
Abstract
MicroRNAs (miRNAs) are crucial regulators of gene expression involved in various pathophysiological processes. Their ability to modulate multiple pathways simultaneously and their involvement in numerous diseases make miRNAs attractive tools and targets in therapeutic development. Significant efforts have been made to advance miRNA research in the preclinical stage, attracting considerable investment from biopharmaceutical companies. Consequently, an increasing number of miRNA-based therapies have entered clinical trials for both diagnostic and therapeutic applications across a wide range of diseases. While individual miRNAs can regulate a broad array of mRNA targets, this also complicates the management of adverse effects seen in clinical trials. Several candidates have been discontinued due to toxicity concerns, underscoring the need for comprehensive risk assessments of miRNA therapeutics. Despite no miRNA-based strategies have yet received approval from regulatory agencies, prominent progress in the miRNA modulation approaches and in the nano-delivery systems have been made in the last decade, leading to the development of novel safe and well-tolerated miRNA drug candidates. In this review, we present recent advances in the development of miRNA therapeutics currently in preclinical or clinical stages for treating both rare genetic disorders and multifactorial common conditions. We also address the challenges related to the safety and targeted delivery of miRNA therapies, as well as the identification of the most effective therapeutic candidates in preclinical and clinical trials.
INTRODUCTION
MicroRNAs (miRNAs) are a class of non-coding RNAs with an average length of 22 nucleotides (nt). They finely regulate gene expression post-transcriptionally through a powerful mechanism of sequence-specific recognition. More than 2000 different miRNAs have been identified in humans to date (many of which are conserved in other animals) with tissue- and/or developmental stage-specific expression. Each miRNA is predicted to modulate about 200 mRNA targets, resulting in a pleiotropic fine-tuning of related transcripts that confers robustness to pathway regulation. 1
miRNA biogenesis is a complex multistep process (reviewed in 1,2 ) (Figure 1). They are transcribed from both exons and introns of protein-encoding genes or intergenic regions by RNA polymerase II or III to form long primary miRNAs (pri-miRNAs). Each pri-miRNA contains a self-complementary region, which forms a hairpin. pri-miRNAs are further processed in the nucleus by the Drosha-DGCR8 microprocessor complex to generate a miRNA precursor (pre-miRNA) of about 70 nt. In noncanonical biogenesis, intron-encoded pre-miRNAs (mirtrons) can be directly processed along with their co-encoded transcripts through spliceosomes. The pre-miRNA is then exported to the cytoplasm through the XPO5/RanGTP complex and cleaved again by the RNase III endonuclease Dicer and the double-stranded (ds) RNA binding protein TRBP into a miRNA duplex of approximately 22 nt (miRNA/miRNA*): one of two strands is selected as a guide (miRNA), while the complementary passenger one (miRNA*) is usually subject to degradation. The mature single-stranded (ss) miRNA product is then incorporated in a ribonucleoprotein complex known as miRNA containing RNA-Induced Silencing Complex (RISC), containing AGO proteins, and several additional factors.

In most cases, miRNAs direct post-transcriptional repression of mRNA targets. This process is mediated through imperfect binding to target sequences, primarily located in the 3′-untranslated regions (UTR). 2 A critical component in recognizing these target sequences is the “seed region,” a 7–8 nt region in the mature RNA sequence, usually found at positions 2–7 from the miRNA 5’-end. 1,3 Additionally, non-canonical miRNA binding sites (MBSs) have been detected in other regions of a mRNA target, 4 such as in the coding sequence and 5′-UTR, which ensures silencing effects on gene expression 5,6 or even within gene promoters as a way to promote transcription. 7 The binding of the RISC to its target can result in different outcomes. Typically, this binding is partially complementary to the target sites, leading to translation repression. However, in cases where the binding is fully complementary (mainly in plant cells), it results in the degradation of the mRNA target.
miRNAs seem to regulate at least one-third of the human protein-coding genes. 8 Thus, they are involved in a variety of biological processes regulating different cellular and tissue functions, and their dysregulation has been shown in multiple human diseases, making miRNAs valid biomarkers and potentially powerful therapeutic targets.
miRNA therapeutics may be applied to reverse pathological changes in miRNA expression by enhancing or restoring endogenous miRNAs that can act as suppressors of disease, or reducing the expression or blocking the function of miRNAs that act as drivers of pathology. 9 Moreover, modulation of miRNA may also be applied as a therapeutic tool to synergistically modulate different genes and pathways that are deregulated in a disease condition. Indeed, a variety of preclinical and clinical trials were in progress, although some clinical studies were interrupted because of clinical and business issues. In this review, we introduce new advances in miRNA therapeutics development that are currently in the preclinical or clinical stages for the treatment of both rare genetic disorders and multifactorial common conditions. We also address the challenges related to the safety and targeted delivery of miRNA therapies and the identification of the most effective therapeutic candidates.
miRNA THERAPEUTICS DEVELOPMENT
miRNAs are emerging as powerful tools and targets in therapeutic development due to their pivotal role in regulating gene expression and their involvement in a wide array of diseases. Several strategies have arisen to use them as therapeutic targets in recent years. These approaches can be categorized into two main groups based on their mechanisms of action: miRNA enhancement or replacement approaches that functionally increase and restore the expression of specific miRNAs, and miRNA inhibitors that block the action of miRNAs (Figure 2). The former category includes miRNA mimics and recombinant viral vectors encoding miRNA sequences. The latter comprises antimiR oligonucleotides (AMOs), miRNA masks, miRNA sponges, and miRNA decoys. 9,10 The following paragraphs will examine these strategies in detail, focusing on the state of the art in preclinical research.
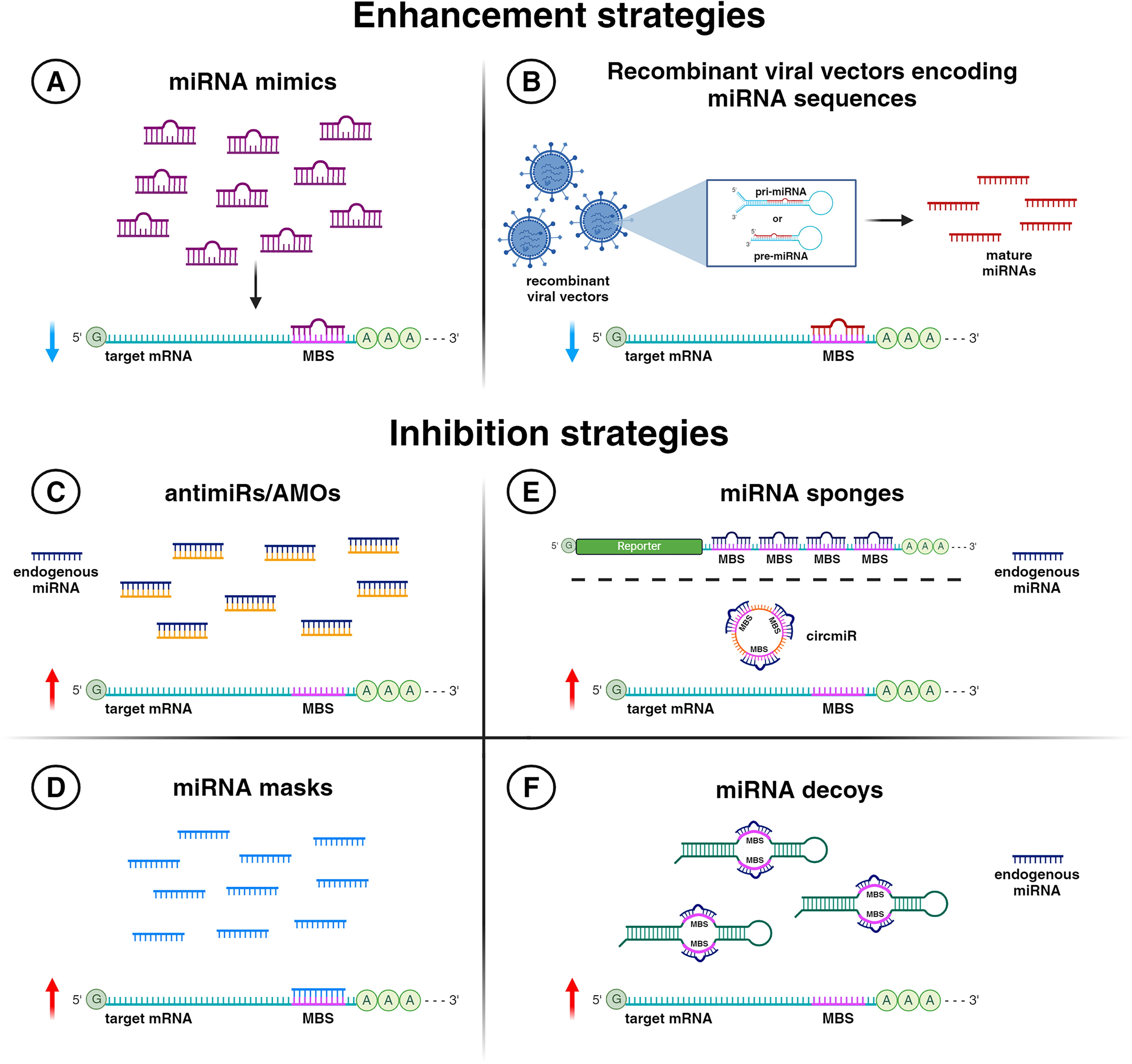
Strategies to increase miRNA expression
miRNA mimics represent the main therapeutic approach to increase miRNA levels. They are synthetic oligonucleotides designed to mimic endogenous miRNAs, aiming to enhance or restore the function of miRNAs that are downregulated or lost in various diseases. These mimics are ds RNAs processed by the cellular machinery into ss miRNAs, which are then incorporated into the RISC to regulate target gene expression by binding to complementary sequences on mRNAs (Figure 2A). They are characterized by the same sequence of the endogenous miRNA, but the passenger strand has some mismatches to avoid functioning as an AMO after its loading into the RISC. 11 As for all the oligonucleotide therapeutics, miRNA mimics need chemical modifications to enhance their stability and efficacy in the body, making them viable candidates for clinical applications. However, the design and chemical improvements of miRNA mimics remain underrepresented in published literature. Indeed, mimics are usually developed by pharmaceutical companies, and as such, the structure of these molecules is covered by intellectual property. Nevertheless, some modifications are shared across all oligonucleotide-based strategies and include the replacement of the 2’-OH moiety with 2’-O-methyl (2’-O-Me) or 2’-fluoro (2’-F) substituents. These improvements have been shown to lower the toxicity and enhance the binding affinity of mimics to their targets. Furthermore, such substitutions, as well as the replacement of the phosphodiester with phosphorothioate (PS) linkages, stabilize these molecules by increasing their resistance to intracellular nucleases. 12 In a recent work, Garreau and colleagues systematically investigated the role of these structural improvements on mimic activity and revealed how critical is not only their presence but also their pattern along the molecule. 13 Other mimic modifications do not focus on the mere RNA structure but instead aim to increase their effectiveness or cellular uptake by combining mimics with other molecules. Mimics conjugation with cholesterol (CHO), indeed, has been successfully applied to increase its cellular uptake upon delivery. 14 Another interesting approach is to directly conjugate mimics and antitumoral molecules to combine the effects of both molecules. In a recent paper, indeed, it has been shown that conjugation of miR-15a and miR-194 mimics with the chemotherapeutic Gemcitabine exerts significant beneficial effects both in in vitro and in vivo models of pancreatic ductal adenocarcinoma. Indeed, although conjugated with a small molecule, both the miRNA mimics retained their capability to inhibit target transcripts. In addition, the conjugated mimics proved to have a stronger inhibitory effect on tumoral growth with respect to the Gemcitabine-only treated controls. 15
