
Abstract
Monoclonal antibodies (MAbs) against prion protein (PrP) are powerful tools for diagnosis and research in transmissible spongiform encephalopathies. Ten MAbs to recombinant/native cellular PrP (PrPc) in mammals were prepared with a simple method and identified in detail. Normal BALB/c mice were immunized with the recombinant bovine mature PrP (rbomPrP) and PrP27-30 (rboPrP27-30) expressed in Escherichia coli. The immunized splenocytes were fused with SP2/0 mouse myeloma cells, and positive hybridomas were selected by indirect enzyme-linked immunosorbent assay (ELISA). The characterizations of these MAbs, such as Ig, Ig subclass, titer, affinity index, specificity, epitopes recognized, and binding to recombinant/native PrPc of cattle, sheep, or human beings, were evaluated by Western blotting and indirect or sandwich ELISA. Ten MAbs could be divided into five groups depending on the results of indirect ELISA additivity test and their reaction to E. coli-expressed truncated-PrPs. Isotyping of the MAbs revealed that they belong to IgG1, IgG2a, and IgG2b subclass. Their indirect ELISA titers were between 103 and 106. Affinity constants were between 109 and 1012 M−1. Ten MAbs specifically reacted with the rbomPrP, without binding to prion-like protein Doppel and the lysates of E. coli. These MAbs could also respond to the recombinant mature PrP (rmPrP) of sheep and human beings. Also of interest was the ability of the MAbs to bind with dimer of rmPrP and PrP extracted from the brain tissue of cattle or sheep. We conclude that anti-PrP MAbs successfully prepared with a simple method could potentially be useful in mammalian prion research.
Introduction
TSEs do not elicit any immune response to the inoculated infectious particles due to the immune tolerance, and no scrapie-specific antibodies or interferons are found.(4,5) However, with the discovery of PrP27-30 in fractions highly enriched for scrapie infectivity and the successful purification of a large-scale of prion rods, antisera were obtained in rabbits.(6) Subsequently, MAbs were produced in mice.(7) At the time, preparation of antisera or MAbs required a large amount of purified PrP27-30 as the immunogen. The amount of PrP27-30 was greatly reduced when the knockout mice (Prnp0/0) with disrupted PrP genes were immunized.(8) The purified PrP27-30 as the immunogen to generate antibodies in Prnp0/0 mice is now often substituted by synthetic peptide or the recombinant protein, and more anti-PrP MAbs are easily produced.(9) Although there is a lot of anti-PrP MAbs, only a few of them are generated in normal BALB/C mice, with recombinant PrP expressed in Escherichia coli, the most familiar expression system. Ishiguro and his co-workers have successfully prepared the MAbs against chicken PrPc by immunizing BALB/c mice with recombinant PrP.(10) To our knowledge, it is the simplest way for preparation of anti-PrP MAbs using normal BALB/c mice and the recombinant PrP expressed in E. coli, which avoids the limitation of Prnp0/0 mice and purified PrP27-30. By this method, we obtained 10 anti-PrP MAbs, which generate good reaction to the recombinant PrP or PrPc from brain tissue of mammals.
Materials and Methods
Production of MAbs
Cloning of gene coding prion protein fragment
Genome DNA was extracted from the vein blood of healthy Chinese yellow cattle. The open reading frame (ORF) of the bovine prion protein was amplified with the genome DNA as the template using primers 1 and 11. The amplified ORF was cloned into the vector pMD-18T (Takara Bio, Dalian, China) and the recombinant plasmid boPrPORF-18T was generated. The gene coding mature prion protein (mPrP) was amplified with boPrPORF-18T as the template using primers 2 and 22 and then cloned into the expression vector pET-30a(+) (Novagen, Madison, WI) digested with enzymes EcoRI and XhoI, and the recombinant expression plasmid bomPrP-30a was produced. The gene coding bovine PrP27-30 fragment was amplified with primers 3 and 33 (Table 1) and inserted into enzyme sites NdeI and XhoI of pET-30a(+), and the expression plasmid boPrP27-30-30a was formed.
