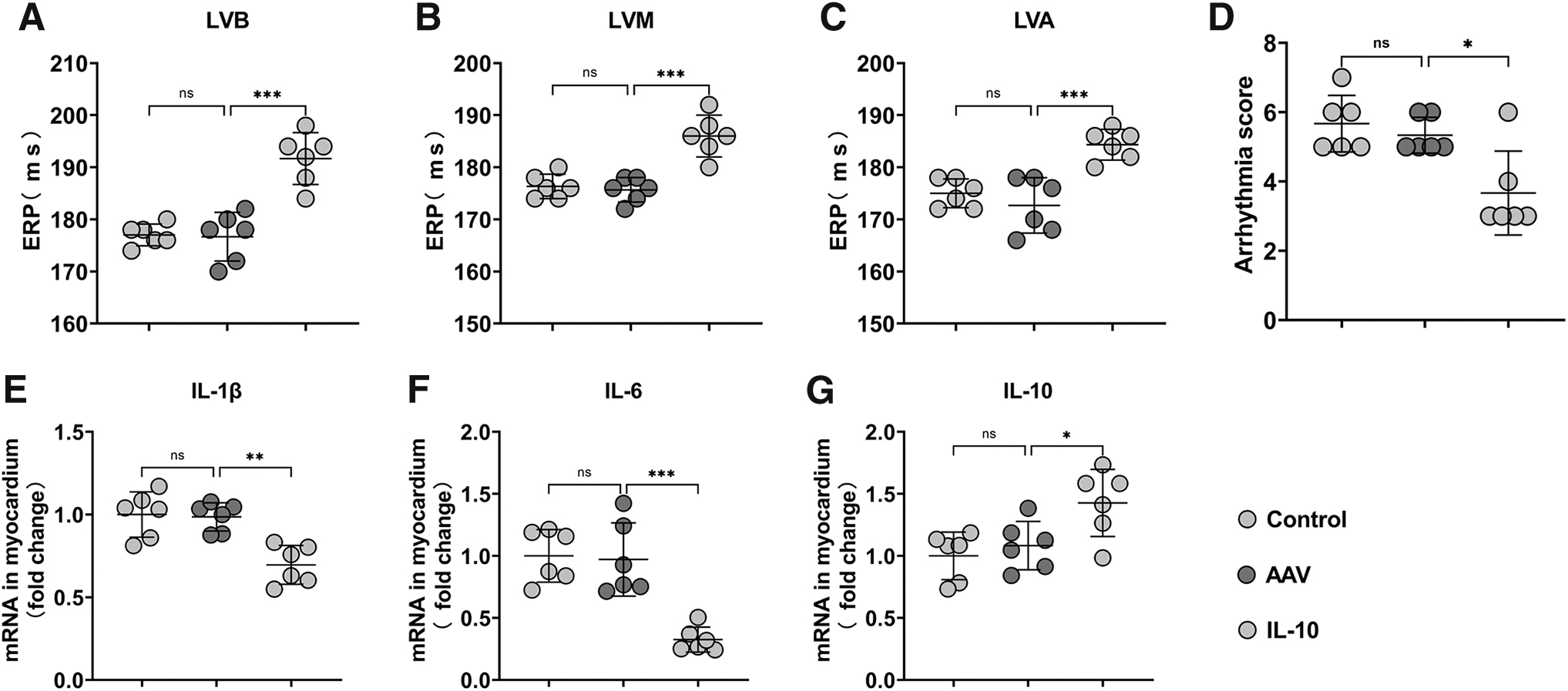
Effects of IL-10 overexpression on electrophysiological stability and inflammation in the myocardium after MI.
To verify whether IL-10 overexpression could effectively suppress acute ischemia-induced VA by modulating the LSG, VA events after acute ischemia were recorded in all groups. These events included ventricular premature beats (VPBs), ventricular tachycardia (VT), and ventricular fibrillation (VF). Depending on the duration, the VT includes sustained VT (SVT, persisting more than 30 s) and non-sustained VT (NSVT, spontaneously terminating within 30 s). IL-10 significantly reduced the number of VPBs and SVT events (Supplementary Fig. S2A, B). IL-10 overproduction resulted in a lower incidence of sustained VT and VF (Supplementary Fig. S2C).
