
Abstract
CHD2 is a member of the CHD family that contains chromodomain, helicase domain as well as DNA-binding domain. The CHD family is involved in gene expression and transcription by ATP-dependent chromatin remodeling. Analysis of mutant mouse revealed that CHD2 is involved in development as well as hematopoiesis, which suggests the involvement of CHD2 in gene expression. However, CHD2 has not yet been analyzed biochemically as there is no specific antibody against it. Here, we report on the establishment of specific monoclonal antibody (MAb) against CHD2 utilizing a rat medial iliac lymph node method. Through cell immunostaining utilizing established MAb to CHD2, we confirmed that CHD2 was localized in euchromatin. Additionally, IP-Western revealed that the expression level of full-length CHD2 did not change during the differentiation stage. Additionally, a specific signal was confirmed around 95 kDa at the undifferentiated stage. This clearly indicated that CHD2 was involved in specific gene expression at this stage. Thus, this antibody can contribute to elucidating the function of CHD2 in cell expression.
Introduction
Chromo (chromatin organization modifier) domain has approximately 50−60 amino acid conservative regions.(2,3) Chromodomain was first identified in HP1 as well as Polycomb, which are chromatin proteins in Drosophila. Chromodomain has been found in various proteins, associating structure with function in eukaryotic chromosomes. Chromodomain proteins are involved in the functional organization in the nucleus through protein-protein interaction and protein-nucleic acid interaction in chromodomain.(4)
Helicase/DNA-binding domain is located in the center of the N-terminal in CHD2. Helicase is classified into five superfamilies.(5) ATP-binding proteins containing helicase domain are composed of Walker A, which has a GxxxxGK[S/T] motif (x is arbitrary amino acid), and Walker B, which has an hhhh[D/E] motif (h is hydrophobic amino acid). CHD2 belongs to the superfamily2, which also contains a number of RNA and DNA helicases. Numbers of superfamily2 migrate on single- or double-strand nucleic acid.(6) In addition, CHD2 has a function to unbind double-strand nucleic acid in an energy-dependent manner, in the same way as the other RNA and DNA helicases.
It is presumable that CHD family proteins function with the help of ATP-dependent chromatin remodeling activity, yet their functions are not the same.(7) Along with CHD1, CHD2 belongs to subfamily1 of the CHD family. It has been reported that CHD1 is involved in the transcription elongation in drosophila and yeast.(8,9) On the one hand, it has been suggested that CHD1 associates with transcriptional co-repressor NCoR.(10) As described above, CHD1 is involved in regulation by promoting and repressing gene expression. It is predicted that CHD2, a homolog of CHD1, is also involved in gene expression. Analysis of chd2 mutant mouse revealed that CHD2 is involved in the development and hematopoiesis.(11–13) Given these facts, CHD2 appears to play a critical role in gene expression. However, the CHD2 protein has not yet been identified and its function has not been sufficiently analyzed. Therefore, to reveal the role of CHD2 in gene expression, we established a specific MAb against CHD2.
Materials and Methods
Cell culture
Mouse skeletal myoblast C2C12 cells were cultured in growth medium (GM), Dulbecco's modified Eagle's medium supplemented with 20% fetal bovine serum (FBS), penicillin (100 U/mL), and streptomycin (100 μg/mL) (Nacalai tesque, Kyoto, Japan) under humidified atmosphere with 5% CO2 at 37°C. C2C12 cells were differentiated in differentiation medium (DM), Dulbecco's modified Eagle's medium supplemented with 2% horse serum (HS), penicillin (100 U/mL), and streptomycin (100 μg/mL) (Nacalai tesque) under humidified atmosphere with 5% CO2 at 37°C.