Antigen preparation
The plasmid bomPrP-30a and boPrP27-30-30a were transformed into E. coli JM109(DE3) (Promega, Shanghai, China), respectively. The corresponding recombinant E. coli were cultured and induced by 1 mM isopropylthiogalactoside (IPTG) (Calbiochem, Beeston, United Kingdom). The induced E. coli was harvested by centrifugation and then subjected to sonication. Then, antigens were purified by affinity chromatography with Ni-NTA resin. The purified recombinant antigens, namely, rbomPrP and rboPrP27-30, were concentrated by centrifugation after dialysis and suspended in phosphate-buffered saline (PBS). Protein concentration was determined by absorbance at 280 nm and at 260 nm according to the equation.
Immunization and cell fusion
Murine MAbs were produced against purified rbomPrP and rboPrP27-30 using the modified method of Kohler and Milstein.(11) Splenocytes were collected from female BALB/c mice immunized intraperitoneally (ip) with four biweekly doses of 50 − 80 μg purified proteins. Splenocytes were fused with SP2/0 myeloma cells at a ratio of 5:1 in the presence of 50% polyethylene glycol 1500 (Sigma, Beijing, China). The fused cells were suspended in hypoxanthine-aminopterin-thymidine (HAT) RPMI 1640 medium (Gibco BRL, Grand Island, NY) and incubated at 37°C in 5% CO2 incubator.
Hybridoma screening and cloning
Hybridoma supernatants for antibody production were primarily screened with indirect enzyme-linked immunosorbent assay (ELISA) against purified antigen of rbomPrP and rboPrP27-30. Specific antibody-producing hybrid clones were recloned four times by limiting dilution and specific hybrid clones were expanded.
Purification of the MAb ascites
Positive clones were injected into pristane-primed mice and finally ascitic fluid was collected. The ascitic fluid was precipitated with 55% saturated ammonium sulfate (Amresco, Solon, OH) and further purified with protein A affinity resin (Calbiochem). The purified MAb was analyzed by sodium dodecylsulfate polyacrylamide gel electrophoresis (SDS-PAGE) to detect the heavy and light immunoglobulin chains.
Characterization of MAbs
Isotyping of MAbs
Isotyping of MAbs was performed by capture ELISA using mouse monoclonal antibody isotyping kit (Sigma) following the manufacturer's recommendations.
Measurement of MAb titer by ELISA
The titer of MAbs was measured by a classical indirect ELISA. Briefly, the rbomPrP or rboPrP27-30 (10 μg/mL in sodium carbonate buffer) was bound to the polystyrene microtiter plate. Bovine serum albumin (BSA) and skim milk powder diluted at 5% were added to block the blank of the wells. The purified MAb ascites with serial dilutions was added to the well. Horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG was then added. The substrate solution containing O-phenylenediamine and hydrogen peroxide (OPD/H2O2) was added. The reaction was stopped by the addition of 1 M H2SO4 and the absorbance at 492 nm for the stopped product was measured. Triplicates of each of the MAb dilutions were run. Absorbance versus MAb dilution was plotted using the mean value. The dilution fold was the MAb titer when the absorbance reached 0.8.
Measurement of MAb affinity constant
Indirect ELISA, as described above, was applied to make the antibody dilution curve. The rbomPrP with lower concentration (1 μg/mL) worked as the antigen, and MAb in series of dilution was added. Relative affinity may be estimated from the MAb concentration (mol/L), at which ∼50% of plateau binding occurs. The affinity constant was the reciprocal of the MAb concentration at half saturation.(12)
Identification of MAb specificity
Specificity of MAbs was identified by indirect ELISA through their reaction with E. coli JM109(DE3) containing pET30a(+) vector and the prion-like protein Doppel. The fusion mature Dpl of cattle, sheep, and human beings was generated in E. coli JM109(DE3) containing the corresponding recombinant plasmid with pET30a(+) as the expression vector, as described above (primers not shown). In addition, Dpl extracted from the testes of cattle and sheep was also applied to test the specificity of the MAbs.