Another way to increase miRNA levels is through non-pathogenic recombinant viral vectors carrying miRNA sequences. miRNA sequences can be delivered through this method at different stages of their biogenesis, as pri-miRNAs or as pre-miRNAs, and then further processed by the host cell machinery to mature miRNAs 16 (Figure 2B). Different types of recombinant viral vectors can be used to improve miRNA expression 10 and they will be discussed in the “Viral-based systems” section.
Strategies to decrease miRNA expression
The most common method to inhibit miRNA activity involves synthesizing antisense oligonucleotides with a complementary sequence. AMOs are ss antisense oligonucleotides (ASOs) designed to bind specifically to miRNAs and prevent them from interacting with their target mRNA 11 (Figure 2C). As seen for miRNA mimics, chemical modifications to AMO structure enable substantial advancements in their binding affinity, stability, and target modulation efficiency. 17 Essentially, antimiR modifications are those already developed for ASOs. A first-generation modification is based on replacing phosphodiester with PS linkages to increase nuclease resistance. The second-generation improvements change the 2’-OH group of the sugar ring with 2’-O-Me, 2’-O-methoxyethyl (2’-MOE), or 2’-F moieties, obtaining less toxicity and an enhanced binding affinity to targets. AntimiRs with a 2’-MOE modification, CHO-conjugated to improve intracellular uptake, are called antagomiRs. 18
Finally, the third-generation antimiRs include the locked nucleic acids (LNAs), the peptide nucleic acids (PNAs), and the phosphorodiamidate morpholino oligomers (PMOs), characterized by alterations of the furanose ring to further ameliorate the protection from nucleases, the binding affinity, and pharmacokinetics. 11,19,20 Besides the alterations of the phosphodiester-ribose backbone, ester modification at the 3’-end of AMOs has been proven to increase their activity up to ninefold. 21 In addition, it is possible to conjugate antimiRs to cell-penetrating peptides (CPP) to increase the delivery in the desired tissues. In this regard, miR-23b and miR-218 PMOs conjugated to CPPs have successfully been used to improve the phenotype of a myotonic dystrophy type 1 mouse model with no indications of toxicity. 22 Also, antimiR-21 linked to trimannose is efficiently and selectively delivered to pulmonary macrophages upon inhalation, inhibiting their activation and preventing pulmonary dysfunction in an acute lung damage mouse model. 23 These examples indicate that conjugating antimiRs with specific structures is an encouraging strategy for increasing the effectiveness and delivery of such molecules. Notably, most clinical trials with miRNA therapeutics are based on antimiR strategies, discussed in detail in the “miRNA-based therapeutics in clinical trials” section. 9 These significant milestones in the journey of AMOs confirm the promising therapeutic potential of this approach, instilling confidence in the future of therapeutic development. 23
A different type of AMO methodology is the miRNA-masking approach, referred to as miR-mask. As antimiRs, miR-mask molecules are ss oligoribonucleotides with the 2’-O-Me, or the other chemical modifications discussed above. Unlike AMOs, they do not directly target a miRNA but rather are perfectly complementary to the binding site of that miRNA in the 3’-UTR of the target mRNA (Figure 2D). This strategy is unique in its dual specificity, being both miRNA-specific and gene-specific. It allows for the precise inhibition of the miRNA action on a particular gene without affecting the miRNA regulation of other genes. 24,25
Another way to inhibit miRNA activity is through miRNA sponge technology. First described in 2007 by Ebert and colleagues, artificial miRNA (a-miRNA) sponges are tandemly repeated oligonucleotide sequences with MBSs for a mature miRNA of interest. The number of MBSs needed to achieve optimal inhibition ranges from 4 to 10, separated from each other by some nt (spacers). A higher number of MBSs can affect the sponge’s effectiveness, increasing the risk of construct degradation. MBSs can be totally complementary to miRNAs or characterized by central mismatches (positions 9–12 of MBS), named bulges (Figure 2E, top). This last modification makes sponges more efficacious, probably due to a more stable binding with miRNAs within the RISC complex. 26,27 miRNA sponges offer several advantages, making them a trustworthy therapeutic alternative to antimiRs. They are typically specific for a single miRNA due to their reliance on extensive sequence complementarity beyond just the seed region. Therefore, inhibiting a miRNA family might require using different MBSs to target various family members. However, if the sponge construct is highly expressed, it can effectively bind to and block the activity of an entire miRNA family sharing the seed sequence. miR-181 sponge, for example, has been proven to inhibit the whole miR-181 family (miR-181a, −181 b, −181c, −181d) in a cardiac cell line. 28 Delivering oligonucleotides in vitro or in vivo is often challenging, as cells are frequently resistant to the uptake of exogenous nucleic acids. In this context, a significant advantage of miRNA sponges is the possibility of their delivery through viral vectors. Typically, sponges are expressed downstream of an open reading frame coding for a reporter gene (e.g., the Green Fluorescent Protein, GFP), allowing an easy check for their expression in the desired cell system or tissue of interest. Importantly, unlike AMOs, viral vector-delivered miRNA sponges eliminate the need for frequent administrations to maintain miRNA inhibition due to their continuous expression. This characteristic is particularly beneficial in post-mitotic tissues, such as the neuronal one, ensuring a long-term efficacy of the therapeutic approach. Additionally, sponge activity can be restricted to specific cell types or tissue/organs using specific promoters, thereby avoiding undesired outcomes in other parts of the body. 26,27 For example, a sponge specific for miR-181a and miR-181b was cloned downstream of the GFP gene and delivered it in in vivo models of retinal diseases through adeno-associated viral (AAV) vectors as therapeutic strategy. The use of specific promoters to express the miR-181a/b sponge in different retinal cell types allowed for a better understanding of the therapeutic contribution of the sponge in distinct retinal compartments. 29 Multi-potent miRNA sponges have been optimized to simultaneously target various miRNAs acting in the same process. For instance, a sponge against miR-21, miR-155, and miR-221/222 has been proven to show an antitumor function in breast and pancreatic cancer cells. 30 Alongside the a-miRNA sponges discussed so far, natural miRNA sponges, called circular RNAs (circRNAs), have been identified more recently. 31 Unlike the previously described sponges with linear sequences, the circRNAs, as their name suggests, are characterized by circular sequences, which make them resistant to exonucleases due to the lack of 5′/3′-ends. 32 circRNAs are not only considered more stable than linear sponges but also capable of binding a higher number of miRNA molecules due to the presence of a really huge number of MBSs on each circRNA: 73 conventional MBSs have been identified for miR-7 on the ciRS-7 circRNA. 31 For these reasons, engineered circRNA sponges, known as circmiRs, have been promptly developed to explore their therapeutic potential 33 (Figure 2E, bottom). Compared to earlier versions, they require lower dosages and have longer half-lives. Therefore, although not yet tested in clinical trials, sponge technology quickly progresses and offers promising prospects for rapid clinical application.
