
Abstract
Corneal neovascularization (CoNV) is both a sight-threatening condition in and of itself and a major risk factor associated with corneal graft failure. Here, we determine the effectiveness of an adeno-associated viral vector (AAV)-based gene therapy targeting both hematic and lymphatic neovascularization in a murine model of severe CoNV. We first assessed the profile of transgene expression mediated by intrastromal injection of AAV2/8[Y733F] via longitudinal visualization of an enhanced Green Fluorescent Protein (eGFP) transgene and found that this serotype mediates a temporary (∼18 day) transduction of the corneal epithelium and sustained (≥148 day) transduction within the stroma. Constitutively expressed sFlt1 or sFlt4 were prophylactically delivered via intrastromal injection of AAV2/8[Y733F] vector at various intervals prior to aggressive induction of CoNV in a murine model. The extent of CoNV induced was quantified by fluorescein angiography and immunohistochemistry 17 days after induction. AAV2/8[Y733F]-CMV-sFlt1 was highly effective in the prevention of hemangiogenesis (HA) induced at 3, 28, and 210 days after intrastromal injection, but ineffective in the prevention of lymphangiogenesis. Two variants of AAV2/8[Y733F]-CMV-sFlt4 were ineffective in the prevention of angiogenesis when delivered alone, but combined delivery of AAV2/8[Y733F]-CMV-sFlt1 and AAV2/8[Y733F]-CMV-sFlt4 suggested a synergistic effect. Our results show that a single intrastromal injection of AAV2/8[Y733F]-CMV-sFlt1 is sufficient to protect against a robust stimulus for corneal HA over the long term. This technique could also be applied ex vivo to reduce the risk of failure in cases of “high-risk” corneal transplantation.
INTRODUCTION
The healthy cornea is maintained in an avascular state to preserve optical transparency and minimize light scatter. This avascularity is sustained by a complex balance of both pro- and antiangiogenic factors present within the cornea, 1,2 with soluble Vascular Endothelial Growth Factor (VEGF) receptors having been shown to play a key role. 3 In particular, sFlt1 and sFlt4 (also known as sVEGFR1 and sVEGFR3) are critical for maintenance of hematic and lymphatic avascularity, respectively. 4,5
Pathological corneal neovascularization (CoNV) can result from a wide range of different conditions—from extended contact lens wear to trauma and alkali burns, infections, parasitosis, and immunological disorders. Similarly, its severity may range from nonsight-threatening to blinding. The development of pathological blood vessels is the result of an unregulated and misdirected process that results in the blood vessels formed being functionally immature and lacking in structural integrity. Therefore, in addition to their inherent negative impact upon corneal transparency, such vessels are also often associated with lipid keratopathy, in which lipid leaks from the vessels into the surrounding tissue to form opaque deposits within the cornea. Pathology can be further complicated as the neovascular growth of blood vessels (hemangiogenesis—HA) is almost invariably accompanied by a similar growth of lymph vessels (lymphangiogenesis—LA). 6
In most cases, CoNV is the secondary result of a primary inflammatory insult. Inflammatory responses to injury or infection are associated with the cascading activation of numerous cytokine pathways within the cornea, initiated by IL-1 secretion from the cells of the corneal epithelium and infiltrating innate immune cells. 7,8 Many of these cytokines are potent pro-angiogenic factors themselves; IL1-β, for example, has been shown to be an angiogenic factor comparable in strength to VEGF-A that also leads to the widespread downstream upregulation of the VEGF pathway itself. 9,10 Such prolific cytokine activation disrupts the angiogenic homeostasis of the cornea leading to neovascularization. Furthermore, the pathogenesis may not be limited to neovascularization as upregulation of TGF-β can also result in the activation of stromal keratocytes leading to stromal fibrosis and scarring. 11 If these pathologies become severe enough to lead to corneal blindness, the only effective intervention to restore vision is corneal transplantation and, in this context, CoNV is also of clinical relevance.
The prognosis of penetrating keratoplasty under ideal conditions is excellent, with 5-year graft survival rates of up to 95–99%. 12 The immune privilege enjoyed by the cornea 13,14 reduces the need for tissue typing and means only minimal immune suppression is required. However, the presence of preexisting neovascularization in ≥2 quadrants of the recipient cornea is strongly associated with a higher risk of transplant rejection and consequent graft failure. 12,15 –18 This increased risk is thought to derive from corneal neovascular vessels compromising the cornea’s immune-privileged status 19 –23 by facilitating greater exposure of the alloantigens to the host immune system and ingress of immune effector cells into the graft.
Pharmaceutical treatments for neovascularization are most effective when applied shortly after the onset of inflammation, well before neovascularization has become established. In cases of infection, a careful balance must be struck between the use of topical steroid to control inflammation (and any associated neovascularization) and inhibition of an immune response that might otherwise facilitate removal of the pathogen. Once neovascularization is more established more direct treatments may be applied, including the direct ablation or occlusion of vessels by means of lasers, fine needle diathermy, 24 photodynamic therapy, 25 and, potentially, corneal collagen cross-linking. 26 More recently, the extensive clinical use of VEGF-binding therapeutics for the treatment of choroidal neovascularization in wet age-related macular degeneration has led to use of these drugs being directed toward CoNV both in the clinic and in animal models. 27 –29 Initial results from these studies are promising overall, but treatment regimens often involve repeated administration at relatively high doses.
In this study, we aimed to develop a single-dose gene therapy able to protect the cornea against an aggressive pro-angiogenic insult, using an adeno-associated viral vector (AAV) to deliver transgenes able to modulate the VEGFR1 and VEGFR3 pathways (shown in Supplementary Fig. S1) to ameliorate HA and LA, respectively. Such a therapy could potentially be applied to donor corneas ex vivo prior to transplantation to improve clinical outcomes in cases of “high-risk” corneal grafts.
MATERIALS AND METHODS
Animals
All mice used were of the wild-type strain C57BL/6J. All mice were female, adult (6+ weeks), and maintained under a 12-h light–dark cycle. All experiments were approved by the local Institutional Animal Care and Use Committees (UCL, London, United Kingdom) and conformed to the guidelines on the care and use of animals adopted by the Society for Neuroscience and the Association for Research in Vision and Ophthalmology (Rockville, Maryland). Prior to all surgeries, mice were anesthetized with a single intraperitoneal (IP) injection of a mixture of Domitor (1 mg/mL medetomidine hydrochloride), ketamine (100 mg/mL), and water in a ratio of 5:3:42. Subsequent to all surgeries, anesthesia was reversed and a 1% chloramphenicol ointment (FDC International) was applied topically to the cornea.