Identification of recognized epitopes of MAb
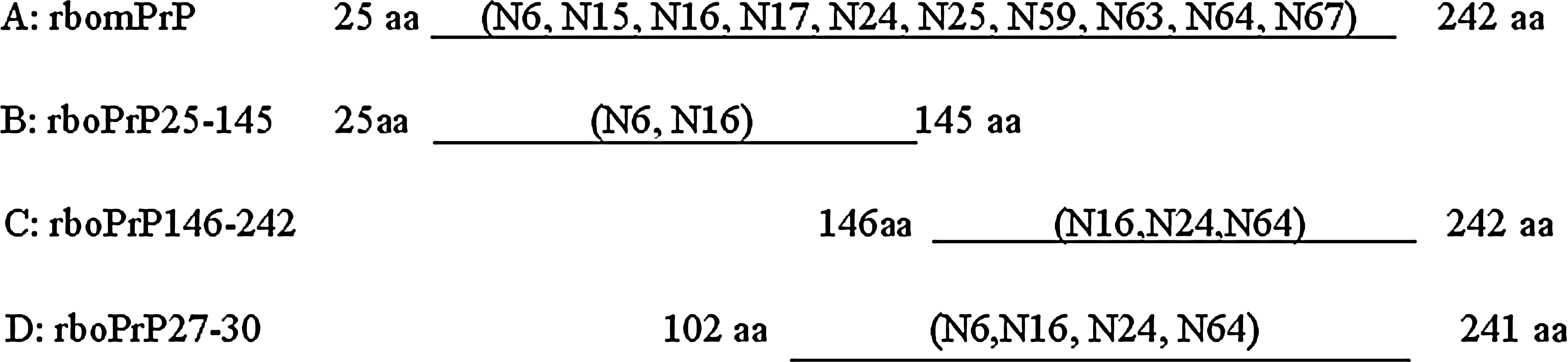
The epitopes recognized by MAb were identified depending on indirect ELISA additivity test and the reactive capability with the E. coli-expressed deletion fragments of PrP detected by indirect ELISA. The additive ELISA was based on the method of Friguet et al.(13) The coating was carried out with 50 μL per well of purified rbomPrP. The additivity index (AI) was calculated according to the formula, AI = {(2A1+2/(A1 + A2)) − 1} × 100, where A1, A2, and A1+2 are the absorptions reached with the first MAb alone, the second MAb alone, and the two MAbs together, respectively. Giving AI ≥ 50%, the two MAbs would be regarded to bind different epitopes. For the E. coli-expressed deletion experiment, genes encoding the fragments comprising amino acids 25 to 145 and 146 to 242 of bovine PrP were inserted into enzyme sites EcoRI and XhoI of pET30a(+) vector. The fusion proteins rboPrP25-145 and rboPrP146-242 were produced by the E. coli BL21(DE3) plys containing the corresponding plasmid induced by 1 mM IPTG. The purified fusion proteins, including rbomPrP, rboPrP25-145, rboPrP27-30, and rboPrP146-242 were used to coat the microtiter plates, respectively. The other procedures of the indirect ELISA were the same as above.
Efficacy of developed MAbs
To value the developed MAbs, Western blotting and sandwich ELISA were carried out to detect the fusion mPrP or the prion protein existing in the healthy cattle and sheep using the prepared MAbs.
Sample preparation
The fusion mPrP of sheep or human beings was prepared as described above. For the preparation of prion from brain, the obex tissue should be removed as soon as possible after death. The samples were homogenized in ice-cold PBS containing 0.5% Nonidet P-40 and 0.5% deoxycholate to give a 10% (w/v) final suspension. Homogenates were centrifuged for 10 min at 5000 g at 4°C. Supernatants were aliquoted and stored at −70°C for detection.
Western blot analysis
When Western blotting was carried out, native or PK-digested (final PK concentration of 5 μg/mL for 30 min at 37°C, heat inactivated) samples were electrophoresed on 12% gel. The separated proteins were transferred to the nitrocellulose membrane. The sheet was incubated in 5% BSA and skim milk powder solution in PBS to block nonspecific binding. The membrane was then incubated in the solution of MAb, followed by incubation with goat anti-mouse IgG-HRP. After thorough washing, the sheet was incubated with chromogenic substrate diamino benzidine (DAB, Sigma) and the specific bands were noted.