miRNA decoys represent a variation of miRNA sponges. 27 Like sponges, decoys consist of tandem sequences complementary to miRNAs, which can effectively inhibit their activity. They are characterized by stem and loop secondary structures, the loop of which contains MBSs for miRNAs (Figure 2F). In 2009, a miRNA decoy system was first described. 34 Several decoy structures were designed and tested to determine the most efficient configuration for repressing miRNA function. It has been showed that a Tough Decoy (TuD) with two MBSs connected by 3-nt-linkers to two flanking stem structures induced specific, strong, and long-term miRNA suppression in mammalian cells, 34 in light of which further decoys used in preclinical studies were developed.
PRECLINICAL PROGRESS OF miRNA-BASED THERAPIES
Cancer
It is extensively recognized that many miRNAs are dysregulated in tumors. Depending on the context, they can act as either oncogenes (oncomiRs) and/or tumor suppressors. 35 –37 Therefore, it is unsurprising that significant attention has been paid to developing miRNA-based anticancer therapies in recent years. miR-101, for instance, was found to be downregulated in endometrial cancer and colorectal carcinoma. Overexpression of miR-101 through miR-101 mimics inhibited cell proliferation, invasion, and tumor growth. 38,39 Also, miR-184 is a thoroughly investigated miRNA for its role as a tumor suppressor. It is predominantly downregulated in multiple malignancies, such as lung cancer, endometrial carcinoma, colorectal cancer (CRC), breast cancer, and many others. 40 It was demonstrated that miR-184 mimic significantly inhibited CRC cell proliferation, migration, and invasion. 41
Some miRNAs play pivotal anti-tumoral activities during human hematopoiesis. It is the case of miR-193b, which downregulates several MAP-kinase signaling cascade genes. Such activity is critical for the correct regulation of cell proliferation. miR-193b was found to be downregulated in many cases of acute myeloid leukemia, irrespective of the genetic etiology of the disease. Treating patient-derived blasts with a miR-193b mimic showed strong anti-proliferative and pro-apoptotic activities. Furthermore, the strategy significantly decreased cancer spread and progression in in vivo models. 42 Similarly, lentiviral delivery of miR-15a and miR-16 in a mouse model of chronic lymphocytic leukemia showed both short- and long-term effects, reducing malignant B-1 cells and decreasing splenic and hepatic involvement with minimal systemic toxicity. 43
Several studies have highlighted the crucial role of miR-181 family members in cancer, functioning either as tumor suppressors or as oncomiRs. These microRNAs influence a variety of target mRNAs involved in key cancer-related pathways. Recent research indicates that miR-181 family members are implicated in various solid tumors as well as hematological malignancies. 44 Particularly, miR-181a/b mimics can block melanoma growth in melanoma cells resistant to BRAF inhibitors, while miR-181c/d mimics affect cell viability and enhance sensitivity to chemotherapy in biliary tract cancer cell lines. 45,46
Conversely, miR-21 is extensively investigated as an oncomiR causing the development of various malignancies. Notably, the inhibition of miR-21-5p through an RNA decoy system led to a significant tumor growth arrest in in vitro and in vivo models. 47 Likewise, the oncomiR miR-22 is implicated in the metastatic process of triple-negative breast cancer (TNBC). Its suppression using an LNA-modified antimiR-22 has effectively reduced epithelial-mesenchymal transition, suppressed metastatic spread, and increased survival of TNBC mouse xenograft models. 48
Other multifactorial disorders
Aside from cancer, modulation of different miRNAs has also been applied in treating multifactorial diseases, such as cardiac dysfunctions, obesity, and neurodegenerative disorders (e.g., Parkinson’s and Alzheimer’s disease). In this regard, an LNA-modified antimiR-132 has been proven effective and safe in multiple models of heart failure. It successfully restores the expression levels of miR-132 targets, Foxo3 and Serca2a. FOXO3 is involved in antihypertrophic effects and in the regulation of autophagy, while SERCA2a contributes to myocardial relaxation by enhancing calcium reuptake. 49,50 Additionally, circmiR sponges targeting miR-132 and miR-212 (recognized as cardiac pro-hypertrophic miRNAs) mitigated the phenotype and preserved the cardiac activity of a transverse aortic constriction mouse model through the attenuation of stress markers such as Nppb, which regulates cardiovascular homeostasis, and Myh7, which plays a role in the cardiac muscle contraction. 33 Furthermore, a second-generation AMO with the 2’-MOE modification against the liver-specific miR-122 had beneficial effects in a diet-induced obesity mouse model, decreasing plasma cholesterol levels and significantly ameliorating liver steatosis. 51 The inhibition of miR-122 results in a decrease of several transcripts acting on lipid metabolism, i.e., Acc2 and Scd1, which are key regulators of fatty-acid oxidation and synthesis. Also, the increase of the AMPK enzyme has been detected. This metabolic sensor, among its various functions, promotes fatty-acid oxidation and inhibits fatty-acid synthesis. However, it is still not clear the connection between AMPK activation and downregulation of the aforementioned genes involved in lipid metabolism. 51 The upregulation of miR-221/222 has been reported in patients with metabolic-associated steatohepatitis (MASH, formerly non-alcoholic steatohepatitis or NASH). Treatment of a MASH murine model with miR-221/222 AMOs considerably arrested steatohepatitis progression and liver fibrosis increasing hepatic TIMP3 (an important regulator of inflammation and fibrosis) levels, and reducing collagen related genes (a-SMA, Col1a1, Col1a2, Col3a1). 52 Remarkably, antimiR-140-5p was demonstrated to have beneficial effects in a rat model of Alzheimer’s disease (AD), improving learning and memory deficits caused by the accumulation of amyloid-β (Aβ) oligomer. A possible mechanism of action is through the augmentation of miR-140-5p main targets, Pin1 and Adam10, observed in treated animals. 53 PIN1 exerts a pivotal role in AD progression, stimulating the non-amyloidogenic processing of amyloid-beta precursor protein (APP) and, therefore, reducing Aβ production. 54 ADAM10 is responsible for the constitutive and regulated alpha-secretase cleavage of APP. 55 In addition, Stein et al. developed a miR-181 TuD molecule showing a significant neuroprotective effect of the construct in a mouse model of Parkinson’s disease (PD). RNA-seq analysis in this mouse model revealed that miR-181s act on genes that are involved in synaptic signaling, axon growth and stability, ion transport, as well as mitochondrial function as already reported. 57 Some of these are reported to be protective in PD models (e.g., Kcnh1, Slc2a3, Atg5, Nrg1) or to act into the establishment and maintenance of the dopaminergic cells (e.g., Rspo2, Bmp2). 56
Genetic disorders
Despite the efficacy of miRNA-based therapeutic strategies being widely reported in multifactorial diseases, considerable scientific evidence demonstrates the validity of this therapeutic approach also in genetic disorders. Various preclinical studies have successfully been conducted on miRNA therapeutics in genetic disorders. For instance, a study on spinal and bulbar muscular atrophy (SBMA), an inherited neurodegenerative disorder, showed how the AAV-mediated delivery of miR-196a in an SBMA mouse model ameliorated the motor impairment of this murine model. 58
An important work was conducted by Gomez et al on the antimiR-21 effects in a mouse model of Alport nephropathy. The authors demonstrated that antimir-21 treatment drastically ameliorated disease phenotypes and improved the survival of Alport mice with no adverse effects. 59 Notably, these promising findings reached the clinical trial stage. Recently, a study on a new miRNA-based therapeutic strategy for Angelman syndrome (AS) was published. AS is a severe neurodevelopmental disease caused by mutations in the maternal allele of the paternally imprinted UBE3A gene. The most studied therapeutic intervention aims to unsilence the paternally imprinted copy of UBE3A with a narrow therapeutic window. For this reason, efforts have been made to develop an alternative approach based on the inhibition of miR-134, which directly regulates the UBE3A transcript. Interestingly, the use of antimiR-134 resulted in improvements in behavioural tests and seizure suppression in both juvenile and older mice, showing a broader therapeutic window with this alternative strategy. 60
Several miRNAs have been reported to be important for retinal function, and their dysregulation has been associated with different retinal disorders. 61 miR-204, for example, is very abundant in the retina, and it has been proven that AAV-mediated delivery of this miRNA in different models of inherited retinal diseases protects retinal function in a mutation-independent manner. 62 Similarly, sponge-mediated inhibition of miR-181a/b ameliorates retinal morphology and visual function of various mouse models of retinitis pigmentosa. 29 Interestingly, miR-181a/b were also shown to be key modulators of mitochondrial pathways, and their genetic inactivation resulted protective in different mitochondrial disorders independently from the pathogenetic mutations underlying the disorders. 57 Also, we demonstrated that PMO-mediated downregulation of miR-181a/b is protective in a medaka fish model of microphthalmia with linear skin lesions, a rare inherited form of mitochondrial disease, notably reducing cell death in the eye and brain. 57 These findings further underline the therapeutic potential of miRNA modulation, which can be particularly effective in heterogenous genetic disorders since they can act in a gene/mutations independent fashion. 61 , 63
All miRNA therapeutic technologies discussed in this section and applied to various types of diseases show promising preclinical results, confirming the rapid development of the miRNA therapy field and hopefully paving the way for clinical trials.