Human tissue
Whole human corneas from deceased multi-organ donors unsuitable for transplantation were obtained from Moorfields Eye Bank, London, United Kingdom, after consent for use in research was obtained from relatives. Specimen collection and arrangements for consent were managed in accordance with the Declaration of Helsinki and approved by the local research ethics committee. Corneas were maintained in organ culture in 25 mL of Culture Medium I (Biochrom AG) in a tissue culture incubator at 37°C/5% CO2. Media were refreshed weekly.
Recombinant AAV genomic plasmids
All plasmids encoding recombinant AAV (rAAV) genomes used in this study shared a common backbone. All therapeutic constructs shared with the following features: AAV 2/2 ITRs Expression driven by the ubiquitous CMV promoter SV40 polyadenylation site
Our sFlt1 construct encoded the murine Flt1 CDS (NCBI Reference Sequence: NM_010228.4) truncated at nt 2499, thus encoding the first 13 exons. Our initial “sFlt4” transgene was a synthetic soluble version of murine Flt4 created by mimicking the splicing of sFlt1, truncating the Flt4 CDS (NCBI Reference Sequence: NM_008029.3) at nt 1513, a position equivalent to the end of exon 10, truncating the protein at residue 474. However, it is now known that the cornea endogenously expresses a soluble form of Flt4. 5 This endogenous version differed from our synthetic version in that its CDS included exon 11 and a small portion (27 bp) of the Flt4 intron 11-12. We therefore prepared our “sFlt4[Ex11+]” construct to express this endogenous form. As controls we also produced constructs encoding eGFP and a “NULL” genome. The NULL plasmid lacked an exogenous promoter and encoded an incomplete 3444 bp CDS without a start codon.
rAAV vector preparation and titer
rAAV vectors were produced via triple transient transfection. All AAV made was of either the 2/8 or 2/8[Y733F] serotypes. The plasmid construct, AAV serotype-specific packaging plasmid, and helper plasmid were mixed in a ratio of 1:1:3 at 20 µg total DNA per mL of Dulbecco's Modified Eagle Medium (DMEM), then mixed with polyethylenimine (Polysciences Inc.) to a final concentration of 50 mg/mL and incubated for 10 min at room temperature to form transfection complexes. These were added to HEK293T cells at 50 µg DNA per 175 cm2 and left for 72 h. The cells were collected, concentrated, and lysed by freeze–thaw (4×) in TD buffer to release the vector. AAV was purified from this lysate by affinity-based Fast Protein Liquid Chromatography (FPLC) with an AVB Sepharose column (GE Healthcare), eluted with 50 mM glycine pH 2.7 into 1 M Tris pH 8.8. Vectors were washed in 1× Phosphate Buffered Saline (Magnesium Chloride, Potassium Chloride) (PBS-MK) and concentrated to a volume of 100–150 µL using Vivaspin 4 (10 kDa) concentrators. Vector genome (vg)-based titers were determined by quantitative real-time PCR (qPCR) using an Inverted Terminal Repeat (ITR) binding assay as previously described. 30
Intrastromal injections
Intrastromal injections (mice, in vivo) were performed under direct observation with an operating microscope. Vector solution was injected via a 34-gauge hypodermic needle mounted on a 5 μL Hamilton syringe (Hamilton). The needle tip was positioned (bevel up) about a half to a quarter of the corneal radius inward from limbus and penetrated the cornea in this position. The needle was then pushed into the cornea, taking care to remain within the stroma. Once the needle tip was positioned centrally, a volume of vector solution was injected. Unless otherwise stated, the injection volume was 2 µL. The cornea was observed to become opaque in the area of the injection (usually extending over a majority of the cornea); this stromal hydration completely resolved 24 h postinjection.
Intrastromal injections (human, ex vivo) were performed by securing the tissue on a plastic artificial anterior chamber (Barron). An approximately physiological “intraocular” pressure was obtained by injection of dPBS into the device. Intrastromal injection of vector or control solution was then performed using a ½ cc tuberculin syringe with attached 27G × ½ in. needle and an operating microscope. Injections then proceeded identically as in the mice but with an increased injected volume of 50 μL.
Induction of corneal neovascularization by placement of corneal sutures
Corneal sutures were placed under direct observation with an operating microscope. The eye to be sutured was dilated by topical application of 1% tropicamide (Minims®, Bausch & Lomb UK Ltd), mildly prolapsed, and secured in position with an elasticated vinyl drape. Suturing was performed using a 6.5 mm, 3/8 circle, spatulated, advanced micro-point double needle with attached 11-0 nylon (Ethilon, Johnson & Johnson Medical). Three sutures were placed in a trefoil configuration, each sited by penetrating the full thickness of the cornea paracentrally, passing the needle tip into the anterior chamber, while avoiding the iris, and exiting as close to the limbus as possible. The suture was then drawn through needle track, tied, and secured with a 2-1-1 knot, taking care not to overtighten and distort the cornea. Sutures were not buried, and ends were left at about 2 mm to aid with removal. A schematic and image showing suture placement can be seen in Supplementary Fig. S2.
Fluorescein angiography (SLO-FA)
Prior to imaging, mice were anesthetized to minimize motion blurring and then injected intraperitoneally with 0.2 mL of 2% fluorescein (in sterile water). At the point of injection, a timer was started, and imaging then began immediately. The right eye was always imaged first, followed by the left, in total imaging took no more than ∼5 min.
Fluorescein angiographic images were acquired by scanning laser ophthalmoscopy (SLO). The instrument used was a Spectralis HRA2 with a 55° lens (Heidelberg Engineering), modified for use with animals with increased 488 laser power. All images were taken using the 25° angle setting. Infrared (790 nm) images were taken using a power setting of 75%, and 30 images were averaged and normalized. Green Fluorescent Protein (GFP) (488 nm) images were taken using a variable power setting (chosen to minimize noise), and 30 images were averaged and normalized. A single infrared image was taken to visualize the positioning of the eye. Fluorescein expression over an entire cornea was visualized by compositing multiple 488-channel images from multiple focal planes.
Corneal fluorescent imaging
Prior to imaging, mice were anesthetized to minimize motion blurring. Corneal GFP or background fluorescence was recorded using a slit lamp (SC-16, Keeler) modified both for animal use and with an excitation filter optimized to GFP (λ = 475 ± 25 nm). Corneal refractive power was neutralized by placing a 5 mm coverslip on the cornea covered with a coupling medium solution (Viscotears, Novartis Pharmaceuticals). Images were acquired using an attached DC500 digital camera and associated software (Leica). All images were taken at 25× magnification, and the following exposure times were used: Bright field: 0.2 s; GFP: 30 s.