Sandwich ELISA
Microtiter plates were coated with 5 μg/mL MAb in 50 mM carbonate/bicarbonate buffer (pH 9.6). The plates were blocked as above, and 50 μL samples were applied to the wells. After 1 h of incubation at 37°C, another developed MAb recognizing different epitope, conjugated with HRP, was added to the wells. After 1 h incubation at 37°C, reaction was detected as an indirect ELISA.
Results
Expression and purification of fusion protein his-tagged PrP
Nine his-tagged fusion proteins, including rbomPrP, rboPrP25-145, rboPrP146-242, rboPrP27-30, rovmPrP (mature PrP of sheep), rhomPrP (mature PrP of human beings), rbomDpl (mature Dpl of cattle), rovmDpl (mature Dpl of sheep), and rhomDPl (mature Dpl of human beings), were expressed in E. coli JM109(DE3) transformed with the corresponding recombinant plasmids using pET30a(+) as the expression vector upon the induction of 1.0 mM IPTG. Fusion proteins were mainly expressed in the form of inclusion bodies. Inclusion bodies were solubilized with 8 M urea, 0.1 M NaH2PO4, and 0.1 M Tris-HCl (pH 8.0). The supernatant was purified by Ni–NTA column and refolded by dialysis with a gradient of decreasing concentration of urea. The product achieved up to 90% purity, as determined by SDS-PAGE (results not shown).
Establishment of hybridoma cell lines
Two cell fusions were performed with spleens from mice immunized with rbomPrP and rboPrP27-30. The hybridoma supernatants were screened by indirect ELISA for their capacity to bind rbomPrP or rboPrP27-30. Lysates of E. coli JM109(DE3) harboring pET30a(+) empty vector were used to eliminate antibody against proteins produced by the expression vector and host strain. Nine hybridoma cell lines (clones N15, N17, N25, N27, N59, N63, N64, N67, and N79) secreting specific antibody against rbomPrP and three cell lines (clones N6, N16, and N24) against rboPrP27-30 were isolated and subcloned by limiting dilution. Ten clones except for N27 and N79 could stably produce MAb after being cultured for 3 months in vitro. Therefore, 10 clones (N6, N15, N16, N17, N24, N25, N59, N63, N64, and N67) were selected for ascites production. MAbs were precipitated with 55% saturated ammonim sulfate followed by purification in protein A affinity columns from ascites. Purification of ascites was 95% homogeneity as assessed by SDS-PAGE (Fig. 1).

SDS-PAGE analysis of MAb N64 from ascitic fluids purified by Protein A agarose. Lane M, protein molecular weight marker; lanes 1 and 3, purified MAb; lane 2, unpurified MAb.
Characterization of MAb
Table 2 summarizes the characterization of 10 clones selected in terms of titer, affinity, class, and subclass, indicating that the MAbs are G1, G2a, and G2b subclass. They are high-affinity type MAbs ranging from 109 to 1012 M−1.
Antibodies N6, N15, N16, N17, N24, N25, N59, N63, N64, and N67 could specifically react with rbomPrP revealed by indirect ELISA. They did not bind lysates of E. coli harboring pET30a(+) vector, the recombinant Dpl and Dpl from the testes tissue.
The results of additive ELISA showed that 10 MAbs could recognize five different epitopes. On the basis of the comparison between AI (Table 3), 10 MAbs can be divided broadly into five groups—group 1: antigen epitopes recognized by N15, N25, N59, N63, and N67 could be close to each other; group 2: N24 and N64 could recognize the same or close epitope; group 3: N6 recognized the different epitope; group 4: N16 was against another kind of epitope; and group 5: N17 was against the distinguished epitope. The additive ELISA analysis was further confirmed by the reaction of MAbs with the expressed truncated fragment of PrP. Ten MAbs could react with rbomPrP. N24 and N64 could react with rboPrP27-30 and rboPrP146-242, indicating that the epitope was among 146 to 241aa. N6 could react with rboPrP25-145 and rboPrP27-30, indicating that N6 was against the epitope from 102 to 145aa. N16 had response to the three truncated fragments of mPrP, demonstrating that its epitope was around 145aa. There was no reaction when MAbs N15, N17, N25, N59, N63, and N67 were used to detect the three truncated fragments, indicating that they may recognize some conformation epitopes of mPrP (Fig. 2).