ADVANCES IN miRNA-BASED THERAPEUTICS DELIVERY SYSTEM
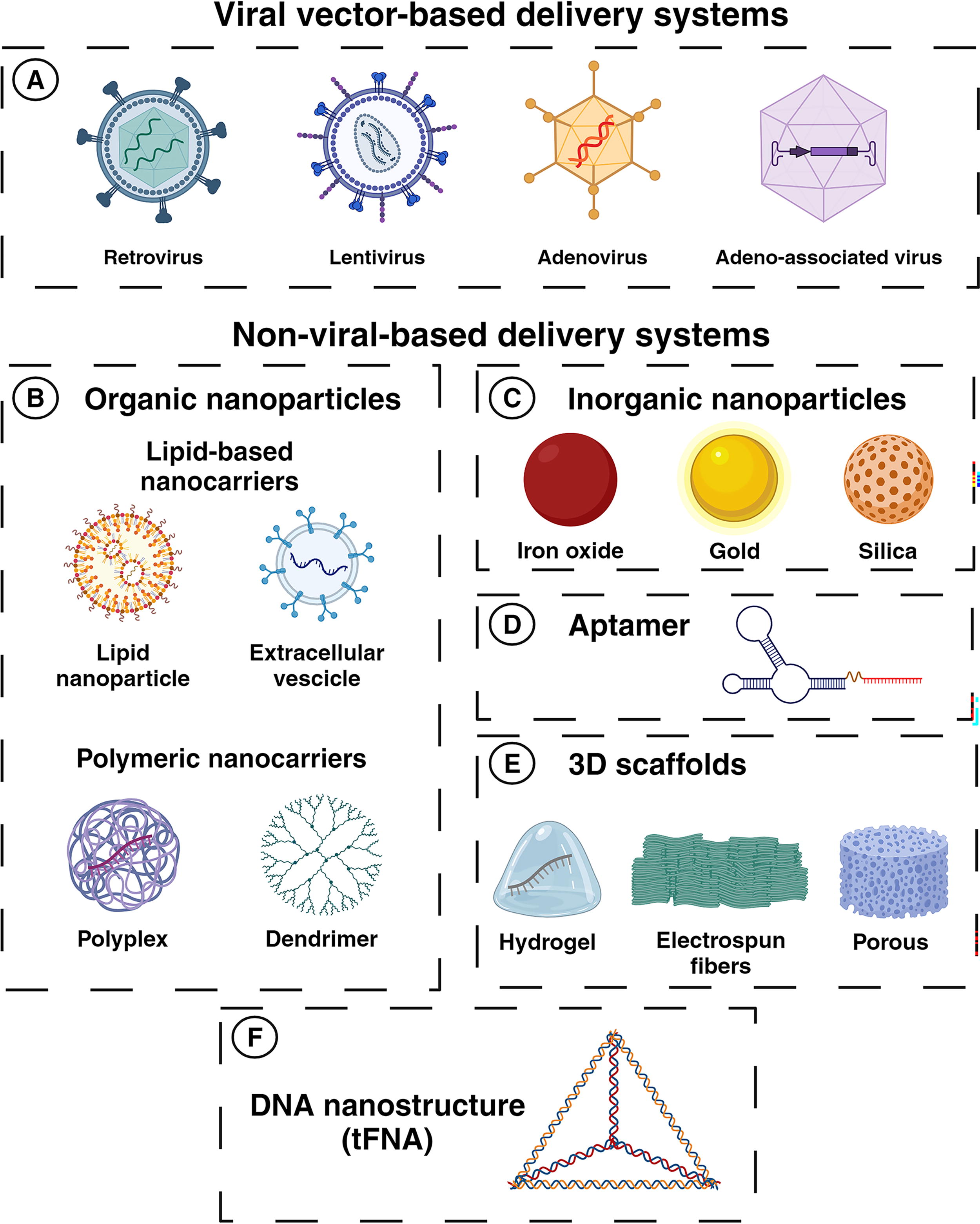
The design of secure and efficient systems to deliver miRNA therapeutics to the target tissues represents a major challenge for the clinical application of miRNA-based therapies. A powerful delivery method should overcome miRNAs’ early degradation into the blood and poor tissue penetration, over the risk of immunotoxicity and undesired off-target effects. As described before, chemical modifications of the sugar ring or 3’-end have significantly improved miRNA stability and protected against nucleases. Moreover, the buildout of increasingly sophisticated carriers and transport strategies is contributing to addressing miRNA delivery issues. Currently, miRNA therapeutics transport can be achieved through viral vectors, such as retroviral (RV), lentiviral (LV), adenoviral (Ad), and AAV vectors, and non-viral-based systems, which include various types of organic and inorganic vehicles (Figure 3).
Viral-based systems
Preclinical studies have extensively tested non-pathogenic recombinant (r) viral vectors to deliver nucleic acid into target organs or specific cell types. Viral-based systems ensure extremely high efficiency and sustained expression over time, emerging as promising strategies to overexpress or repress endogenous miRNAs. To date, several viral vectors have been developed, each with distinct characteristics that make one vector more suitable than another depending on the therapeutic aim (Figure 3A). 64
RV vectors demonstrated effectiveness in increasing 65 and inhibiting 66 miRNA expression limited to dividing cells. In contrast, LV and Ad systems showed high transduction efficiency and stable expression of miRNAs delivered to dividing, post-mitotic, and terminally differentiated cells, like neurons. RVs and LVs stably integrate into the host cell genome, thus allowing potential persistent transgene expression. However, the integration may occur at undesired locations representing a critical point concerning the safety of the above vectors in clinical applications. 64
Due to their high cloning capacity, LV and Ad vectors are preferred to co-express multiple miRNAs from a single polycistronic transcript. Indeed, polycistronic constructs carrying a-miRNA precursors can be developed to reinforce the silencing of a gene or different genes simultaneously. 67 Moreover, mature miRNA expression, achieved by pre-miRNA precursor format, reached on average several-fold higher levels than mature miRNA format, beyond remaining stable upon long-term virus propagation. For example, LV-pre-miR-195 has been used to treat Aβ plaques in a mouse model of AD. 68 The expression of the miRNA can be further improved by integrating the pri-miRNA flanking sequence at both ends of pre-miRNA fragments into the vector. In this way, the physiological mechanism of miRNA processing is reproduced in the cell, improving its expression efficiency. 64
Like LVs and Ads, AAV vectors can transduce both dividing and non-dividing cells. AAV systems show many advantages including tissue-specific tropism, long-term expression, and high transduction efficacy. Despite the AAVs are the smallest known viral vectors, the tiny size of miRNA genes gets around the restricted capacity of the AAV genome. However, AAV systems represent more appealing alternatives to the other viral vectors especially for their poor ability to transduce antigen-presenting cells notably lowering the risk of a significant immune response from the host. Recent success in clinics makes AAV systems ideal and powerful in in vivo gene delivery platforms compared with the other viral vectors. A remarkable example of rAAV-delivered miRNA therapies was achieved using rAAV9, able to cross the blood-brain barrier to deliver miR-196a to motor neurons. 58 More recently, rAAV9 carrying an a-miRNA targeting SOD1 has been tested as a potential treatment to silence SOD1 expression in amyotrophic lateral sclerosis (ALS) cases. 69 , 70 Moreover, AAV systems represent one of the most promising delivery tools for transducing retina layers. rAAV2/8 vectors expressing a miR‐181a/b sponge construct were used to efficiently transduce retinal pigment epithelium and photoreceptors upon subretinal administration in mouse models of inherited retinal diseases. 29 Currently, a rAAV5-mediated delivery of a-miRNA expression is ongoing in a clinical trial. 71 Overall, the use of viral vectors to deliver miRNAs presents many advantages, mainly due to their higher transfection efficiency and stable expression. However, immunogenicity, liver toxicity at high dosages, and genotoxicity due to viral integration capacity represent issues that need to be solved.