Immunohistochemistry and histology
For corneal flat mounts, eyes were harvested postmortem, and corneas were removed by circumferential excision posterior to the limbus. Tissue was fixed in 4% paraformaldehyde for 1 h before four evenly spaced radial cuts were made with a scalpel. Tissue was then counterstained with 4′,6-diamidino-2-phenylindole (DAPI) and mounted with DAKO fluorescent mounting media. For corneal sections, eyes were harvested postmortem, and globes were punctured posterior to the limbus using the tip of a 30G needle and then fixed in 4% paraformaldehyde for 1 h before embedding in optimal cutting temperature medium. Twenty micrometer cryosections were cut in sagittal orientation, counterstained with DAPI and mounted with DAKO fluorescent mounting media (DAKO, S3023, Denmark). All images were acquired by confocal microscopy (Leica DM5500Q). The immunohistochemistry (IHC) of both hematic and lymphatic vessels was achieved largely according to protocol outlined in Cao et al. 31 using the following primary antibodies in Table 1.
Antibody Reagents for Vascular and Cellular Marker Detection
Quantitative analysis of images
All analyses were masked to the observer and proceeded according to the following basic principles, applied in sequence. Define a region of interest (ROI) for analysis. Apply an arithmetic filter (pixel intensity threshold) to separate signal from background. Quantify the pixels representing signal within the ROI. Correct for any variability in size of the ROI between samples (divide by area).
For corneal fluorescence images, ROI was the entire cornea defined subjectively from a brightfield image. Fluorescence images were converted to grayscale, and a threshold of 0–80 (out of 255) was applied. Pixels were quantified by ImageJ’s integrated density measurement. 32 For SLO-FA images the ROI was the entire cornea defined subjectively from an infrared image. The threshold was set subjectively on an experiment-by-experiment basis, and pixels were quantified using GNU Image Manipulation Program (GIMP, open source). For immunohistological flat mounts of vascular staining images, the ROI was the entire cornea delimited by the staining of limbal vascular arcades. Fluorescent signal from IHC proved more variable, necessitating the threshold to be set subjectively on an image-by-image basis. Pixels were quantified using GIMP (open source). The CD31 antibody supplied by eBiosciences was found to cross-react and stain both hematic and lymphatic vessels. CD31 pixel quantification for this antibody therefore proceeded as follows: [Corrected CD31+ pixels] = [CD31+ pixels] − [LYVE1+ and CD31+ colocalized pixels], an example of this correction is shown in Supplementary Fig. S3. For immunohistological flat mounts assessing eGFP transgene expression, the ROI was a 480 × 960 × 10 µm region extending from the central cornea toward a randomly determined quadrant. Z-stack images were maximally projected, and the pixel intensity threshold applied was 25–255 in all cases.
Statistics, randomization, masking
All statistical tests were performed with GraphPad Prism version 10.4.0 for Windows (GraphPad Software, Boston, Massachusetts, www.graphpad.com).
Within a cohort where no more than two groups were compared at a time the test applied in all cases was a nonparametric t-test (Mann–Whitney U test). In cases where multiple, replicate cohorts were performed, Fisher’s combined probability test was applied to generate a combined p value. Within an experiment where two groups were compared over time, a two-way repeated measures analysis of variance with the Greenhouse–Geisser correction was performed.
In all treated cohorts vector identity (intervention or control) was masked prior to injection and was not unmasked until all analysis was complete. In addition, which eye (OD or OS) received which vector was randomized on a per animal basis.
RESULTS
Intrastromal injection of AAV2/8[Y733F] results in widespread, long-lasting transgene expression within the murine stroma
The potential efficacy of a gene therapy is heavily dependent upon the spatiotemporal pattern of transgene expression that the chosen vector can mediate. We therefore assessed this property for AAV2/8[Y733F] following intrastromal injection in mice by imaging a fluorescent transgene product longitudinally in vivo. The serotype AAV2/8[Y733F] was selected based on our previous work 33 showing it to mediate the highest and most reproducible level of transgene expression within the corneal epithelium of the six serotypes compared. A cohort of four mice received 0.5 µL intrastromal injections of either AAV2/8[Y733F]-CMV-eGFP (1.8 × 109 vg/eye, n = 5) or control vector AAV2/8[Y733F]-NULL (1.8 × 109 vg/eye, n = 3). Transgene expression was then assessed by longitudinal fluorescent imaging in vivo between 7 and 148 days postinjection (DPI), and images were processed for integrated density analysis. Representative processed images at each time point are shown in Fig. 1A for an eGFP vector-injected cornea and in Fig. 1B for a control vector-injected cornea. Corneal eGFP fluorescence plotted over time is shown in Fig. 1C. Fluorescence was maximal at 7 DPI and decreased before plateauing around 28 DPI and was maintained significantly above controls until 61 DPI. Subsequent histological analysis (Fig. 1D) revealed that the peak in eGFP expression arises from the transduction and then turnover of the corneal epithelium (as shown previously 33 ) and that stromal expression of eGFP transgene is maintained until at least 180 DPI (Fig. 1Diii). In our previously published dataset, 33 however, we did not analyze stromal transgene expression. This dataset was therefore reanalyzed to compare the stromal transgene expression mediated by AAV2/8[Y733F] with the more commonly used AAV2/8 at 8 DPI. In agreement with the epithelial data, AAV2/8[Y733F] was found to mediate approximately 4× greater eGFP transgene expression than AAV2/8 (Supplementary Fig. S4C, Mann–Whitney U test, p = 0.0022). Co-localization of eGFP transgene expression with CD34+ staining indicates transduction of keratocytes (Supplementary Fig. S5).

Time course of corneal transgene expression following murine intrastromal injection of AAV2/8[Y733F]-CMV-eGFP.
Intrastromal injection of AAV2/8[Y733F]-CMV-sFlt1 effectively and reproducibly prevents hemangiogenesis induced by suture placement
To determine the effect of intrastromal injection of AAV2/8[Y733F]-CMV-sFlt1 in a model of severe CoNV by suture placement, disease induction was timed so that the drive for maximal neovascularization coincided with the peak of transgene expression observed in Fig. 1C. The experimental schedule is outlined in Fig. 2A.