The reaction of 10 MAbs with expressed truncated bovine PrP.
Efficacy of developed MAbs
Indirect ELISA showed that 10 MAbs could bind the fusion mature PrP of cattle, sheep, and human beings. The ELISA results were further confirmed by Western blotting. The size of the protein bands was approximately 28 kDa and 56 kDa, which were in good accordance with that of the monomer and dimer of mPrP (Fig. 3).

Western blot analysis of MAb N64 reaction with rmPrP and PK-digested rmPrP of cattle, sheep, and human being. Lane 1, proteins of E. coli harboring pET30a(+) vector; lanes 2, 4, and 6, recombinant mPrP of cattle, sheep, and humans, respectively; lanes 3, 5, and 7, PK-digested recombinant mPrP of cattle, sheep, and humans, respectively.
Because the bovine PrP used in immunization was prokaryotic protein, its structure might be somewhat different from the PrP found in eukaryotic cells. Therefore, PrP extracted from the obex region of cattle or sheep was used to detect the binding ability of MAbs with PrP in eukaryotic cells. The brain tissue proteins were subjected to SDS-PAGE and Western blotting with the produced MAbs as the primary antibody. Western blotting results showed the MAbs had specific binding ability with native PrP and no response to PK-digested samples (Fig. 4). In addition, natural or PK-digested brain homogenates was detected by sandwich ELISA with MAb N6 working as the capture antibody coated on the microtiter plate and MAb N64 working as the detective antibody conjugated with HRP. The result of sandwich ELISA showed that PrP in the natural brain homogenate could be detected using MAbs N6 and N64.

Western blotting of the healthy brain tissue PrP and the PK-digested product detected by prepared MAb. Lanes 1, PK-digested bovine brain PrP; lane 2, bovine brain PrP; lane 3, bovine brain PrP; lane 4, PK-digested bovine brain PrP.
Taken together, these data showed that a strong specific response to recombinant mPrP or PrP27-30 or natural PrP in brain tissue could be generated by the MAbs produced through recombinant PrP immunization in normal BALB/C mice.
Discussion
TSEs are fatal neurodegenerative disorders that afflict both animals and humans. BSE, scrapie, and nvCJD are among the most notable TSEs. Propagation of TSE is believed to involve the conversion of PrPc into PrPsc. The mechanism of this conformational transition remains poorly understood and, despite intensive studies, the normal function of host-encoded PrPc is still enigmatic. Antibodies, especially MAbs, have been widely applied in the prion research and many new findings have been revealed through anti-PrP MAbs. The preparation of anti-PrP MAbs underwent a long period of research. Based on the differences in the immunized mice and the immunogen, the anti-PrP MAb preparation could be broadly divided into three stages. At first, the production of anti-PrP MAbs required large amounts of purified SAF extracted from brains of infected animals and used as an immunogen in BALB/c mice.(7,14) Then, transgenic mice Prnp0/0 with an ablated PrP gene were born and production of anti-PrP antibodies were facilitated.(8) Anti-PrP MAbs were prepared with less amounts of SAF in Prnp0/0 mice.(15) After this development, anti-PrP MAbs surged due to the use of synthesized peptides(9,16) or the recombinant PrP(10,17) working as the immunogen in BALB/C or Prnp0/0 mice. More and more researchers applied the latter method to produce anti-PrP MAb according to their own interests because it obviates the need for purified SAF immunogen or Prnp0/0 mice and greatly reduces the labor and difficulty during the preparation. Recombinant bovine mature PrP and PrP27-30 were prepared and used as the immunogen to generate MAbs in BALB/C mice in our study. Ten MAbs againt PrP were produced through two instances of cell fusion. Those MAbs provided us with very useful tools to research the PrP in vertebrate animals.
