
Abstract
Recombinant adeno-associated viruses (rAAVs) are widely used viral vectors in human gene therapy. However, DNA impurities, such as plasmid DNA and host cell DNA, remain a significant quality control concern for final products. Our study examined purified rAAV1-ZsGreen1, rAAV2-ZsGreen1, rAAV5-ZsGreen1, and rAAV6-ZsGreen1 samples and found that they contained 0.69–3.27% DNA impurities derived from three plasmids, as detected by droplet digital PCR. These plasmid-derived impurities primarily consisted of those derived from the pAAV plasmid (≥98.88%), with small amounts of pRC1, pRC2mi342, pRC5, or pRC6 (≤0.91%), and pHelper (≤0.21%) plasmids. To determine the DNA strand form of these impurities within the capsids, we used two different DNases with distinct substrate specificities. The extracted DNA impurities from the rAAV samples exhibited high sensitivity to nuclease P1 but not to lambda exonuclease. Similarly, host cell DNA encapsulated within the capsids revealed similar sensitivities to the nucleases. These findings indicate that DNA impurities derived from the plasmids and host cell DNA are encapsulated into rAAV capsids as single-stranded DNA, likely through a mechanism similar to that of the rAAV genome.
INTRODUCTION
The adeno-associated virus (AAV) is a small, nonenveloped, and icosahedral particle with a diameter of 25 nm. Recombinant AAV (rAAV) has been widely used in gene therapy as vectors to transfer genes into target cells or tissues due to several advantages. 1,2 rAAV can transfer a gene of interest into dividing and nondividing cells. Different serotypes of AAV have specific tropisms and a low probability of causing an inflammatory response. The most widely used platform for producing rAAVs involves transfecting HEK293 cells with either two or three plasmids: in case of three plasmids, one encoding the gene of interest, one containing the AAV rep/cap genes, and another carrying helper genes. 3 –5 However, a notable issue is the presence of nonvector genome DNA within the capsid. 5,6 Two sources of DNA impurities arise from the host cell DNA and plasmids that support vector production (e.g., rep/cap genes, helper genes, or their background of the plasmids). Host cell DNA encapsulated in capsids has been quantified using quantitative PCR (qPCR) 7 –9 or digital droplet PCR (ddPCR). 10 Packaged residual DNA impurities derived from vector templates and helper sequences were reported to range from 1% to 8% of the vector genome DNA in purified vector particles. 11 –14 The presence of transforming factors such as the E1 gene in HEK293 derivatives as cell substrates raises concerns over the existence and amount of these potentially deleterious DNA sequences in preparations of viral vectors. 15 In addition, these DNA impurities, including those of the plasmid or virus origin, have the potential to unintentionally activate the immune system. 5 A report has suggested that after administration of rAAV in vivo, plasmid DNA is detected within target tissues or cells, occasionally resulting in the detection of transcripts and translation products. 15 Hence, rAAVs used for gene therapy should aim to minimize the potential of causing unintended immune responses. Moreover, the quantity of DNA impurities contained in manufactured rAAVs should be minimized as much as possible. Because encapsulated DNA impurities are resistant to DNase treatment and are still present in the final stage of the products, several attempts have been made to reduce the amount of DNA impurities within the capsid. 11,16 –19 In this study, we analyzed packaged DNA impurities in rAAV samples that were highly purified using ultracentrifugation. The samples were subjected to ddPCR, and DNA strand form assays were performed; the results indicated that DNA impurities were packaged into the capsid in a single-stranded form.