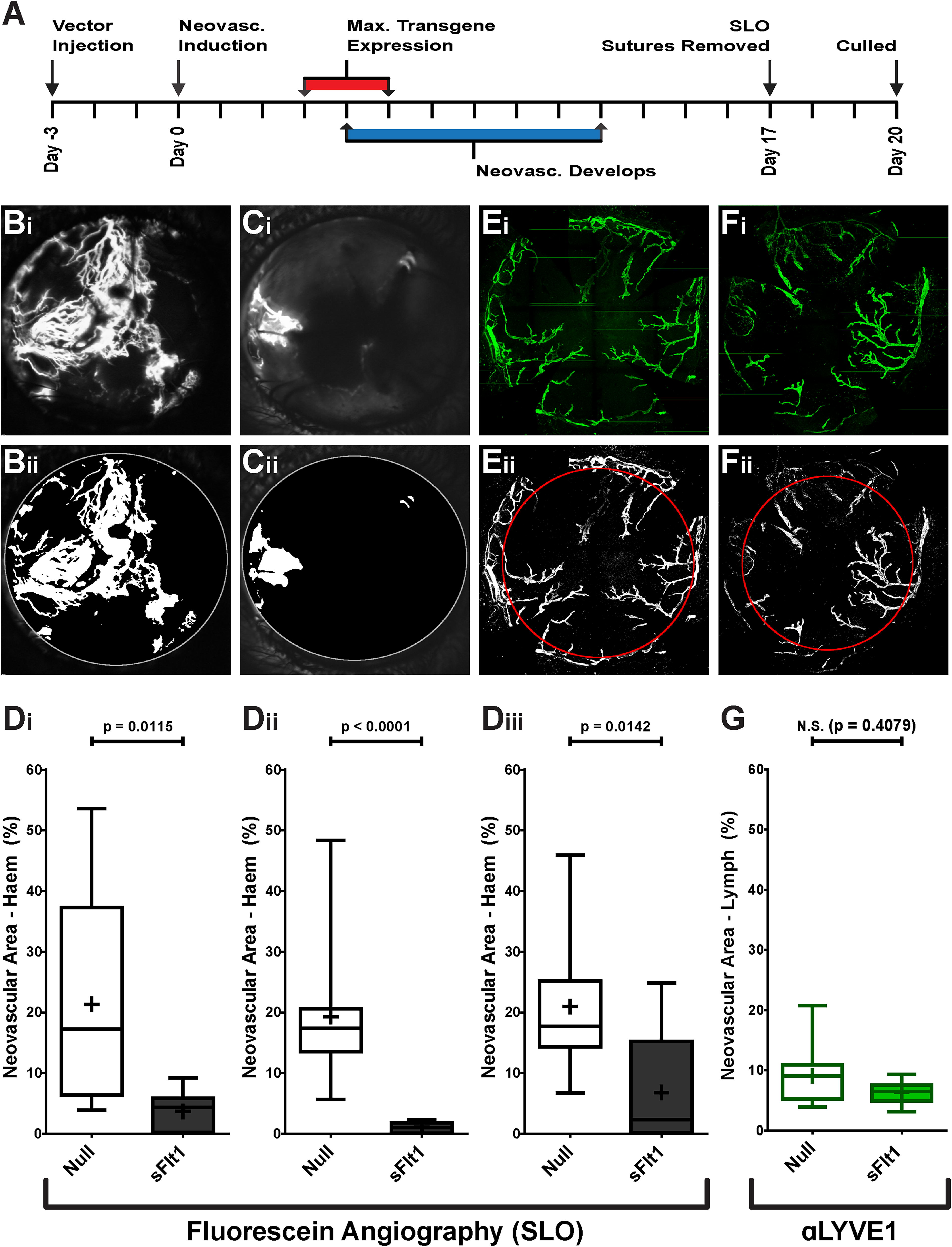
Effect of AAV2/8[Y733F]-CMV-sFlt1 on induced HA and LA over 17 days.
In each experiment, a total of 10 animals received intrastromal injection of AAV2/8[Y733F]-CMV-sFlt1 in one eye and AAV2/8[Y733F]-NULL in the other. Injection volume was 2 µL and vector dose was titer-matched to 2.4 × 109 vg/eye. Vector identity was masked prior to injection, and eyes were randomized to receive either the NULL vector or intervention. Representative raw and processed SLO-FA-derived images obtained at 17 days after suture placement are shown in Fig. 2B for NULL-injected eyes and Fig. 2C for sFlt1-injected eyes. Vector identity was only unmasked after completion of analysis. In three independent experiments, intrastromal injection of AAV2/8[Y733F]-CMV-sFlt1 resulted in a highly significant (Mann–Whitney U tests) reduction in the severity of induced HA (Fig. 2D) with a combined p value of 1.33 × 10−6 (Fisher’s combined probability test).
In one of these cohorts, the effect of AAV2/8[Y733F]-CMV-sFlt1 injection on induced LA was also assessed. At 20 DPI the animals were culled and IHC performed on corneal tissue for the lymphatic endothelial cell marker LYVE1 before being flat-mounted and imaged by confocal microscopy. Representative examples of raw and processed images are shown in Fig. 2E (NULL-injected) and Fig. 2F (sFlt1-injected). Intrastromal injection of AAV2/8[Y733F]-CMV-sFlt1 resulted in no significant reduction in LA (Fig. 2G).
Lymphangiogenesis induced by suture placement is not prevented by intrastromal injection of AAV2/8[Y733F] overexpressing sFlt4 or sFlt4Ex11+
The inability of sFlt1 overexpression to prevent LA is not unexpected, as it is not known to act directly upon the VEGFR-3 signaling pathway. We therefore determined whether intrastromal delivery of AAV2/8[Y733F] incorporating transgenes that do directly regulate this axis would be effective in ablating LA after suture placement. All experiments proceeded exactly as described in the previous section according to the same schedule. HA was assessed both by SLO-FA and IHC for the endothelial cell marker CD31. LA was assessed by IHC for LYVE1. Example of IHC for both markers can be seen in Supplementary Fig. S6. Vector dosages for these experiments (study and NULL control) were matched to 1.1 × 1010 vg/eye.
We determined the effect of both a synthetic sFlt4 and of an sFlt4 known to be endogenously expressed in the cornea, sFlt4[Ex11+], 5 after delivery by intrastromal injection of AAV2/8[Y733F]. Neither protein resulted in a significant change (Mann–Whitney U test) to the area of HA, as determined by SLO-FA (Fig. 3A) or IHC (Fig. 3B). The severity of induced LA was similarly unaffected (Fig. 3C).