Non-viral-based systems
The use of non-viral carriers, as opposed to viral systems, enormously limits the risk of immunogenicity and cytotoxicity but presents the significant disadvantages of much lower transduction efficiency and a short half-life. However, much progress has been made in the last years to improve nonviral methods including a broad variety of organic and inorganic carriers (Figure 3B–C).
Among organic drug delivery systems, lipid nanoparticles (LNPs) are intriguing due to their biocompatibility and biodegradability (Figure 3B). Liposomes, the earliest generation of LNPs, consist of one or more lipid bilayers of various types of lipids and stabilizers and can vary in size and surface charge (neutral, anionic, or cationic). Positively charged LNPs are the most used to carry nucleic acid due to favorable electrostatic interactions with negatively charged cell membranes. 72 However, lipoplexes (cationic lipid-nucleic acid complexes) require complex preparation methods and drug loading, which result disadvantageous in performing on large scales. 72 Solid lipid nanoparticles (SLNs) and nanostructure lipid carriers (NLCs) are improved lipid-based systems produced using various organic solvent-free methods that are upgradable to production scale. They are composed of a core of solid (SLNs) or a mixture of solid and liquid lipids (NLCs) and stabilized by emulsifier. 72
Extracellular vesicles (EVs) are held responsible for distant cell-to-cell communication, representing other promising candidates for miRNA delivery due to their low immunogenicity and cytotoxicity. Three main types of EVs have been identified based on their intracellular origins: exosomes (EXOs), microvesicles (MVs), and apoptotic bodies (ApoBDs). 73 Secreted by almost all human cells and present in biofluids, exosomes are endosome-derived membrane-bound small EVs. As natural carriers of RNA molecules, EXOs can mediate targeted delivery through receptor-mediated binding, protecting the encapsulated content from RNases in body fluids and escaping from phagocytosis and endosomes. 74 EXOs shuttling miRNAs play a pivotal role in both diagnostics and therapeutics. Engineered EXOs harboring both miR-21 inhibitor and an anticancer drug were exploited to target HER2-expressing cancer cells. 75 MVs are a heterogeneous population of EVs that are shed from multiple cell types during certain pathological and physiological conditions. 73 As examples, MVs packaged with miR-200b, a miRNA with anti-fibrotic properties, and obtained from stimulation of bone marrow mesenchymal stem cells were employed to treat intestinal fibrosis. 76 ApoBDs are membrane blebs specifically generated and released from apoptotic cells that can be used to deliver chemical inputs, as for the endothelial cell-derived ApoBD containing miRNA-126 that triggers the secretion of chemokine protecting mice from atherosclerosis. 77
Apart from lipid-based particles, polymeric nanocarriers have been extensively studied as a novel delivery tool and as a component to enhance above-mentioned systems (Figure 3B, bottom). Polyplexes (negatively charged nucleic acids complexed with positively charged polymers) offer the advantages of ease of synthesis, biodegradability, and low immunogenicity. 78 Among polycations, polyethylenimine (PEI) and chitosan are the most used for miRNA delivery. PEI promotes cellular uptake and endosomal escape and can be attached to polyethylene glycol (PEG) to further improve its performance. For example, PEG-PEI/miR-221/222 was used to transfect prostate cancer cells. 79 Chitosan is a natural biocompatible and mucoadhesive polysaccharide that is able to prevent miRNA degradation with negligible cytotoxicity. Poly(lactide-co-glycolide) (PLGA) is a biodegradable and biocompatible copolyester made of lactic and glycolic acid monomers taken up by cells via endocytosis. As an FDA-approved biomaterial, PLGA shows excellent drug-release properties and, hence, has been frequently used in the clinic. PLGA nanoparticles are often prepared with an additional polycation such as PEI 80 or chitosan to increase miRNA-loading. Dendrimers are hyperbranched globular polymers with multiple terminal functional groups that provide ease of conjugation. Polyamidoamine (PAMAM) dendrimers are positively charged polymers showing high transfection efficiency and very low toxicity compared to other polymers. PAMAM dendrimers can be coated with targeting ligands to favor miRNA delivery to specific cells. 72 , 78 A mixture of cationic lipids and polymeric components represents an additional system that has been exploited to deliver miRNAs. Lipopolyplexes (LPPs) are made of a nucleic acid, like a miRNA, complexed with a polymer that can be either coated or encapsulated in a liposomal structure. Hybrid LPPs show the advantages of both systems, that are high stability, fine cellular uptake, and low cytotoxicity as lipoplexes, high transfection efficiency, better nucleic acid condensation, homogenous particle size, and facilitated endosomal escape as polyplexes. 72
Inorganic vehicles (Figure 3C) have gained interest as miRNA delivery systems suitable for clinical application. Inorganic nanoparticles are realized to be non-toxic, hydrophilic, biocompatible, and highly stable by secure and simplified manufacturing procedures. Gold, silica, and iron oxide nanoparticles are currently employed for miRNA delivery. Gold nanoparticles (AuNPs) are versatile materials easy to manufacture, and less reactive, presenting a high surface-to-volume ratio that can be modified to introduce functional modifications for targeted delivery. 81 , 82 Silica represents a valid alternative to Au since silica nanoparticles (SiNPs) are biodegradable and can be easily functionalized. 81 , 82 Concerning the other inorganic vehicles, iron oxide nanoparticles (IONPs) show unique magnetic properties, such as superparamagnetism, that make them appealing for clinical application. 81 , 82 Superparamagnetic IONPs (SIONPs) exploit their magnetic susceptibility to be driven to target tissues by external magnetic fields. Moreover, SIONPs are efficiently cleared from the human body via the iron metabolism pathways. 81 , 82
Another strategy to promote the selective accumulation of miRNAs to target tissues involves aptamer-based approaches (Figure 3D). Aptamers (Apts) are short synthetic ss oligomers picked from a pool of random sequences through multiple rounds of in vitro selection. 83 Apts show ease of synthesis and modification and can be selected to target both known and even unknown cell surface markers with high affinity binding. Aptamer-miRNA (Apt-miR) conjugates are simply produced by sticky-end annealing and are internalized via receptor binding. Apt69.T is a 2’-fluoro-pyrimidine modified aptamer selected for targeting BCMA-expressing myeloma cells. Apt69.T complexed with miR-137 and antimiR-122 was rapidly and efficiently internalized through the binding of BCMA. 84