MATERIALS AND METHODS
Plasmids used for rAAVs production and preparation of rAAVs
For the production of rAAV, three plasmids were used as follows: pAAV-ZsGreen1 (SYN6339; Takara Bio, Inc., Kusatsu, Japan), pHelper (SYN6340; Takara Bio, Inc.), and a rep-cap plasmid, which varied depending on the rAAV serotype (pRC1 [SYN63341-2; Takara Bio, Inc.] for rAAV1, pRC2-mi342 [SYN6341-1; Takara Bio, Inc.] for rAAV2, pRC5 [A101-1; Takara Bio, Inc.] for rAAV5, or pRC6 [SYN6285; Takara Bio, Inc.] for rAAV6). 293T cells were obtained from the American Type Culture Collection (CRL-3216) and cultured in Dulbecco’s modified Eagle’s medium (D5796; Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (S1760; Biowest, Nuaille, France). Purified rAAV1 (1908_rAAV1-CMV-ZsGreen1), rAAV2 (1909_rAAV2-CMV-ZsGreen1), rAAV5 (1910_rAAV5-CMV-ZsGreen1), and rAAV6 (1911_rAAV6-CMV-ZsGreen1) were provided by the Manufacturing Technology Association of Biologics. To generate the rAAVs, 293T cells at 70–90% confluence were transfected using the PEIpro reagent (no. 115; Polyplus Transfection, Illkirch, France) in a HYPERFlask cell culture vessel (no. 10034; Corning, NY). The plasmid mixture used for transfection contained pAAV-CMV-ZsGreen1, pHelper, and rep-cap plasmids in a 1:1:1 weight ratio. The PEIpro reagent and DNA were mixed at a 1:1 weight ratio in serum-free Dulbecco’s modified Eagle’s medium. After 72 h, the cells were lysed using a buffer containing 0.5% Triton X-100 (no. 1.08643.1000; Merck, Rahway, NJ) and 2 mM MgCl2 in phosphate-buffered saline (PBS; 10010-049; Thermo Fisher Scientific, Waltham, MA). The lysate was treated with 90 U/mL benzonase (1.01697.0001; Merck, Darmstadt, Germany) at 37°C for 1 h to degrade residual nucleic acids. The crude lysate was concentrated to 1/8th of its original volume using tangential flow filtration (N02-E100-05-N; Spectrum, Stamford, CT) and loaded onto a HiTrap AVB Sepharose column (28-4112-12; GE Healthcare) for purification. After washing the column, rAAV was eluted using 50 mM glycine-HCl (pH 2.7). Full particle condensation was achieved using CsCl density gradient centrifugation at 148,500 g for 46 h at 21°C. The rAAV particles were finally dialyzed against PBS and stored at −80°C.
Titration of rAAV and quantitation of DNA impurities by ddPCR
For ddPCR, rAAV samples were treated with or without 1 unit of DNase I (2270A; Takara Bio Inc.) at 37°C for 30 min, and the encapsulated rAAV genome and DNA impurities were extracted using heat treatment at 95°C for 10 min. 20 All primers and probes were custom-synthesized and purified using high-performance liquid chromatography (Eurofins Genomics K.K., Tokyo, Japan). For titration of rAAV, forward primer 5′-GGAACCCCTAGTGATGGAGTT-3′, reverse primer 5′-CGGCCTCAGTGAGCGA-3′, and probe 5′-[FAM]CACTCCCTCTCTGCGCGCTCG[MGB]-3′ were used. 10 For quantitation of the amp R gene, forward primer 5′-TTGATCGTTGGGAACCGGAG-3′, reverse primer 5′-TTGTTGCCGGGAAGCTAGAG-3′, and probe 5′-[FAM]ACGACGAGCGTGACACCACGA[BHQ1]-3′ were used. For quantitation of pAAV-ZsGreen1 backbone unique, forward primer 5′-CTTACAGACAAGCTGTGACCG-3′, reverse primer 5′-CGTATCACGAGGCCCTTTCG-3′, and probe 5′-[FAM]CACCGTCATCACCGAAACGCGC[BHQ1]-3′ were used. For quantitation of AAV2 rep gene in pRC1, pRC2-mi342, pRC5, or pRC6, forward primer 5′-CGGAGAAGCAGTGGATCCA-3′, reverse primer 5′-ATTTGGGACCGCGAGTTG-3′, and probe 5′-[FAM]CACCGTCATCACCGAAACGCGC[BHQ1]-3′ were used. For quantitation of adenovirus type 5 E4 gene, the forward primer 5′-CGTCCGGCGTTCCATTTGGC-3′, reverse primer 5′-ACATTCGGGCAGCAGCGGAT-3′, and probe 5′-[FAM]TTGAGACAGAGACCCGCGCTACC[BHQ1]-3′ were used. The reaction mixture for ddPCR was composed of 10 μL ddPCR supermix for probes (Bio-Rad Laboratories, Hercules, CA, USA), 2 μL of primers (final concentration of each primer was 0.9 μM), 1 μL of probe (final concentration, 0.25 μM), and 1 μL of the template diluted with TE (10 mM Tris, 1 mM EDTA) in a final volume of 20 μL. The plates were transferred to a QX200 Automated Droplet Generator (Bio-Rad). A 96-well plate containing the generated droplets was transferred to a C1000 Touch Thermal Cycler (Bio-Rad). The cycling conditions were as follows: 10 min at 95°C, followed by 40 cycles of a two-step thermal profile comprising 30 s at 95°C and 60 s at 60°C. The plate was then transferred to a QX200 droplet reader (Bio-Rad), and data analysis was performed using QuantaSoft software (version 1.7.4.0917; Bio-Rad). The threshold separating the negative and positive droplets was manually set just above or below the cluster of negative and positive droplets, respectively.