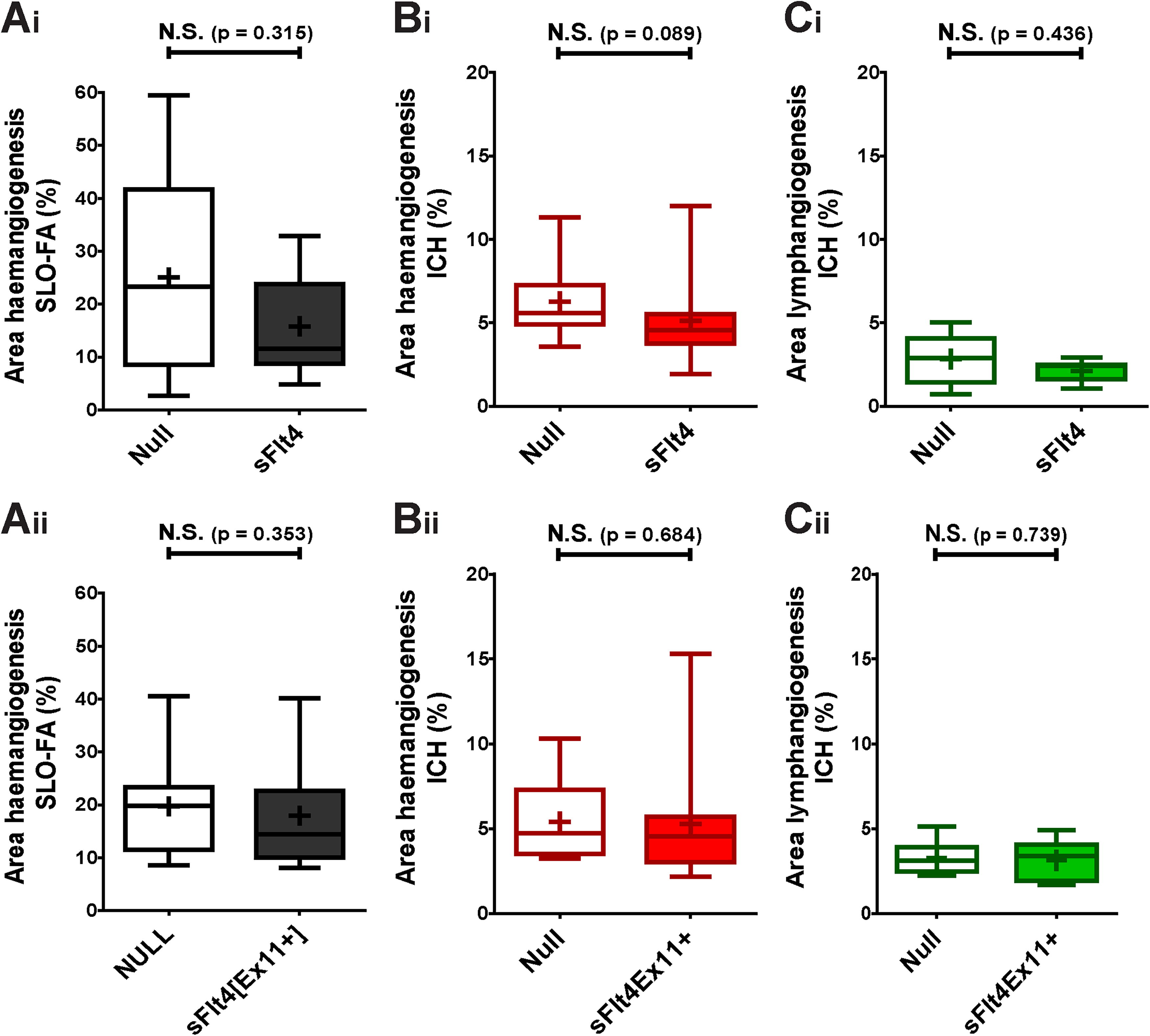
Effect of AAV2/8[Y733F]-CMV-sFlt4 and AAV2/8[Y733F]-CMV-sFlt4[Ex11+] on induction of HA and LA over 17 days.
Combined expression of sFlt1 and sFlt4 may act synergistically to attenuate induced lymphangiogenesis
As it has been suggested that the VEGFR1 pathway may indirectly impact that of VEGFR3, 34 we assessed whether AAV2/8[Y733F]-CMV-sFlt1 and AAV2/8[Y733F]-CMV-sFlt4 might act synergistically by mixing and co-injecting both vectors at a dose of 1 × 109 vg/eye each. In the first experiment, co-injection of the two vectors resulted in a significant reduction in HA (Fig. 4Ai–Bi) and LA (Fig. 4Ci, p = 0.023) in comparison with control (AAV2/8[Y733F]-NULL at 2 × 109 vg/eye). To confirm this novel finding, we repeated the experiment. On this occasion, we again observed a significant reduction in HA as determined by SLO-FA (Fig. 4Aii, p = 0.028). However, results obtained from IHC staining for CD31 did not quite reach significance (Fig. 4Bii, p = 0.054). Nor in this repeat did we see any significant reduction in induced LA (Fig. 4Cii, p = 0.397). However, applying Fisher’s combined probability test to these two experiments generates highly significant differences in regard to HA, as analyzed by SLO (p < 0.0001) and IHC (p < 0.0001), and finds the difference in LA to be close to significance (p = 0.052).