The use of 3D scaffolds represents a recent innovation in miRNA delivery. They consist of hydrogels, electrospun fibers, and other porous 3D scaffolds (Figure 3E). Hydrogel composition resembles the one of the native extracellular matrixes and shows many advantages compared to other systems. Indeed, hydrogels may improve RNA stability, reducing the dispersion of therapeutics associated with systemic delivery, limit undesirable off-target toxicities, and avoid the necessity of multiple doses. 85 Hydrogel and electrospun fibers scaffolds can be used to develop integrated systems, as for the miR-21-mesoporous SiNPs encapsulated in an injectable hydrogel matrix, 86 or for the miR-145 carried by a PEG/chitosan polymer complexed in an electrospun fibers membrane. 87
The latest research advances in nanotechnology for miRNA delivery concern the design of DNA nanostructures, which refer to nanomaterial made of synthetic DNA that can serve as 3D scaffolds for the fabrication of more complex structures. Tetrahedral framework DNA (TfDNA) has been used for the design and synthesis of a bio-switchable miRNA delivery system, consisting of a nucleic acid core surrounded by three miRNA molecules with an RNase H‐responsive sequence tail that enables the release of miRNAs upon entering cells. These nanostructures are biocompatible, highly stable, able to easily penetrate cells and tissues, easy to synthesis, and cost-effective representing an ideal vehicle for miRNA therapies. 88 , 89
Ways to improve delivery efficiency
To improve delivery systems’ effectiveness, several component modifications as well as the covalent conjugation with small ligands, peptides, or monoclonal antibodies, may be applied to carriers. For example, certain carriers’ formulations can significantly improve delivery efficiency. Conventional cationic lipids, such as DOTMA {N-[1-(2,3-dioleyloxy)propyl]-N,N,N-trimethylammonium chloride} and DOTAP (1,2-Dioleoyl-3-trimethylammonium propane), are limited by their fixed charge surface that make them unresponsive to pH changes during subcellular trafficking in endosomes. The permanently positive charge of cationic carriers causes also easly aggregation with negatively charged serum proteins and, hence, their rapid clearance in the bloodstream, beyond the toxicity due to their hemolytic activity. 90 Ionizable lipid-based LNP (iLNP), such as DODMA (1,2-dioleyloxy-3-dimethylaminopropane), DODAP (1,2-dioleoyl-3-dimethylaminopropane), and DLin-MC3-DMA (MC3) show the ability to change their charge across different pH levels ensuring the efficient encapsulation of miRNA at low pH, the escape from endosomes and very low toxicity during recycling under physiological conditions holding great potential for the translation of miRNA-based therapies. 91 Moreover, the addition of helper components such as the neutral lipid DOPE (dioleoyl phosphatidylethanolamine), the lipophilic CHO, and the amphiphilic PEG groups at the carrier’s surface enhances the stability and delivery efficiency. Lipoplexes composed of the helper lipid DOPE and the cationic lipid DE {[2-(2,3-didodecyloxypropyl)-hydroxyethyl] ammonium bromide} showed superior miRNA loading efficiency and faster miRNA release as demonstrated to DE/DOPE/miR-1 lipoplexes delivered to adult human cardiac fibroblasts for cardiac cell reprogramming. 92 The CHO/PGEA nanocarrier favorites miR-182 inhibitor delivery into cardiomyocytes. 93 The CHO coating has also been employed in a therapeutic test in clinics for the treatment of the Keloid disorder. 94 Cationic lipids coated with PEG limit the risk of phagocytosis and the interaction with plasma proteins, resulting in prolonged circulation time and improved overall efficiency. Moreover, conjugation with certain proteins may enhance targeted drug delivery. Folate receptors are overexpressed in many cancers and on macrophages, which are abundant in inflammatory diseases; hence, the use of folic acid as ligand on nanocarriers allows for specificity for tumor cells over noncancerous cells or to deliver anti-inflammatory drugs, respectively. 72 N-acetylgalactosamine (GalNAc) is a sugar molecule that recognizes and binds to the asialoglycoprotein receptor, which is abundantly expressed in hepatocytes. GalNAc has been widely used to target miRNAs in the liver, such as GalNAc-conjugated miR-122 inhibitor (RG-101), 95 and a miR-103/107 inhibitor (RG-125/AZD4076) 96 in clinical trials on chronic hepatitis C virus (HCV) and on MASH, respectively. Asp-Gly-Arg (RGD) tripeptide is the most common peptide motif responsible for cell adhesion to the extracellular matrix via integrin receptor binding. The conjugation of the RGD tripeptide has been demonstrated to significantly enhance the uptake of SLNs co-loaded with miR-150 and the small molecule quercetin, known to have anti-angiogenic properties via intravitreal injection in a mouse model of age-related macular degeneration. 97 Recently, human cord blood mesenchymal stem cells-derived exosomes, loaded with miR-26a mimics, have been engineered to conjugate anti-GPC3 single-chain variable fragments on their surface and deliver miR26a to GPC3-overexpressing hepatocellular carcinoma cells. 98 Transferrin (Tf)-conjugated PEI/DOPE lipopolyplexs for synthetic miR-29b (Tf-NP-miR-29b) delivery in cells overexpressing Tf receptor has been designed as anti-cancer strategies in acute myeloid leukemia. 99 SiNPs coated with anti-GD2 have been used to targeted delivery of miR-34a for the treatment of GD2-expressing neuroblastoma cells. 100 TTX-MC138 antimiR10b, in clinical trial for the treatment of advanced solid tumor, consists of an IONP coated with dextran, which further facilitates the rapid uptake of the drug by exploiting the high metabolic activity of cancer cells. 101
miRNA-BASED THERAPEUTICS IN CLINICAL TRIALS
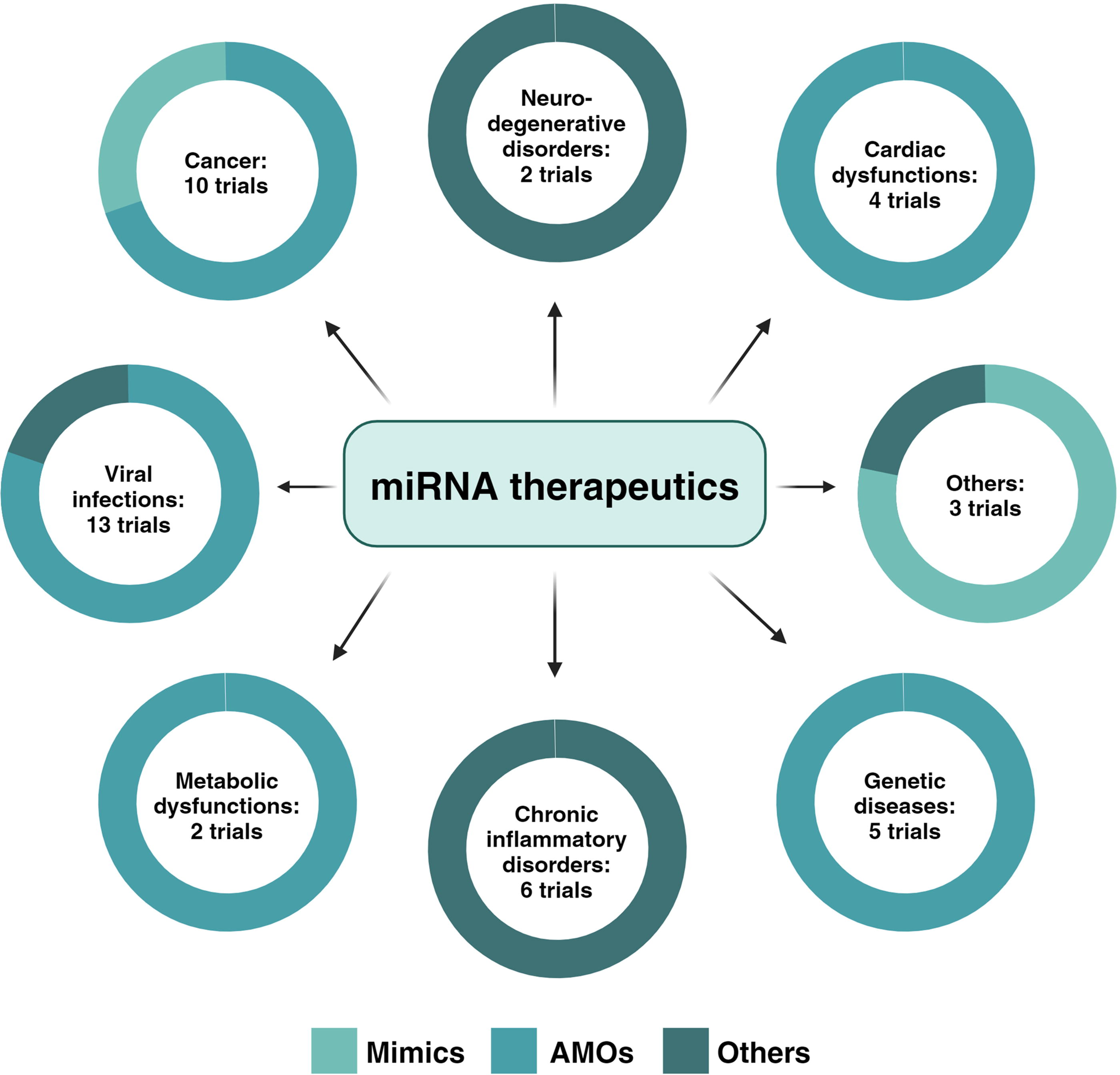
As stated above, miRNAs are emerging as potential diagnostic markers and therapeutic targets in various clinical fields, as confirmed, according to PubMed records, since 2014 by more than 600 articles published under the heading of ‘miRNA-based biomarkers’ and 850 articles searching for ‘miRNA-based therapeutics’. Over the past decade, scientists have documented changes in miRNA expression in many types of cancer, as well as viral infections, cardiovascular diseases, autoimmune diseases, and neurodegenerative disorders, among others (Figure 4). Considerable efforts have been made to advance miRNA preclinical research, raising interest in investing in many biopharmaceutical companies such as SantarisPharma, Mirna Therapeutics, and Regulus Therapeutics. As a result, lately, a growing number of miRNA molecules entered clinical trials for diagnostic and therapeutic applications across a spectrum of diseases. Currently, about 552 studies have been found under the heading “miRNAs” as intervention/treatment searching in the ClinicalTrials.gov portal, which includes a few clinical trials of diagnostic miRNAs recently completed. However, the results have not been reported, and some others related to miRNA therapeutics were interrupted because of clinical and business issues. Nonetheless, the number of clinical trials of upgraded miRNA drug strategies, in progress and just recruiting, is steadily increasing, inclusive of encouraging examples of miRNA-based therapeutics close to approval from regulatory agencies (Table 1).