DNA strand form assay for DNA impurities
After treatment of the rAAV with 5 units of DNase I (Takara Bio Inc.) for 30 min at 37°C, the encapsulated DNA was extracted using Quick-DNA/RNA Viral MagBead (R2140; Zymo Research, Orange, California, USA). For the DNA strand form assay, the extracted DNA was treated with 10 units of DNase I (2270A, Takara Bio Inc.), 5 units of Lambda exonuclease (M0262, New England Biolab Inc.), or 100 units of Nuclease P1 (M0660, New England Biolab, Inc.). The DNase activity was terminated by adding equal volume of 20 mM EDTA (ethylenediaminetetraacetic acid) and heating at 75°C for 15 min. Copy number of DNA impurities was determined by qPCR using the primers listed in Supplementary Table S1. For qPCR of host cell DNA, forward and reverse primers for 18S rRNA were used (Supplementary Table S1). 10,21 Each 20 μL qPCR prepared with TB Green Premix Ex Taq II (Tli RNaseH Plus) (RR820; Takara Bio Inc.) contained a final concentration of 0.4 μM of each primer. qPCR was performed on a StepOnePlus system (Applied Biosystems, Waltham, MA, USA) under the following thermal cycling conditions: 20 s at 95°C, followed by 40 cycles of a two-step thermal profile comprising 3 s at 95°C and 30 s at 60°C. Each DNA template was tested in triplicate.
RESULTS
Detection of DNA impurities contained in rAAV samples by ddPCR
To investigate DNA impurities in purified rAAV1, rAAV2, rAAV5, and rAAV6 samples, ddPCR was performed using common and unique primers and probes for the plasmids. The rAAVs were produced by transfection using three plasmids (weight ratio, 1:1:1), namely, pAAV-ZsGreen1, rep-cap plasmid pRC (pRC1, pRC2-mi342, pRC5, or pRC6), and pHelper (Fig. 1). The total copy number of the three plasmids was measured using the ampicillin resistance gene (amp R) and independently measured using the pAAV backbone unique sequence for pAAV-ZsGreen1, rep gene for pRC, and E4 gene for pHelper. Following treatment of rAAV with DNase I, there was no decrease in rAAV1-ZsGreen1 and rAAV6-ZsGreen1 titers, whereas the titers of rAAV2-ZsGreen1 and rAAV5-ZsGreen1 decreased to 82.3% and 90.9%, respectively (Supplementary Fig. S1). Following DNase I treatment, 1.45E + 11 copies of amp R in the 1.76E + 13 vg of rAAV1-ZsGreen1 sample were detected (Table 1). DNase I treatment did not cause a distinct decrease in the copy numbers of amp R, rep, and E4, either (Supplementary Fig. S2). After DNase I treatment, the copy numbers of amp R responding to the sum of the three plasmids were almost equal to the number of pAAV backbone unique sequences (98.88%), and the copy numbers of the rep and E4 genes were 0.91% and 0.21%, respectively (Tables 1 and 2). rAAV2-ZsGreen1, rAAV5-ZsGreen1, and rAAV6-ZsGreen1 contained 0.69–3.27% DNA impurities derived from the plasmids (Table 1), and 99.17–99.51% of them were from pAAV-ZsGreen1 (Table 2). These results indicated that pAAV-ZsGreen1 showed high preferential encapsulation into an rAAV capsid, although the copy number of pAAV-ZsGreen1 was measured with the 247-base backbone unique sequence discriminated from pRC and pHelper.
