
Abstract
The X-box binding protein-1(XBP-1) is a 29 kDa protein belonging to the basic region/leucine zipper (bZIP) family of transcription factors. Previous studies showed that XBP-1 mediated a wide range of responses in B-cell differentiation, unfolded protein response (UPR), and tumorigenesis.(1–4) For these reasons, it is believed that XBP-1 would be a novel therapeutic target in some pathogenic processes. In this study, a set of XBP-1 MAbs were raised and cloned. Then it was proven that some clones among them could be used in Western blot analysis, immunocytochemistry, or immunohistochemistry. Therefore, the obtained MAbs not only provided new powerful tools for investigation of expression profile and functions of XBP-1 protein, but also provided the possibility of generating gene engineering antibodies specific for targeting XBP-1.
Introduction
Although XBP-1 RNA transcript expressions have been revealed by in situ hybridization in mouse and human tissues, expression profiles of XBP-1 at the protein level have not been fully demonstrated. This is partially due to the lack of high affinity MAbs suitable for immunohistochemical staining. For this reason, based on the successful immunization with expressed GST-hXBP fusion protein and followed by cell fusion, three ELISA-positive MAbs, named hXBP1.1–hXBP1.3, have been cloned. Subsequent experiments showed that the set of MAbs could be used in Western blot analysis, immunocytochemistry, and immunohistochemistry. Thus, our preliminary work provides novel powerful tools for investigation of the expression profile of XBP-1 protein and furthers the study of XBP-1 function.
Materials and Methods
Expression and purification of human XBP-1
Full-length wild-type human XBP-1 (hXBP-1) cDNA was subcloned into the pGEX-4T-1 vector and expressed as a glutathione S transferase (GST) fusion protein in BL21 E. coli following induction with IPTG (0.5 mM) for 6 h at 32°C. Soluble GST-hXBP-1 protein was recovered from bacterial cell lysates on glutathione S-Sepharose (Amersham Biosciences, Piscataway, NJ), washed three times in lysis buffer, and eluted with 20 mM glutathione. GST-hXBP-1 was then dialysed into phosphate-buffered saline (PBS) and used as antigen to immunize mice and to screen the monoclonal antibodies.
Production of hybridomas
Female BALB/c mice (8 weeks old) were immunized with 20 μg of GST-hXBP-1 in complete Freund's adjuvant and applied to the mice by subcutaneous (s.c.) injection. Subsequently, immunizations were carried out two more times with 20 μg of recombinant proteins mixed with incomplete Freund's adjuvant by s.c. injection at 4-week intervals. The total titer of anti-GST-hXBP-1 antibodies in blood sera was monitored by ELISA.
At the next stage, one of the immunized mice was boosted with an additional 20 μg of antigen by intraperitoneal (i.p.) injection in phosphate-buffered saline (PBS) without adjuvant. Three days later, splenocytes from the boosted mouse and SP2/0 myeloma cells cultured in RPMI 1640 medium containing 20% fetal calf serum (FCS) were fused in the presence of PEG (MW4000, Merck, Darmstadt, Germany) and cultured in RPMI 1640 medium containing 20% FCS and HAT (Invitrogen, Carlsbad, CA). The positive hybrids were selected by ELISA and then subcloned three times using the limiting dilution method.(11)
ELISA assays
96-well plates were incubated with 0.5 μg/well of GST-hXBP-1 or with GST protein as the control in PBS (pH 7.4) for 16 h at 4°C. The plates were then washed three times with PBS containing 0.1% Tween-20. Subsequently, the plates were loaded with 100 μL aliquots of hybridoma media and were incubated for 1 h at 37°C followed by four washes with PBS containing 0.1% Tween-20. After washing, 100 μL of horseradish peroxidase (HRP)–conjugated goat anti-mouse IgG antibodies (1:5000 v/v, Promega, Madison, WI) were added to each well and plates were incubated at 37°C for 1 h. After three more washes, 100 μL of TMB substrate were added to each well. The reaction was stopped by the addition of 50 μL of 2.5 M H2SO4 to each well and plates were read at 450 nm on a micro-plate reader.