List of microRNA-Based Therapeutics Entered Clinical Trials to June 2024
Ref, references; HTT, huntingtin; PS, phosphorothioate; MLM, malignant pleural mesothelioma; NSCLC, non-small cell lung cancer; ADPKD, autosomal dominant polycystic kidney disease; SCLC, small cell lung cancer; MM, multiple myeloma; RCC, renal cell carcinoma; MAFLD, metabolic—associated fatty liver disease; MASH, metabolic-associated steatohepatitis; HV, healthy volunteer; ATLL, adult T-cell leukaemia/lymphoma; CTLCL, cutaneous T-cell lymphoma; CLL, chronic lymphocytic leukaemia; DLBC, diffuse large B-cell lymphoma; MF, mycosis fungoides; pSil, pSilencer; PMIS, plasmid-based miRNA inhibitor system; n/a, not applicable.
Examples of miRNA-based therapeutics in clinical trials
Miravirsen
Miravirsen (formerly SPC3649), developed by Santaris Pharma A/S, was the first miRNA-targeted drug to enter in-human clinical trials. Miravirsen is a β-d-oxy-LNA-modified PS-ASO targeting the liver-specific miR-122 and proposed as an antiviral therapeutic for HCV genotype 1 infection. miR-122 regulates hepatic lipid metabolism and inflammation in hepatocytes and is essential for HCV stability and proliferation by binding to three sites in the 3′- and 5′-UTRs of the HCV genome. 102,103 Thus, the antiviral activity of miravirsen was tested in several preclinical studies both in vitro on hepatocyte culture models and in vivo on HCV-infected chimpanzee model. 104,105 Miravirsen moved forward with Phase II clinical trials testing safety and tolerability in multiple ascending dose studies (CT.gov ID NCT00979927, NCT01200420), 106,107 long-term safety and antiviral efficacy of miravirsen monotherapy (CT.gov ID NCT01727934), 107 co-administered with telaprevir (CT.gov ID NCT01727934, NCT02508090) or in combination with telaprevir plus ribavirin (CT.gov ID NCT01872936, NCT02452814) on null responders to standard-of-care therapy for HCV patients. Although miravirsen treatment resulted in undetectable levels of HCV RNA in 50% of test subjects for up to 14 weeks, viral rebound was observed due to novel point mutations in HCV binding sites resulting in acquired viral resistance to miravirsen. 104,108,109
RG-101
A second miR-122 antagomiR was developed by Regulus Therapeutics and tested in clinical trials in patients with chronic HCV genotype 1, 3, and 4 infection in a single subcutaneous administration at two test doses. 95 RG-101 is a PS-ASO conjugated to a multivalent GalNAc structure designed to enhance the uptake by hepatocytes. 95,110 This modification results in a consistent increase in the RG-101 potency with respect to the non-conjugated oligonucleotide. RG-101 treatment reduced HCV RNA levels for up to 76 weeks with no serious adverse events. 95,110 Further clinical trials tested the combination of RG-101 treatment with the oral agent GSK2878175, a pan-genotypic non-nucleoside NS5B polymerase inhibitor of HCV virus (EudraCT Number: 2015–001535-21; 2015–004702-42; 2016–002069-77). 111 Outcomes of the studies reported successful responses to combined therapy for most patients enrolled, one non-responder, and seven patients experienced viral relapse. 111
MRX34
miR-34a is the main member of the miR-34 family along with miR-34b and miR-34c discovered as natural p53-regulated tumor suppressor miRNAs. 112 It has been found to downregulate the expression of more than 30 oncogenes, and it is low expressed in a broad of tumor types. 113,114 The miR-34a mimic, MRX34, sponsored by Mirna Therapeutics, was proposed for the treatment of several advanced solid tumors, including primary liver cancer, lymphoma, melanoma, renal cell carcinoma, and non-small cell lung cancer (NSCLC). MRX34 was designed as a synthetic ds miR34a mimic encapsulated into a liposome-formulated nanoparticle (NOV40). 115 Phase I clinical trial testing MRX34 (CT.gov ID: NCT01829971) was prematurely terminated due to severe immune-related adverse toxicities leading to the death of four patients, causing the withdrawal of a second trial (CT.gov ID: NCT02862145) testing MRX34 and dexamethasone combined therapy for melanoma treatment. 116,117 Over the side effect, the study reported an efficient drug delivery to the tumors and dose-dependent repression of miR-34a target genes. Despite the liposomal carrier used to deliver the drug was excluded from being in charge of MRX34 toxicity, severe side effects experienced by patients during this trial have made it clear that a greater awareness of the delivery system and an accurate study of dose-dependent response were fundamental to go on further clinical trials.
TARGOmiR
Mesomir-1 clinical trial (CT.gov ID NCT02369198) tested TARGOmiR, a miR-16 mimic developed by Asbestos Diseases Research Foundation and proposed as 2nd or 3rd line treatment for patients with recurrent malignant pleural mesothelioma (MLM) and NSCLC. TARGOmiR exploited a more sophisticated delivery strategy based on EGFR-targeting EnGeneIC Dream Vectors (EDVs) system to precisely deliver miR-16 mimics into EGFR-expressing tumor cells. miR-16–1 clusters with miR-15a at chromosome 13q14.3, the most common genomic aberration in chronic lymphocytic leukemia (CLL). 118 miR-16/15 function as tumor suppressor targeting several oncogenes 119 and are downregulated in MLM. The mesomir-1 study reported a variable response rate, with only one patient of the 22 analyzed showing an objective response, probably due to the low dose of TARGOmiR. 120 However, TARGOmiR was well tolerated by patients, and only minor dose-dependent toxicities (anaphylaxis, inflammation, and cardiac events) were reported. 120
Lademirsen
Lademirsen is a ss chemically modified ASO targeting miR-21 provided by Regulus Therapeutics. miR-21 is upregulated in kidney diseases, modulating the tissue repair response following acute or chronic renal injury. 121 The Phase 2 HERA clinical trial (CT.gov ID NCT02855268) sponsored by Genzyme, a Sanofi Company, was designed to test the safety, tolerability, and efficacy of lademirsen in reducing renal failure in participants with Alport syndrome. The planned length of participation in the study was up to approximately 110 weeks. However, the company decided to prematurely stop the clinical trial due to the futility of interim results following lademirsen treatment compared with the placebo, despite no unexpected adverse events were identified. The analysis came from 24 patients who had completed the 24-week, double-blind treatment period.
AMT-130
AMT-130, an a-miRNA based on pri-miR-451 and delivered via an AAV5 vector, is currently under investigation in a challenging clinical trial for allele-nonselective silencing of HTT gene. The efficacy and safety of AMT-130 have been assessed in cultured human neurons and in multiple animal models, such as mice, non-human primates, and transgenic minipigs. 71,122,123 The U.S. Phase I/II clinical trial of AMT-130 (CT.gov ID NCT04120493) explores the safety, tolerability, and efficacy in 36 patients with early manifest Huntington’s disease (HD). 124,125 Patients were randomized to treatment with AMT-130 in single administration through MRI-guided, convection-enhanced stereotactic neurosurgical infusion directly into the striatum (caudate and putamen) or an imitation (sham) surgery. The European Phase Ib/II study of AMT-130 (CT.gov ID NCT05243017) enrolled 13 HD patients across two dose cohorts to assess the optimal drug dose and validate safety and efficacy. FDA recently granted UniQure Regenerative Medicine Advanced Therapy (RMAT) designation based on the promising 24-month interim clinical data for AMT-130 announced in December 2023, intensifying the cooperation with the FDA on regulatory approval planning.