Production and purification of recombinant proteins
An expression vector of glutathione S-transferase (GST) fusion proteins to CHD2 1313 to 1391 for antigen and P1 (1313 to 1339), P2 (1331 to 1357), P3 (1349 to 1375), and P4 (1367 to 1397) for epitope mapping were transformed into Escherichia coli BL21(DE3) (Novagen, Madison, WI).(14) The E. coli cells were grown in MMI medium containing 50 μg/mL calbenicillin (Nacalai tesque) at 37°C. Expression was induced by the addition of 0.5 mM isopropyl-1-thio-b-D-galactopyranoside (IPTG) (Nacalai tesque) followed by incubation for 12 h at 37°C. Lysed bacteria and purification of the fusion proteins were performed as previously described.(15,16)
Immunization of rat and production of MAb
Anti-CHD2 rat MAbs were generated based on the rat lymph node method established by Sado et al.(17–19) A 10-week-old female lzm rat (Japan SLC, Shizuoka, Japan) was injected via the hind footpads with 500 μL of an emulsion containing 1000 μg of recombinant mouse CHD2 protein and Freund's complete adjuvant. After 2 weeks, the cells from the lymph nodes of a rat immunized with an antigen were fused with mouse myeloma Sp2/0-Ag14 cells at a ratio of 5:1 in a 50% polyethylene glycol (PEG1500, Merck, Darmstadt, Germany) solution. The resulting hybridoma cells were plated onto 96-well plates and cultured in HAT selection medium (hybridoma SFM medium [Invitrogen, Carlsbad, CA]; 10% fetal bovine serum [FBS]; 10% BM-condimed H1 [Roche, Basel, Switzerland]; 100 mM hypoxanthin; 0.4 mM aminopterin; 1.6 mM thymidine). After 7 days post-fusion, the hybridoma supernatants were screened by means of an enzyme-linked immunoadsorbent assay (ELISA) against GST-fused CHD2. Positive clones were subcloned and rescreened by ELISA and immunoblotting. For the preparation of hybridoma supernatants containing highly concentrated antibodies, the resulting positive clones 6D2 and 8H3 were then cultured at a high density using a miniPERM bioreactor (Vivascience, Hannover, Germany).
ELISA
CHD2 antigen (5 μg/mL) in TBS-T (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, and 0.05% Tween-20) was adsorbed on the surface of 96-well flexible microplates (Thermo Scientific, Rockford, IL) by overnight incubation at 4°C. To avoid unspecific binding, the plates were blocked with 1% bovine serum albumin (BSA, Nacalai tesque) in TBS-T. Hybridoma supernatants were incubated for 1 h at room temperature, and then washed with TBS-T three times. The plates were incubated for 30 min at room temperature with alkaline phosphatase-conjugated anti-rat IgG antibody (Sigma, St. Louis, MO) diluted 1:10,000. After washing with TBS-T three times, immunoreactivity was visualized by a pNPP phosphatase substrate system (KPL, Gaithersburg, MD).(20)
Epitope mapping
ELISA was performed with GST protein, GST-CHD21313–1391, and GST-P1 to P4 (5 μg/mL) as antigen. The method was the same as that described in the above ELISA section. Each protein of 50 ng was separated by 12% SDS-PAGE, and electrophoretically transferred to PVDF membranes (Millipore, Bedford, MA).
Immunoprecipitation—Western blot (IP–Western)
C2C12 cells were washed twice with phosphate-buffered saline (PBS) and then lysed in lysis buffer (5 mM pipes, 85 mM KCl, 2 M KCl, 1 mM CaCl2, 5% sucrose, 0.5% NP-40, protease inhibitor cocktail [Nacalai tesque]) using a tissue grinder (Dounce, Wheaton, NJ). Lysates were then centrifuged at 300 g for 5 min. The pellets were re-suspended with IP buffer (50 mM Tris-HCl [pH 7.5], 5 mM MgCl2, 0.5 mM EDTA, 175 mM KCl, 0.5% NP-40, protease inhibitor cocktail). The samples were sonicated for 1 s and digested with micrococcal nuclease (10 U/500 mL) (Worthington, NJ) at 37°C for 15 min. The digested samples were then centrifuged at 3000 g for 10 min. The supernatants were rocked with 100 μL supernatant of rat MAb against CHD2 (6D2, ascites, 1:100) with the irrelevant antibody for 12 h, followed by the addition of protein G-Sepharose (GE Healthcare, Buckinghamshire, United Kingdom) and an additional incubation while rocking for 1 h. Beads were washed three times in IP buffer and eluted with 2x SDS sample buffer. The samples were separated by 6% SDS-PAGE, and electrophoretically transferred to nitrocellulose membranes (Millipore). The membranes were blocked for 1 h at room temperature with a blocking solution containing 5% skim milk (Nacalai tesque) in TBS-T (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, and 0.05% Tween-20), and then incubated overnight at 4°C with anti-CHD2 rat MAb 6D2 diluted in a blocking solution. After washing with TBS-T, the membranes were incubated for 1 h at room temperature with anti-rat IgG conjugated HRP (GE Healthcare). After washing with TBS-T, the membranes were detected by LAS3000 (Fuji Film, Tokyo, Japan).