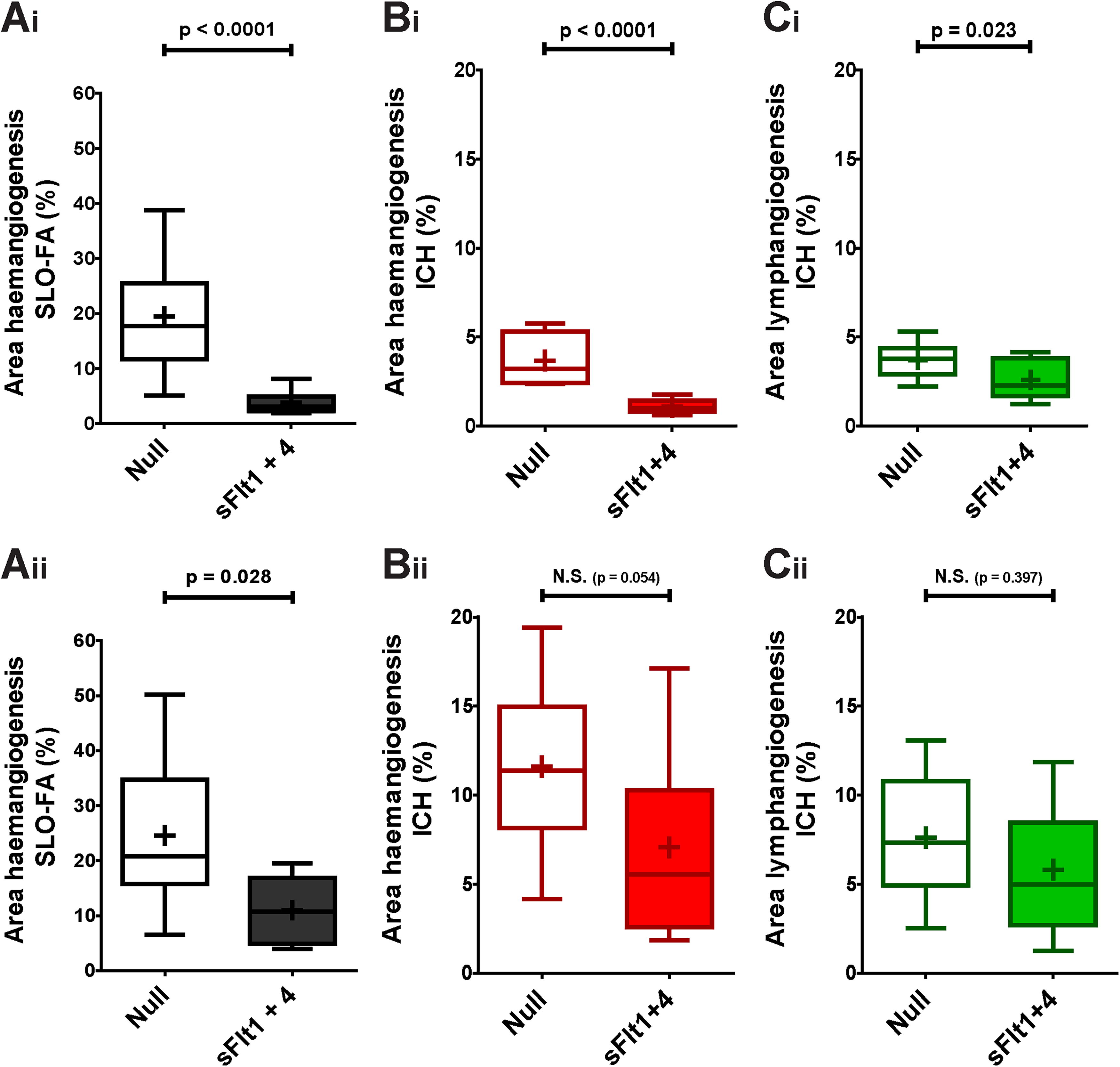
Combined effect of AAV2/8[Y733F]-CMV-sFlt1 and AAV2/8[Y733F]-CMV-sFlt4 upon induction of HA and LA over 17 days.
Intrastromal injection of AAV2/8[Y733F]-CMV-sFlt1 provides long-lasting protection against induction of hemangiogenesis
Previous experiments were timed such that the peak of transgene expression (as seen in Fig. 1) approximately coincided with the onset of HA in the suture model. After this initial peak, however, transgene expression is maintained over the long term at lower level within the stroma (Fig. 1D). We therefore determined whether this lower, sustained level of transgene expression was protective against induction of HA, leaving 4 weeks between vector injection (dose: 2.4 × 109 vg/eye) and induction of neovascularization (Fig. 5Ai). Plateau levels of sFlt1 proved highly effective in mediating a reduction in induced HA (Fig. 5Bi) but not LA (Fig. 5Ci). Further extension of the interval between vector injection and induction of neovascularization to 6 months (Fig. 5Aii) again resulted in a significant reduction in induced HA (Fig. 5Bii), with no effect on LA (Fig. 5Cii).
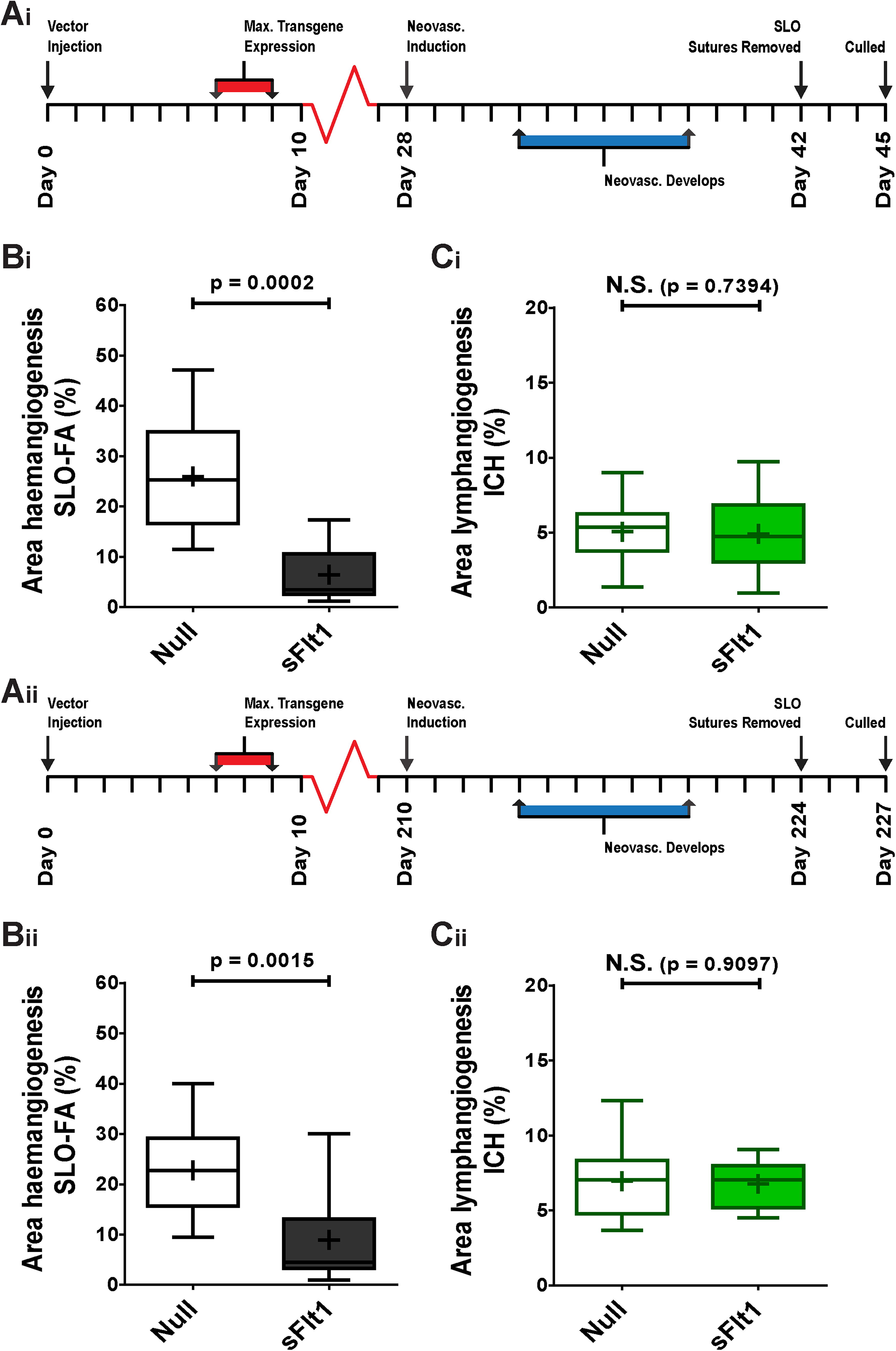
Effect of AAV2/8[Y733F]-CMV-sFlt1 upon induction of HA and LA over 42 days and over 224 days. (
Intrastromal injection of AAV2/8[Y733F]-CMV-eGFP results in stromal transgene expression within the human cornea ex vivo
Having determined that intrastromal injection of AAV2/8[Y733F]-CMV-sFlt1 is highly effective in the short- and long-term suppression of HA in a robust murine model of CoNV, we sought to determine the potential for translation to human corneal transplantation. We examined the transgene expression profile in human tissue ex vivo by injecting the vector intrastromally into human corneas harvested for the purposes of corneal transplantation or research. Injection volume was increased 25-fold to 50 µL to partially compensate for the larger human corneal volume. Vector dose was 1 × 1011 vg/cornea. At 7 DPI of AAV2/8[Y733F]-CMV-eGFP, extensive stromal transgene expression was seen within the area of injection (Fig. 6A–B); however, in contrast to the mouse data, no expression was seen in the epithelium, consistent with the devitalization of corneal epithelium observed in donor corneas stored in organ culture medium. 35