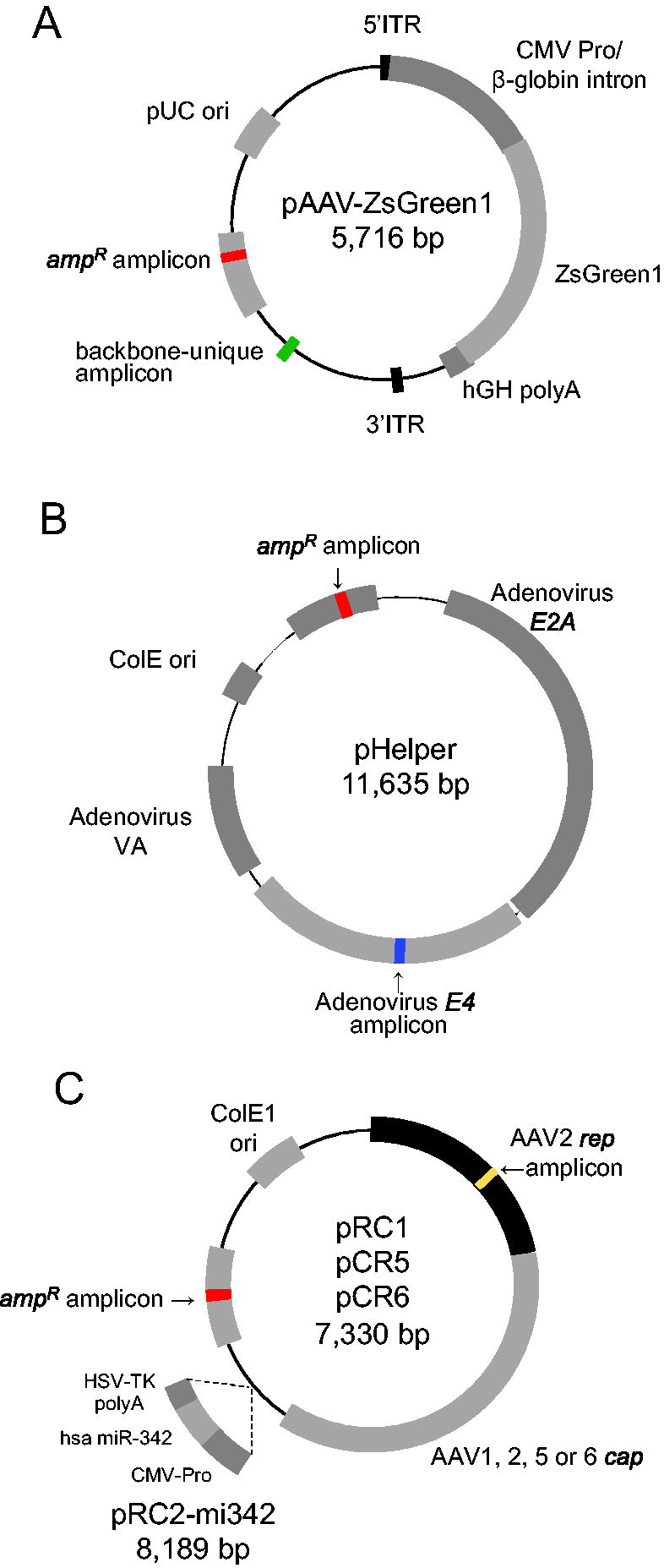
Structure of plasmids used for rAAV production.
rAAV Titers and Copy Number of ampRa Determined by Digital Droplet PCR
amp R is a common amplicon among the three plasmids.
rep is the amplicon for pRC, and E4 is the amplicon for pHelper.