Immunocytochemistry
C2C12 cells grown on coverslips were washed in PBS at 37°C for 2 min and fixed with 4% paraformaldehyde for 15 min at room temperature. After rapid washing in PBS, cells were permeabilized at 37°C for 10 min in 0.5% Triton X-100 in PBS, then incubated in blocking buffer as described.(21) CHD2 was detected with 6D2, followed by an Alexa 488-conjugated goat anti-rat IgG (Invitrogen) for 1 h at RT. Coverslips were mounted with Prolong (Molecular Probes, Carlsbad, CA). CHD2-expressing cells were visualized using a confocal laser microscope (Carl Zeiss, Jena, Germany).
Results and Discussion
It is expected that CHD2 contains full-length 1827 amino acid sequence. To establish a specific antibody against CHD2, we utilized amino acid sequence from 1313 to 1391 as antigen. According to Blast analysis in other organism species, this amino acid sequence utilized as antigen shares homology with CHD2 protein, whereas this region does not have homology with the other protein. Antigen was expressed in E. coli BL21 as a fused protein with GST. We then generated a MAb against CHD2 utilizing antigen protein GST-CHD21313–1391 by the rat lymph node method. ELISA screening was performed and two positive clones, 6D2 and 8H3, were obtained from hybridoma. Then, epitope mapping was performed to identify which part of the amino acid sequence in CHD2 was recognized by MAbs 6D2 and 8H3.