VA scores were calculated throughout ischemia and compared across the three groups. IL-10 pre-treatment during ischemia reduced the arrhythmia score (Fig. 5D), indicating a decrease in arrhythmia severity. Inflammatory factors were measured to assess the effect of IL-10 overproduction in the LSG on myocardial inflammation levels after AMI. Real-time PCR showed that the levels of the pro-inflammatory cytokines IL-1β (Fig. 5E) and IL-6 (Fig. 5F) were downregulated, whereas that of IL-10 (Fig. 5G) was upregulated in the IL-10 group. These changes indicated that chronic IL-10 overexpression effectively attenuated the level of myocardial inflammation after AMI.
DISCUSSION
This study demonstrated that AAV-mediated IL-10 chronic overexpression in the LSG could prevent acute ischemia-induced VA. We observed that IL-10 overexpression in the LSG mediated the modification of the SNS, leading to the suppression of LSG function and the sympathetic components of heart rate variability (HRV). In addition, the stability of ventricular electrophysiology was improved. These findings suggest that chronic IL-10 overproduction has an inhibitory effect on the SNS in vivo, thereby exerting protective effects against MI-induced VA.
Sympathetic overactivation and inflammation play critical roles in the initiation and perpetuation of VA in patients with AMI. Anatomically, the CSG plays a key role in regulating ventricular electrophysiology. 13 Studies have demonstrated that the inhibition of LSG function can enhance cardiac electrophysiological stability and reduce the occurrence of VA after MI. 14 In this study, we demonstrated, for the first time, that IL-10 transgene expression in the LSG reduces LSG function, which is evident from the attenuation of elevated SBP following electrical stimulation of the LSG. To confirm that IL-10 reduces sympathetic outflow, we examined the myocardial and serum NE content, and the results documented significant reductions in both myocardial and serum NE content.
In addition, we assessed the HRV by using AECG, and IL-10 intervention significantly decreased LF and LF/HF, whereas increasing HF. HRV has emerged as the most valuable non-invasive test to assess autonomic nervous system function, and has been identified as a strong predictor of cardiovascular events, 15 although other factors such as genetics may be important determinants of HRV. 16 Compared with nerve recording measurements under anesthesia, AECG data can reflect the autonomic activity of experimental animals in awake and free conditions, making them more representative. Previous studies have shown that cardiac sympathetic activation shortens ventricular ERP. 17 We further evaluated ventricular electrophysiological parameters. IL-10 expression in LSG was found to effectively prolong ventricular ERP and reduce arrhythmia scores after MI. In summary, our results confirm that IL-10 transgene expression can reduce VA after MI by inhibiting sympathetic activation.
Although IL-10, as an anti-inflammatory cytokine, has been described with strong neuroprotective functions in previous studies, 11,18 our results suggest that chronic IL-10 overexpression may lead to cardiac sympathetic neuron degeneration. Importantly, IL-10 has dual effects on neurons. 19,20 Ip et al. found that IL-10 could inhibit the mTOR signaling in a STAT3–DDIT4-dependent manner. 21 Previous studies have shown that DDIT4 plays an essential role in neuronal autophagy and apoptosis through the mTOR signaling axis. 22,23 DDIT4 could also inhibit mTORC1 and reduce synaptogenesis in the nervous system. 24 In addition, the inhibitory effect of IL-10 overproduction on cardiac sympathetic neurons reported herein aligns with other research showing that IL-10 overproduction compromises neural stem cell survival, leading to a reduction in hippocampal neurogenesis in adulthood. 25 Sanchez-Molina et al. demonstrated that IL-10 overproduction could modify microglial receptors, which are involved in neuronal communication, leading to reduced neurogenesis. In this study, chronic IL-10 overproduction came into play in a physiological state, unlike in our work, where enhanced neurogenesis was observed in a pathological situation following IL-10 injections. 26 This difference may be the cause of the varying effects of IL-10. In addition, cholinergic transdifferentiation of cardiac sympathetic neurons has been observed, 27,28 whether the decrease of TH+ neurons is due to cholinergic transformation is also an issue worthy of attention. In any case, the exact mechanism remains to be further studied.