Encapsulation of single-stranded form of DNA impurities derived from plasmid DNA into the capsids
The rAAV genome is encapsulated in the capsid either as a single-stranded positive or negative form. 1,2 During this process, Rep-binding elements in the ITR (Inverted Terminal Repeat) play a crucial role in packaging of the genome. The rAAV genome is packaged into capsids through a pore at a vertex formed by five triangular surfaces of Rep proteins. 22 Rep68/78 binds to the Rep-binding element localized in the ITR, and the complex is recruited to access the pore on the surface of the capsid. Rep52/40 packages single-stranded DNA (ssDNA) into the capsid. In this study, to determine whether DNA impurities are similarly encapsulated in a single-stranded form, we used three types of DNases and established a DNA strand form assay (Fig. 2A, B). 23 DNase I is an endodeoxyribonuclease that catalyzes degradation of both ss- and double-stranded DNA (dsDNA) randomly to the same extent. Lambda exonuclease displays highly progressive degradation of dsDNA in the 5′ to 3′ direction but not of ssDNA, 24 whereas nuclease P1 is an ssDNA-specific endonuclease with no base specificity. 25 These DNases were used to determine the DNA form of DNA impurities encapsulated in the capsids. In the DNA strand form assay, we used dsDNA from the rAAV-ZsGreen1 fragment cleaved from pAAV-ZsGreen1 by PvuII digestion as a control substrate (Fig. 2C). The dsDNA showed susceptibility for digestion with DNase I or lambda exonuclease, whereas rAAV-ZsGreen1 ssDNA prepared by heat treatment displayed susceptibility to nuclease P1 but not to lambda exonuclease as shown in our previous report, indicating that rapid formation of T-shaped hairpin of 5′-ITR and 3′-ITR after denaturation could prevent rehybridization of complementary ssDNA. 23 DNA extracted from rAAV6-ZsGreen1 capsids exhibited DNase sensitivity similar to that of ssDNA (Fig. 2C, D). We next applied the DNA strand form assay to DNA impurities obtained from rAAV1-ZsGreen1, rAAV2-ZsGreen1, rAAV5-ZsGreen1, and rAAV6-ZsGreen1 (Fig. 3). DNA impurities were prepared from the rAAV capsids digested with each DNase for 30 min. Subsequently, undigested DNA impurities were detected by qPCR using primers specific for the three plasmids (Supplementary Table S1). DNA impurities derived from pAAV1-ZsGreen1, pRC1, and pHelper plasmids encapsulated in the capsids were completely digested with DNase I or nuclease P1, but not with lambda exonuclease (Fig. 3A). Other AAV samples tested also showed similar results (Fig. 3B–D). These results suggest that the DNA impurities were translocated into the capsid in a single-stranded form, similar to the rAAV genome packaged by Rep proteins. It should be noted that 38.1–41.3% of the DNA impurities from rAAV1-ZsGreen1 was digested by lambda exonuclease. Other DNA impurities also displayed a similar reduction by lambda exonuclease compared with each control, indicating that part of the ssDNA could successfully rehybridize with the complementary strand to form dsDNA, which was susceptible to digestion by lambda exonuclease. Furthermore, it should be noted that nuclease P1 treatment could destroy the PCR templates of the partially hybridized intramolecular or intermolecular hybridized dsDNA through endonuclease activity.

DNA strand form assay.

DNA strand form assay on DNA impurities derived from the plasmids.
DNA strand form assay to assess host cell DNA encapsulated in the capsids
Similarly, host cell DNA was analyzed using the DNA strand form assay (Fig. 4). Host cell DNA was detected as described previously. 10,21 As observed with DNA impurities derived from plasmids, host cell DNA was susceptible to DNase I and nuclease P1, but resistant to lambda exonuclease. These results also indicate that the host cell DNA encapsulated in the capsids was in a single-stranded form. Notably, the copy number of the 18S rRNA gene digested with lambda exonuclease was comparable to that of the control in rAAV2-ZsGreen1, rAAV5-ZsGreen1, and rAAV6-ZsGreen, indicating that the 18S rRNA gene was less likely to form a partial double-stranded form at the amplicon compared with plasmid DNA. These results indicated that the DNA impurities were encapsulated in ssDNA form, possibly using the same machinery as that of the rAAV genome mediated by Rep proteins.