GST-fused peptide P1 (1313 to 1339), P2 (1331 to 1357), P3 (1349 to 1375), and P4 (1367 to 1397), and quartered GST-CHD21313–1391 utilized for generating antibody against CHD2, were expressed in E. coli BL21. ELISA and immunoblotting were performed to evaluate epitope region of 6D2 and 8H3 (Fig. 1). The result confirmed that 6D2 recognized the amino acid sequence of 1376 to 1391 as it showed strong reaction against P4 (1367 to 1397). Furthermore, 8H3 had an epitope region in the amino acid sequence in P4 (1367 to 1397) (data not shown).
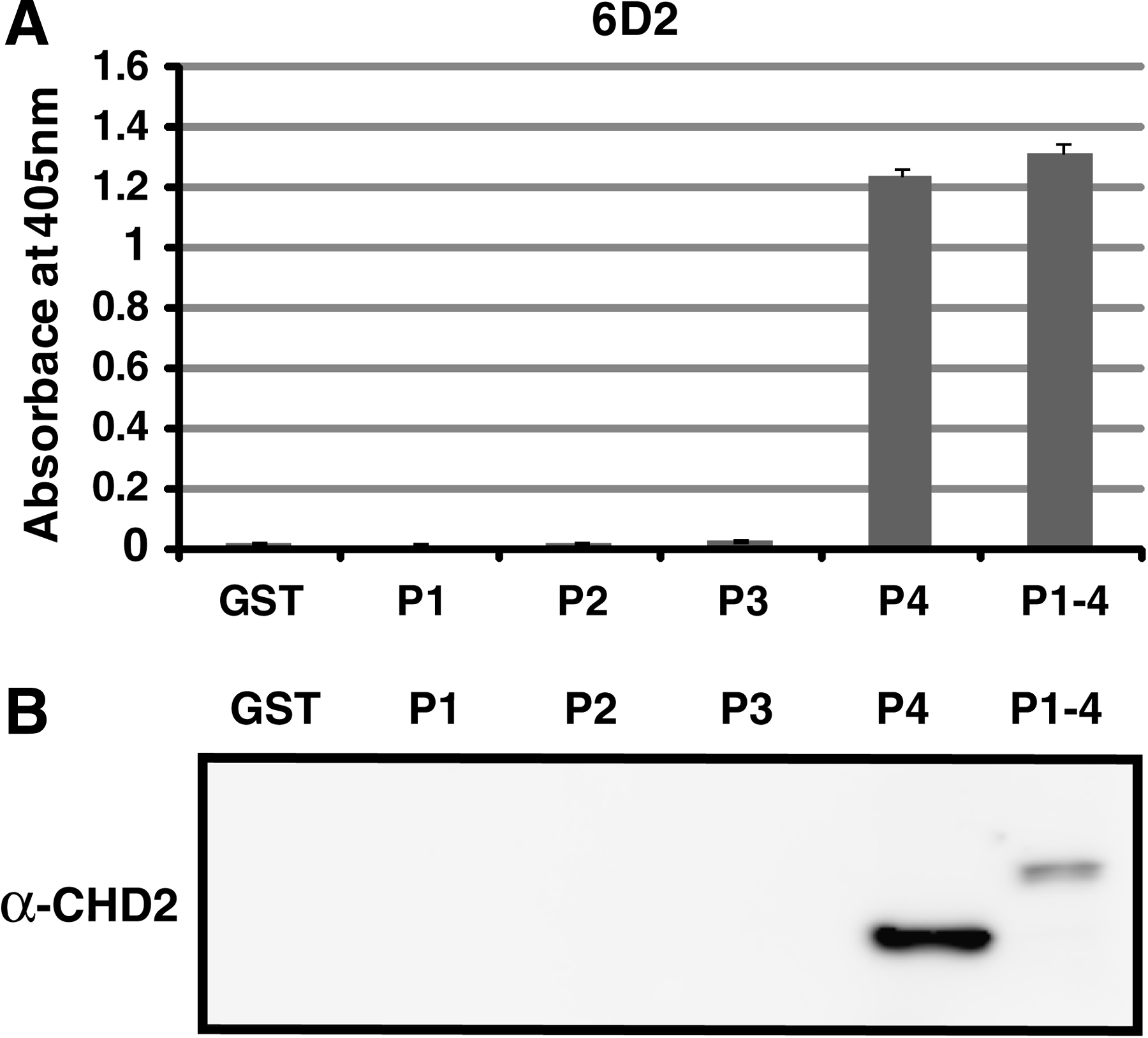
Epitope mapping of MAb 6D2. Epitope of MAb was detected by ELISA (
This indicates that CHD1 was co-localized with SSRP1, which is an HMG box-containing protein and is bound with modified DNA.(22) Therefore, it is conceivable that CHD2 protein is localized in the nucleus. Cell immunostaining was performed to evaluate the nuclear localization of CHD2 protein and the results confirmed that CHD2 did localize in the nucleus (Fig. 2A). Additionally, it was observed that localized CHD2 covered condensed chromatin by shaping retiform in the nucleus. It has been reported that CHD subfamily1 containing CHD2 has a DNA-binding domain. Therefore, as CHD2 is tightly bound with DNA in the nucleus, CHD2 was localized in euchromatin region and poorly stained by Hoechst staining before and after differentiation. These facts suggest that CHD2 is involved in transcription.