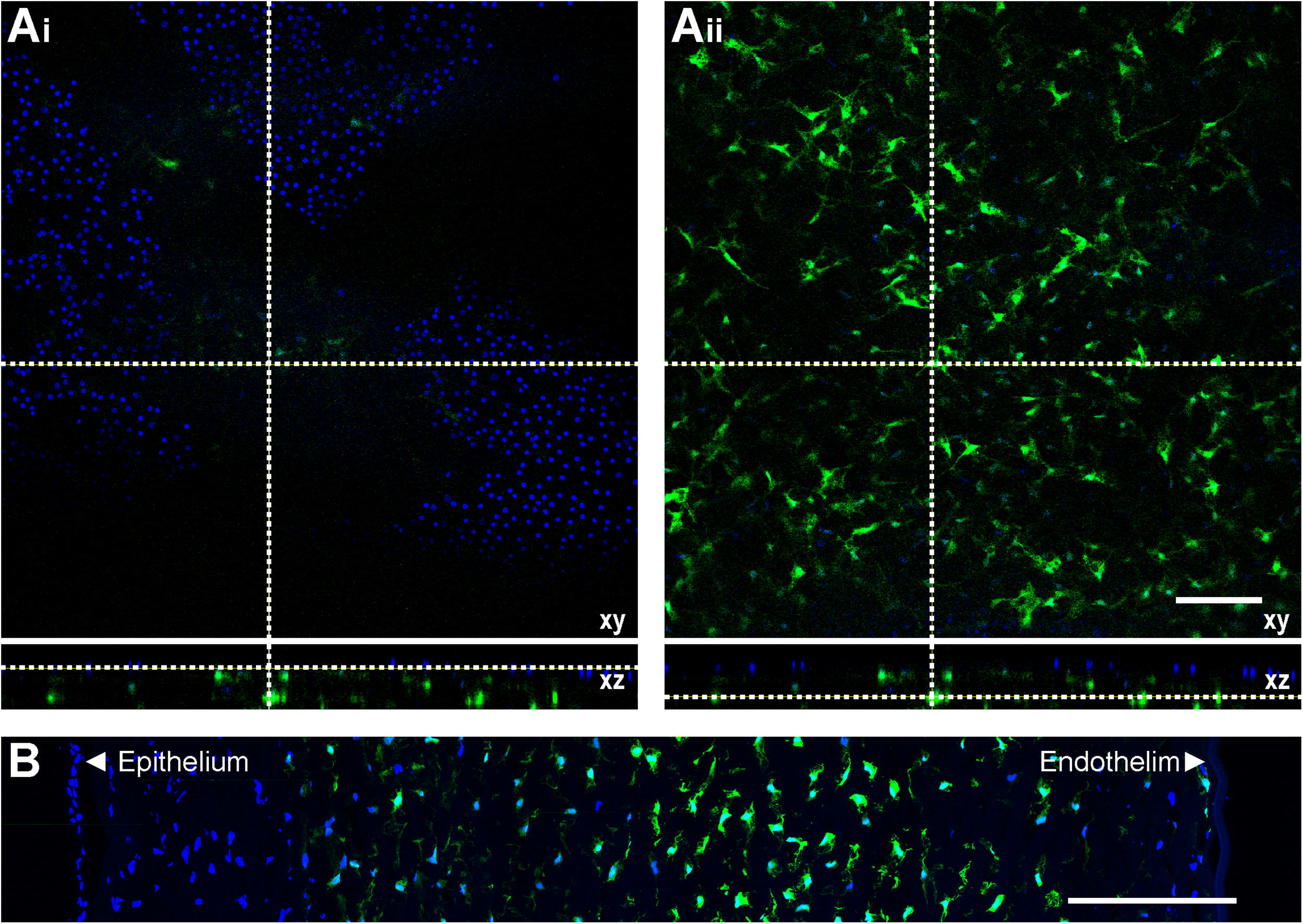
Transgene expression mediated by AAV2/8[Y733F] in a human cornea ex vivo.
DISCUSSION
CoNV, specifically HA, has been established as a key risk factor for corneal transplant rejection and failure for many years, 18 and an approach to mitigate this risk would be of significant clinical value. In this study, we have shown that intrastromal delivery of AAV2/8[Y733F]-CMV-sFlt1 is both highly effective and highly reproducible in ablating HA within a severe model of suture-induced CoNV. This result is in keeping with the wealth of published data supporting the effects of sVEGFR-1/sFlt1 in maintaining corneal avascularity and the effects of VEGF-trap on corneal transplant survival. 4,29,36 We demonstrated that the transduction of corneal stromal cells (CD34+, likely keratocytes) with an AAV2/8[Y733F] vector is sustained over the long term in the murine corneas, with transgene expression observed to occur for at least ∼6 months. Stable reporter transgene expression following intrastromal delivery of AAV2/8 has been observed to occur previously for ≥17 months in the murine cornea and 8 weeks in donor human corneas. 37 Furthermore, we found that this long-term transduction of the corneal stroma with AAV2/8[Y733F]-CMV-sFlt1 remained effective in suppressing HA even 6 months after vector administration. Interestingly, it has been previously shown in the cornea 37,38 and elsewhere 39 that rAAV-CMV based vectors can increase their transgene expression in response to subsequent inflammatory stimuli. It is possible, therefore, that sustained effectiveness we observed was enhanced by such an upregulation. Ultimately, the duration of effectiveness may be limited by keratocyte turnover, but our results indicate high translational potential—including the possibility of undertaking pretreatment of donor tissue prior to transplantation. While we only assessed a prophylactic treatment paradigm in this study, further work could also investigate the effectiveness of this gene therapy applied as treatment postinjury.