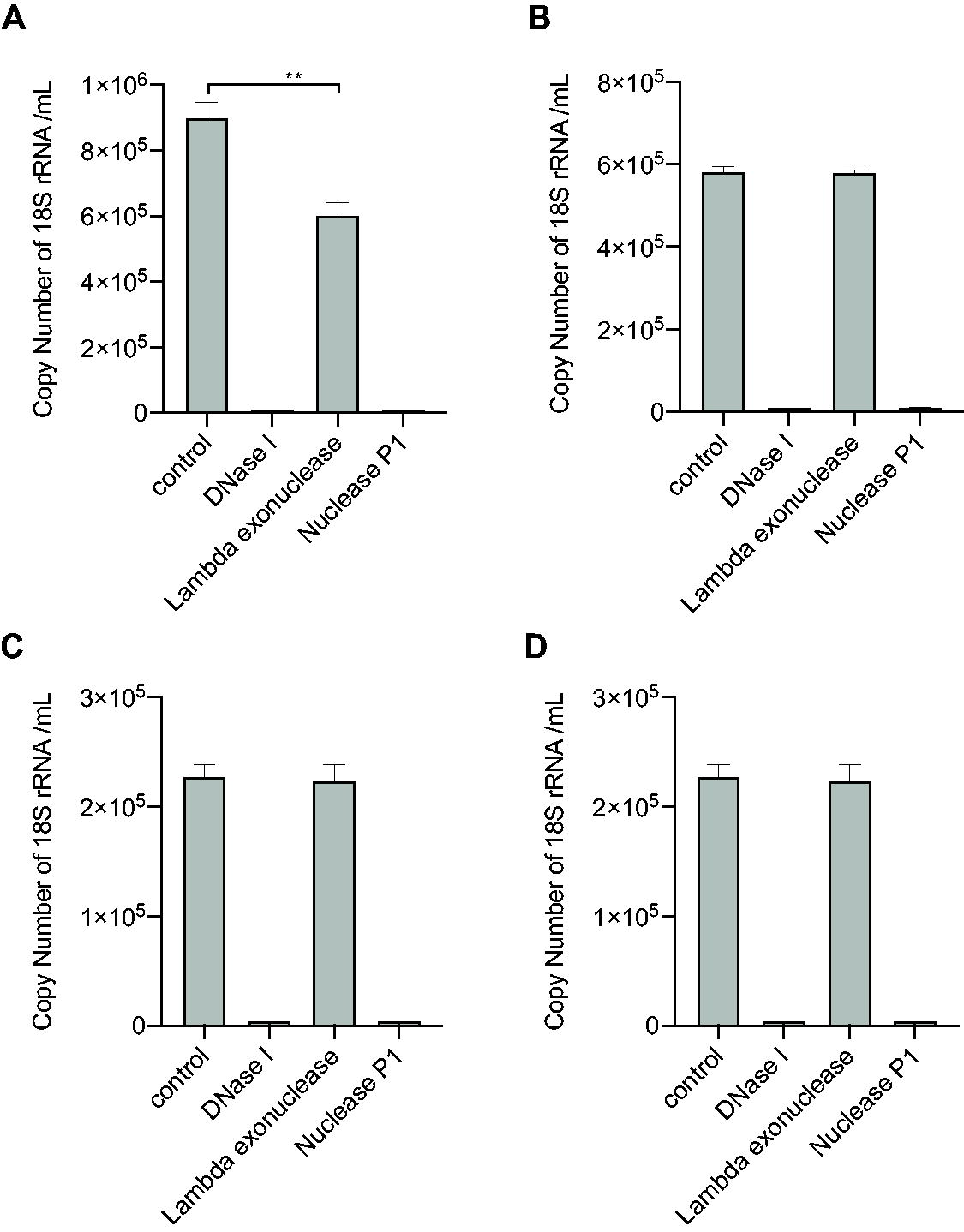
DNA strand form assay on DNA impurities derived from host cell DNA.