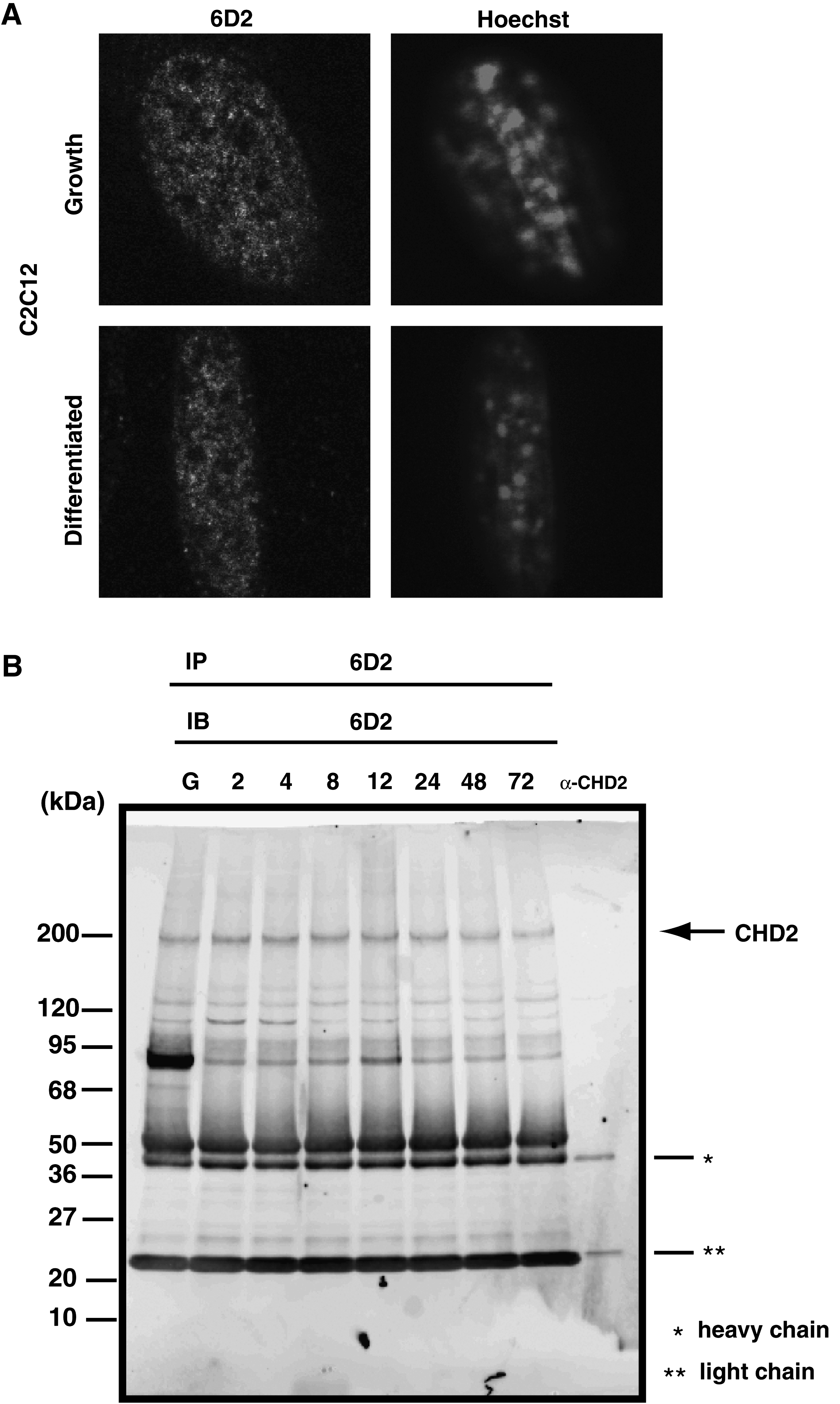
CHD2 is localized to nuclei in C2C12 cells. (
CHD2 molecular weight is presumably 210 kDa. IP–Western was performed to verify whether the obtained positive clones 6D2 and 8H3 could identify CHD2 (Fig. 2B). The result showed that 6D2 strongly reacted around 200 kDa in the nuclear extract of skeletal muscle precursor cell C2C12. It was confirmed that 6D2 could identify deducted full-length CHD2 protein. Additionally, detected signals remained unchanged from the undifferentiated state until 72 h after differentiation induction. This indicated that CHD2 existed before and after differentiation in the nucleus. In addition, a specific strong signal was observed around 95 kDa in the undifferentiated IP samples. Thus, we expect that the specific MAbs 6D2 and 8H3 against CHD2 established in this study are useful for the biochemical analysis of CHD2 in the future.
Footnotes
Acknowledgments
We thank Dr. Yoshikazu Sado (Shigei Medical Institute) for his advice. This work was supported, in part, by grants from the Ministry of Education, Culture, Sports, Science and Technology of Japan.