Given the effectiveness of our AAV-mediated sFlt1 upregulation in the murine cornea and the role of sFlt4 in maintaining corneal avascularity, 5 we next sought to determine whether such an upregulation of sFlt-4 might similarly prevent LA. 5,21,40 However, in contradiction to previously published work, 5,36,41 we found the expression of sFlt4 alone to be ineffective in suppressing the growth of lymphatic (and hematic) vessels. The exact reasons for this discrepancy are unclear, but several differences between our approach and the previously published work are notable. In this study, we applied AAV-mediated gene therapy, whereas previous approaches have utilized either naked plasmid-mediated gene therapy 5 or repeated administration of recombinant protein. 36,41 Such differing approaches would also very likely result in different kinetics and bioavailability of exogenous sFlt4 within the cornea. It is also likely that the modifications we made to the suture-induced CoNV model in our study (shown in Supplementary Fig. S2) represent a more severe inflammatory insult in comparison with how it is more usually applied, 41 –44 in particular, full-thickness penetration of cornea rather than intrastromal placement. Furthermore, we applied the suture model within C57BL/6J mice rather than in the more commonly used BALB/C strain. BALB/C mice have previously been categorized as “low responders” to pathological lymphangiogenic stimuli and may exhibit upregulation of antilymphangiogenic factors independent of VEGFR-2 and VEGFR-3 signaling. 45 Another possibility is that while the pathways involved in HA are highly conserved, there is more redundancy in the pathways associated with pathological LA, so that sequestration of VEGF-C and VEGF-D alone is insufficient to ablate it. Our evidence that combined AAV-mediated upregulation of both sFlt1 and sFlt4 simultaneously may be more effective in ablating LA than either approach in isolation would support this hypothesis.
The canonical VEGF-A pathway is not generally considered to directly contribute to LA. However, VEGF-A has been shown to be the primary mediator of inflammation-induced LA in the context of a sustained inflammatory insult to the skin, 46 and in the cornea VEGF-A has been shown to be lymphangiogenic if delivered at higher doses. 47 Indirect effects of the VEGF-A pathway in facilitating LA have also been demonstrated, such as driving the recruitment and activation of pro-lymphangiogenic macrophages. 48 Reviews of the literature 49 –52 suggest that while the canonical VEGF pathways (as shown in Supplementary Fig. S1) are broadly well supported, they are supplemented by a wide range of noncanonical interactions, 50,51,53 and the system as a whole is complex, adaptable, and intricately regulated. Factors such as mechanosensory context, 54 immune/inflammatory status, 55 and VEGF ligand bioavailability 56 all interact to modulate angiogenesis. Further work is required to dissect this complex pathway within the context of the cornea, whose avascular nature already marks it out as an atypical tissue. A comprehensive, quantitative analysis of VEGF ligands within cornea within the context of sequestration by gene therapy (as applied in this study) would be an appropriate first step in such an investigation, revealing any compensatory mechanism at the level of the VEGF ligands or implicating more complex noncanonical or downstream segments of the VEGF pathways for further study.
The work presented here has shown that AAV-mediated corneal gene therapy to be a robust and effective approach to ablate corneal HA, adding further to the growing body of translational work in this area. Other preclinical studies have investigated a wide range of transgenes (VEGF modulators, 40,57 –59 angiogenic inhibitors, 60 –62 decorin, 63 immune modulators, 64 and miRNAs 65 ), different routes of administration (subconjunctival, 60,61,65 intrastromal, 57,64 or intracameral 59 ), different AAV serotypes (AAV2, 57 AAV5, 63 AAV8, 57 AAV8G9, 64 or AAVrh10 65 ), and angiogenic models (alkali burn, 57,59 –61,64 –66 corneal suture, 57 or corneal pocket 63 ). While some of these other studies have demonstrated sustained transgene expression, 57,60,64 to date the therapeutic efficacy against a delayed angiogenic insult has not been investigated, nor has the AAV serotype AAV2/8[Y733F] been applied to corneal gene therapy.
Any clinical application of gene therapy must be undertaken with safety as a primary outcome, and the potential immunogenicity of the vector is a major consideration, one which might also limit potential efficacy. 67 The eye retains a degree of immune privilege that limits the immunogenicity of AAV, with subretinal delivery appearing to drive less inflammation than intravitreal injection. 68 Less is known about the immunogenicity of AAV in the cornea, but if comparable with the retina, it is likely to be time-limited and vary by age, sex, previous wild-type AAV exposure, and dose. 69 We would propose the approach presented here to be best suited to the pretreatment of donor corneal buttons for transplantation. This would limit the direct exposure of the recipient to the vector, and posttransplantation any immunogenicity in the cornea might be expected to be limited to the first few weeks posttransplantation and so very likely suppressed by the frequent application of topical steroid during this period. While further research is clearly required to determine the immunogenicity of AAV in the cornea and its implications for clinical application, we would propose that this strategy holds promise for reducing corneal transplant rejection in human subjects, for which CoNV is a major risk factor.
Footnotes
ACKNOWLEDGMENT
Ms. Selina Azam provided technical support in relation to viral vector production.
AUTHORS’ CONTRIBUTIONS
M.B.: Conceptualization, methodology, investigation, validation, formal analysis, writing—original draft, visualization, and writing—review and editing. S.R.: Conceptualization, methodology, investigation, validation, visualization, writing—review and editing, and funding acquisition. D.F.P.L.: Conceptualization, methodology, investigation, and supervision. A.J.S.: Conceptualization, methodology, writing—review and editing, and supervision. R.A.P.: Visualization and writing—review and editing. R.R.A.: Conceptualization, methodology, writing—review and editing, supervision, project administration, and funding acquisition.
AUTHOR DISCLOSURE STATEMENT
S.R. receives consultancy fees from Sanofi.
FUNDING INFORMATION
This work was supported by a grant from Fight for Sight to S.R.
SUPPLEMENTARY MATERIAL
Supplementary Figures
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.