DISCUSSION
In this study, highly purified rAAVs using three plasmids contained plasmid DNA impurities ranging from 0.69% to 3.27% by ddPCR (Table 1). We independently assessed the three plasmids using plasmid-specific primer pairs, and the main source of encapsulated plasmid DNA was identified as the pAAV-ZsGreen1 backbone (98.88–99.51%), whereas the copy numbers of DNA impurities from pRC and pHelper plasmids ranged from only 0.39% to 0.91% and 0.10% to 0.21%, respectively (Table 2). DNA impurities, including host cell DNA, were encapsulated into the capsids as a single-stranded form, as shown by a DNA strand form assay (Figs. 2–4). These results indicated that the DNA impurities were packaged into the capsid by Rep proteins, as well as the rAAV genome, as suggested by previous reports. 5 Rep proteins play crucial roles in vector DNA replication and its packaging into capsids. 26,27 In this study, AAV2 Rep was provided by cotransfected plasmids pRC1, pRC2-mi342, pRC5, or pRC6. Four Rep polypeptides (Rep78, Rep68, Rep52, and Rep40) are located in the same (ORF) Open Reading Frame and translated from transcripts generated by the p5 and p19 promoters. 28 Large Rep proteins (Rep78/68) are essential for AAV genome replication and bind to a 16-bp cis element (Rep-binding element) present in ITRs. 29 The small Rep protein Rep52/40 is crucial for packaging rAAV genome into capsids composed of VP1, VP2, and VP3, and rAAV genome was encapsulated by capsid-bound helicase activity through a pore located on the empty capsid surface by orientation from 3′ to 5′ termini. 27 Similarly, the pAAV-ZsGreen1 backbone DNA with ITR(s) was expected to be translocated into capsids in a single-stranded form through Rep-mediated machinery. pAAV-ZsGreen1-derived impurities occupied more than 98.88% of DNA impurities in the rAAV samples because pAAV-ZsGreen1 was cleaved out as a linear DNA fragment at nicking sites present at 15 bp flanking the Rep-binding elements by activity of Rep, and the backbone DNA fragment containing Rep-binding elements was efficiently translocated to the capsids. However, pHelper has no nicking site and retains a circular form, which is not an appropriate substrate for the Rep encapsulation machinery. Although pRC has a Rep binding element in P5, and the nicked pRC gets converted into an open circular form, a nonlinear form of the plasmid is not appropriate for encapsulation. Furthermore, the intact sizes of 11.6 kb of pHelper and 7.3–8.2 kb of pRC constructs are too long to be packaged into the capsids. The rAAV samples used in this study were purified using CsCl gradient centrifugation. The sizes of the 2.6 kb rAAV genome and 3.1 kb pAAV-ZsGreen1 backbone were close to the present in similar size range fractionated by CsCl density, whereas the capsids containing shorter or longer DNA impurities might be removed through the purification process.
ITRs at both ends were able to stabilize ssDNA shown by DNA strand form assay with the different types of DNases as described in our previous report. 20 In this study, the same assay, that is, the DNA strand form assay, was used to determine the form of DNA impurities encapsulated in the capsids (Fig. 2). Lambda exonuclease is a nonspecific endonuclease degrading dsDNA from 5′ to 3′ direction, but has negligible activity for degradation of ssDNA, whereas nuclease P1 is a nonspecific endonuclease that degrades ssDNA but not dsDNA. If the DNA impurities are in the ssDNA form, it is expected that the ssDNA extracted from capsids may take a partial intramolecular double-stranded form or the part of ssDNA may rehybridize with complementary DNA strand to form dsDNA. In our study, several amplicons failed to show susceptibility to degradation using nuclease P1 and resistance to lambda exonuclease activity (data not shown). In the assay, the setting of amplicons on the target templates was critical for obtaining a definite result that DNA impurities were present in an ssDNA form encapsulated in the capsids, as well as rAAV genome.
According to the guidelines, 30,31 DNA impurities in gene therapy administration doses should be maintained at levels of less than 10 ng and a size less than 200 bp. DNA impurities external to the capsids were expected to be degraded by DNase treatment, whereas DNase-resistant DNA impurities were localized within the capsids. 5,6 This implies that after administration of rAAV DNA into a recipient, DNA impurities containing ss- and dsDNA are also introduced into cells or tissues; however, it is notably emphasized that there is a need to convert ssDNA into a double-stranded form for the expression of transcripts and proteins in vivo. This suggests that adverse effects of the single-stranded form of DNA impurities may differ from those of dsDNA. It may be necessary to reconsider or evaluate the effects of single-strand form of DNA impurities independently.
Footnotes
AUTHORS’ CONTRIBUTIONS
Y.Y., K.H., E.I.-K., and K.M.-K.: Designed the study. Y.Y., K.Y., and K.U.: Wrote the article. All the authors have read and approved the final article.
GRAPHS AND STATISTICAL ANALYSIS
Figures were prepared using PowerPoint. Graphs and statistics were generated using GraphPad Prism 8.0.
DATA SHARING STATEMENT
Original data, resources, and protocols are available upon request by email:
AUTHOR DISCLOSURE STATEMENT
The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the article, apart from those disclosed.
FUNDING INFORMATION
This work was funded by AMED under the grant numbers
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.