TTX-MC138
Multiple studies have documented miR-10b as a biomarker of tumor invasion and metastasis. TransCode Therapeutics developed a miR-10b-targeting drug, TTX-MC138, as the lead therapeutic candidate for treating metastatic cancer overexpressing miR-10b. Preclinical studies showed long-lasting regression of metastases in murine models of metastatic breast cancer and pancreatic cancer upon TTX-MC138 treatment. TTX-MC138 consists of a dextran-coated IONP designed to access targets inside tumor cells, conjugated to a ss miR-10b-targeted ASO. Delivery and distribution of radiolabeled TTX-MC138 (TTX-MC138-NODAGA-Cu64) were assessed in a Phase 0 pilot microdose study in a subject with metastatic solid cancer (CT.gov ID NCT05908773). 101,126 At the end of May 2024, TransCode Therapeutics announced novel promising preliminary results from the patient dosed in the trial, reporting a reduction of 66% of miR-10b at 24 h post-treatment in the patient’s blood and evidence of TTX-MC138 accumulation at sites of metastases. Moreover, the FDA has recently authorized the company to proceed with its Phase 1/2, dose-escalation and dose-expansion study of TTX-MC138 in patients with advanced solid tumors (CT.gov ID NCT06260774).
RGLS8429
miR-17 has been found overexpressed in both murine and human Autosomal Dominant Polycystic Kidney Disease (ADPKD). miR-17 is part of a polycistronic cluster (miR-17∼92), which produces six individual miRNAs. The genetic ablation of miR-17∼92 cluster slows down disease progression in ADPKD mouse models, increasing interest in the development of drugs able to modulate miR-17 expression. RGLS8429 is the second-generation therapeutic candidate promoted by Regulus Therapeutics under Phase 1b clinical trial investigation (CT.gov ID NCT05521191). The first miR-17 inhibitor proposed by the company, RGLS4326, 110 has shown dose-limiting central nervous system (CNS) toxicity in mice and monkeys receiving high doses. 111,112,127,128 Like RGLS4326, RGLS8429 targets preferentially the kidney and demonstrates similar potency in inhibiting miR-17 in in vitro and in vivo studies without causing CNS toxicity. A pilot study on healthy volunteers confirmed RGLS8429’s safety and tolerability (CT.gov ID NCT05429073). Preliminary results announced at the end of June 2024, following the completion of tests on a cohort of 16 ADPKD patients, have been greatly encouraging about the potential efficacy of RGLS8429 and may represent a turning point toward a pivotal Phase 2/3 study of RGLS8429 under an Accelerated Approval regulatory pathway to be discussed with the FDA.
CDR132l
The miR-132/212 family has been shown to regulate several cardiac functions. 129 Indeed, over-activation of miR-132 modulates cardiomyocyte contractility driving adverse cardiac remodeling in the context of the heart failure (HF) pathological progression. 49 Cardior Pharmaceuticals developed CDR132L, the first generation of synthetic inhibitor targeting cardiac miR-132, currently under evaluation in HF-REVERT Phase 2 clinical study (CT.gov ID NCT05350969; EudraCT Number 2021–006040-27) 130 in patients with reduced left ventricular ejection fraction (≤45%) after myocardial infarction. CDR132L has already been tested for safety and efficacy at both preclinical (in large animal models, i.e., pigs 131 ) and clinical stage, in a Phase 1b study in chronic HF patients (CT.gov ID NCT04045405; EudraCT Number: 2019–001291-10). 132 A third clinical trial testing CDR132L efficacy on reverse cardiac remodeling in patients with cardiac hypertrophy (REMOD-REVERT; CT.gov ID NCT05953831) was withdrawn following Cardior acquisition from Novo Nordisk, effective starting from May 2024.
ABX464
ABX464 (Obefazimod) is Abivax’s most advanced small molecule drug candidate into Phase 3 clinical development for the treatment of moderately to severely active ulcerative colitis (UC). Specifically, ABX464 acts as a miRNA enhancer, inducing the selective splicing of a single long non-coding RNA to generate the miR-124, which in turn downregulates pro-inflammatory cytokines and chemokines. It has been reported a robust and long-lasting increase in miR-124 expression in colorectal biopsies and blood samples of UC patients upon ABX464 administration, suggesting broad potential as a novel anti-inflammatory agent. The long-term safety and efficacy profile of ABX464 in UC treatment will be assessed in the ABTECT program, which consists of two induction studies and a subsequent maintenance trial (CT.gov ID NCT05507203; NCT05507216; NCT05535946). 133,134 A Phase 2b trial in Crohn’s disease is also planned, with enrollment expected to start in the end of 2024 (CT.gov ID NCT03905109). 135 ABX464 has also been clinically evaluated in patients with moderate to severe active rheumatoid arthritis (CT.gov ID NCT03813199; NCT04049448), in HIV-infected patients (CT.gov ID NCT02735863; NCT02990325) 136 and in patients with COVID-19 (CT.gov ID NCT04393038).
CHALLENGES AND PROSPECTS OF miRNA-BASED THERAPEUTICS
In the last few years, miRNA-based therapeutics achieved significant advancements in preclinical research, and the field continues to evolve. However, while some of them have advanced to clinical trials, none have yet received approval from the regulatory agencies. Individual miRNAs have the ability to regulate a wide range of mRNA targets. Although this makes them promising candidates for therapeutic use, it also complicates the control of adverse effects seen in clinical trials. Notably, several candidates have been discontinued due to toxicity concerns, highlighting the need for a comprehensive risk assessment of miRNA therapeutics before any in vivo studies, aiming to minimize off-target effects and to avoid overdosing of miRNAs.
Key challenges include: Establishing Sensitivity, Specificity, and Selectivity: Ensuring miRNAs accurately target their intended mRNAs without affecting off-target genes. Mitigating Immunogenic Reactions: Reducing immune responses triggered by miRNA modifications and delivery systems. Enhanced Targeted Delivery Methods: Developing advanced delivery systems to efficiently and precisely direct miRNAs to target cells. Optimal Dosing: Determining the correct dosage to achieve therapeutic efficacy while minimizing side effects.
The advancement of biochemistry and bioengineering technologies has made it possible to address these obstacles. Recent preclinical and clinical trials have demonstrated higher safety, efficacy, and specificity of new synthetic RNA oligomers for miRNA mimics and antisense miRNA inhibitors. 11 The approval of numerous antisense, small interfering RNA, and mRNA-based therapeutics and vaccines has affirmed their potential and paved the way for applying RNA therapies for human disorders. New promising delivery systems have been developed to effectively and precisely deliver miRNA drugs. Various strategies can be employed to achieve appropriate dosing in therapeutic approaches. One strategy is the combined use of cooperating miRNAs to keep the therapeutic doses of individual miRNAs as low as possible. Notably, strategies that aim to decrease the activity of an endogenous miRNA in a specific tissue/cell type may be safer compared to those that lead to miRNA overexpression in different parts of the body. Approaches involving the adoption of vector-based expression systems designed to ensure physiologically adapted and tissue-specific expression levels using endogenous promoter sequences may overcome this issue.
Further studies are needed to clarify if immunogenic reactions can arise from viral transfer systems, as well as from modifications of miRNA-based therapeutics such as LNAs and a-miRNAs, miRNA interfering molecules, including small cell-permeable molecules, application systems like biodegradable 3D matrices, carriers such as functionalized metals, viral transfer systems, or biomolecule conjugates such as aptamers. Masking reactive components may potentially reduce these immunogenic reactions.
In conclusion, although the therapeutic potential of miRNAs for treating various diseases is clear, comprehensive risk assessment and ongoing research are crucial to establish their suitability for clinical applications. Despite challenges, the encouraging progress in miRNA therapeutics promises their increasingly impactful role in future medicine.
Footnotes
ACKNOWLEDGMENTS
AUTHORS’ CONTRIBUTIONS
S.B., M.V., and A.I. conceived the study and wrote the article. S.B. and M.V. contributed equally to this work and should be considered co-first authors. M.V. prepared figures; S.B. summarized the table. All authors contributed to the article editing and approved the submitted version.
AUTHOR DISCLOSURE
A.I. is a coinventor of the patent applications “miR-181 inhibitors and uses thereof” (WO2019202162A1). The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
FUNDING INFORMATION
This study was supported by Telethon Foundation and Italian Ministry of University and Research (