The CANS takes a prominent part in the genesis of many life-threatening VA. 29 Modulation of the cardiac sympathetic tone has been considered a potential therapeutic strategy for treating VA. In the past few decades, surgical cardiac sympathetic denervation, 30 percutaneous LSG blockade, 31 and transcutaneous magnetic stimulation 32 have been conducted in the clinical trial setting. However, many drawbacks have prevented the widespread use of these strategies in clinical practice. 33,34 To date, it has been proven in several animal models with neuroimmune diseases that IL-10 transgene expression could play therapeutic roles. 35,36 Compared with the abovementioned neuromodulation strategies, gene therapy may be a more promising strategy for regulating the CANS. Gene therapy, though, possesses the perfect advantage of sustained influence benefits over traditional drug treatments, ensuring the security of the genome, which is also very important. Moreover, further interpretation is required to clarify how to achieve a more ideal viral transfection for safety risk assessment and the evaluation of clinical translation prospects. Ultrasound-guided techniques 37 and thoracoscopy 38 are maneuverable and less invasive approaches for LSG microinjection. These methods have been widely used in clinical practice and could be effectively implemented for LSG delivery. Our study shows that IL-10 transgene expression in vivo can achieve stable, safe, and relatively long-term IL-10 overproduction effects, thereby inhibiting the cardiac sympathetic tone.
In conclusion, we not only revealed the effects of IL-10 overproduction on LSG using a canine model but also verified that CANS intervention using IL-10 can effectively suppress VA after MI. Gene therapy based on CANS neuromodulation targeting IL-10 overproduction may be a potential treatment for cardiovascular disease associated with SNS overactivity.
Study limitations
This study has several limitations. First, we only investigated the effects of chronic IL-10 overexpression in acute ischemia-induced VA. The lasting effects of IL-10 overexpression on preventing ischemia-induced VAs in chronic MI models need to be verified. Second, while we have verified that IL-10 overexpression in the LSG inhibits cardiac sympathetic nerve activity, and restrains the LSG activation, the exact mechanisms underlying these effects need to be verified in future studies. Third, we tried to use HRV to assess the activity of CANS, but HF/LF ratio, a marker of autonomic nervous system balance, did not provide direct and strong evidence, and a more accurate assessment method is worth applying. Finally, the results of our experiment only proved the effectiveness of the AAV-mediated overexpression of IL-10 in preventing ischemia-induced VA. Further experiments, including dose-response relationship studies, need to be completed to gain a more comprehensive understanding of this therapeutic approach.
CONCLUSIONS
Chronic IL-10 overproduction modulates cardiac sympathetic remodeling and prevents AMI-induced VA. Neuromodulation through AAV-mediated IL-10 overexpression may be a potential neuroimmunotherapy approach for preventing ischemia-induced VA.
Footnotes
AUTHORs' CONTRIBUTIONS
R.L, Y.W., H.J., and B.T. designed the study. R.L., L.Z., C.P., C.W., X.X., R.H., and Y.L. conducted the study. R.L., Z.L., Y.L., and W.T. analyzed the data. R.L., C.P., L.Z., Y.W., and L.Y. drafted the manuscript, and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
AUTHOR DISCLOSURE
No competing financial interests exist.
FUNDING INFORMATION
This work was supported by the grants from the National Natural Science Foundation of China (Nos. 82270532, 82241057, 81970287), and the Foundation for Innovative Research Groups of Natural Science Foundation of Hubei Province, China (2021CFA010).
SUPPLEMENTARY MATERIAL
Supplementary Data S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.