Dpl is a PrP-like protein, coded by a gene named PRND, located near the PRNP (prion protein coding gene) locus. Human Dpl is a 179aa protein showing approximately 25% sequence identity with the carboxyproximal two thirds of the human PrPc. The Dpl is highly expressed in the testes of adult mammals and, in contrast to PrP, only at very low levels in the adult CNS.(18) In this study, we were concerned with the similarity between PrP and Dpl, and when analyzing the specificity of the MAb against PrP, Dpl from cattle, sheep, and human beings, either the fusion protein or the Dpl from the testes except for man, were tested. There were no reactions between anti-PrP MAbs and Dpl. Because of their specificity, these MAbs may be usefully applied in studying mammalian PrP.
Ten anti-PrP MAbs could be divided into five groups according to the additive index, and the results were in line with epitope recognization by the produced MAbs on the basis of their reaction with the recombinant truncated segment of PrP. It is also noteworthy that, in the analysis of the epitope, we found that there was no reaction between the MAbs N15, N17, N25, N59, N63, and N67 and the three truncated fragments (25-145aa, 146-242aa, and 102-241aa). What was the epitope recognized by these six MAbs? The problem would be settled through more experiments.
The produced anti-PrP MAbs could detect the recombinant mPrP of sheep and human beings besides cattle. It was interesting that those MAbs could detect the dimer of recombinant mPrP, bands at molecular weights of PrP dimers, of the three mammals as determined by Western blotting. The results were the same when using SAF70 and SAF84 (Prion Protein Monoclonal Antibody, Cayman Chemical, Ann Arbor, MI) as the detecting antibody in the Western blotting (data not shown). Was a large amount of dimer formed during the recombinant mPrP preparation? Meyer and colleagues report that they confirm recombinant PrP to be a monomer by analytical ultracentrifugation and they explain that recombinant PrP could not form dimer due to the lack of glycosylation.(19) However, recombinant PrP could have been refolded into both α-helical and β-sheet structures as well as various intermediates in aqueous buffers. PrPc may possess a relatively open conformation which makes it susceptible to conversion into PrPsc under appropriate conditions.(20) The transition of recombinant PrP from a PrPc-like into a PrPsc-like conformation can be induced in vitro by some treatments. An α-helical dimer was the first step from the monomeric PrPc to highly aggregated PrPsc.(21) Although some treatments could induce recombinant PrP to form PrPsc-like, we could not determine what led to such a large amount of recombinant mPrP dimer formed in our study. The recombinant mPrP dimer and monomer could be completely digested with PK, and the digested products could not be detected with the produced MAb, indicating that those dimers and monomer are not PK-resistant PrP. In addition, the prepared MAbs can detect the nature PrP extracted from brain homogenate of obex region, and no reaction occurred after the obex tissue was treated with PK. In all TSEs, PrPsc accumulates mainly in the CNS and trace amounts of PrPsc exist in peripheral lymph nodes, blood, and urine.(22) PrPsc accumulates in peripheral lymph nodes well before involvement of the CNS. Immunological assays for PrP27-30 are now widely used in tests for the detection of TSEs and preclinical diagnosis on the basis of anti-PrP MAbs.(23,24) MAbs N6, N16, N24, and N64 could recognize the epitope in recombiant PrP27-30, suggesting that they could be the potential MAbs to detect PrP27-30 and diagnose TSEs. In addition, some anti-PrP MAbs could inhibit PrPsc replication and the MAbs directed against codon regions 90-110 or 145-160 are also antiprion-active in vivo.(25–27) Those anti-PrP MAbs could work as the anti-PrP drugs and provide an alternative method to treat TSEs although the mechanism by which anti-PrP antibodies interfere with PrPsc replication is not clear. In a word, anti-PrP MAbs are very useful reagents in the tests that define various epitopes on PrP, reveal the function of PrPc, probe the mechanism of conformation conversion, diagnose TSEs, and antagonize PrPsc propagation.
Footnotes
Acknowledgments
This work was supported in part by grants from the National Natural Science Foundation of China (no. 30671563) and the Natural Science Foundation of Gansu Province, China (no. 3ZS 051-A25-065) and Science and Technology Planning Project of Gansu Province (2GS 042-A41-001-009).