Production and purification of monoclonal antibodies
BALB/c mice were injected with 0.2 mL of pristane and 7–10 days later inoculated with 1 × 106 hybridoma cells. The ascitic fluid was collected after 7–14 days. The fraction of immunoglobulins was precipitated from ascetic fluid with 50% ammonium sulphate and used for affinity purification by Protein G-Sepharose CL-4B (GE Healthcare, Piscataway, NJ) chromatography. The IgG fractions were pulled together and dialyzed in PBS (pH 7.4). The aliquots of purified antibodies were stored at −70°C.
Western blot analysis
Cell lysates of Ramos cells were prepared by using lysis buffer containing 20 mM Tris-HCl (pH 7.5), 150 mM NaCl, 1% Triton X-100, 50 mM NaF, 5 mM EDTA, and a mixture of protease inhibitors (Roche Molecular Diagnostics, Meylan, France). Recombinant GST-hXBP-1 protein (1 μg) or cell lysates (30 μg) was electrophoresed on 12% sodium dodecyl sulphate–polyacrylamide electrophoresis gel and transferred onto PVDF membrane (Millipore, Billerica, MA). The membranes were blocked with 5% skim milk in 25 mM Tris-HCl (pH 7.4), 0.8% NaCl, 0.02% KCl, and 0.05% Tween-20 (TBST) for 1 h at room temperature (RT) and then incubated with anti-hXBP-1 MAbs (diluted to 3 μg/mL) or rabbit anti-hXBP-1 PAb (Santa Cruz Biotechnology, Santa Cruz, California) overnight at 4°C. After washing three times with TBST, HRP-conjugated goat anti-mouse IgG (1:5000 v/v; Promega) were incubated with the membrane for 1 h at RT. Finally, the membrane was detected by Immun-Star HRP Chemiluminescent Kit, images were captured, and the intensity of the bands was quantitated with the Bio-Rad VersaDoc image system (Bio-Rad, Hercules, CA).
Gene transfection
CHO cells were transfected with pCMV-4-hXBP-1 full-length cDNA or pCMV-4 cDNA with transfection reagent Lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. Briefly, CHO cells at 1 × 106 per well in 6-well plates were seeded and incubated for 12–24 h. The DNA-Lipofectamine 2000 complexes were added directly to the wells and mixed gently. The CHO cells were incubated at 37°C in 5% CO2 for another 48 h, and the cells were harvested for immunocytochemistry analysis.
Immunocytochemistry and immunohistochemistry
Pallets of pCMV-4-hXBP-1 transfected CHO cells or pCMV-4 transfected CHO cells were fixed in the 4% paraformaldehyde over 24 h followed by dehydration in gradient ethanol and were made into paraffin block. Then the paraffin blocks were cut at 5 μm thicknesses and mounted on slides. Eight cases of formalin-fixed paraffin-embedded human hepatocellular carcinoma (HCC) biopsies were retrieved from the Department of Pathology at Xijing Hospital (Xi'an, China). Endogenous peroxidase activity of slides of cell block and human hepatocellular carcinoma was blocked with methanol containing 3% H2O2 for 10 min, and non-specific binding was blocked by normal goat serum (10%) for 10 min. The slices were dipped into anti-hXBP-1 MAbs (supernatant) for 2 h at RT. After three washes in PBS, the slices were dipped into biotin-conjugated goat anti-mouse IgG (Promega) for 1 h at RT, followed by streptavidin-HRP complex incubation for another 1 h. Then antibody complexes were visualized by incubation with DAB chromogen. Sections were counterstained with Mayer's hematoxylin for 30 s, dehydrated through gradient ethanol, cleared in dimethyl benzene, mounted, and examined using light microscopy. Normal mouse IgG were applied as negative control.
Results
Generation and characterization of MAbs
Three positive hybridoma clones secreting MAbs against human XBP-1 were obtained and designated hXBP-1.1, hXBP-1.2, and hXBP-1.3, respectively. The immunoglobulin isotype of these MAbs was IgG1 (κ). These hybridoma clones could stably secrete specific MAbs, which reacted with GST-hXBP-1, but not GST protein in ELISA. The MAbs were produced as ascites and purified using affinity purification by Protein G-Sepharose CL-4B.
Screening of MAbs against recombinant hXBP-1 protein by Western blot analysis
Three positive hybridoma clones (designated hXBP-1.1, hXBP-1.2, and hXBP-1.3) were produced as ascites and further tested in immunoblotting with recombinant GST-hXBP-1. Immunoblotting with rabbit anti-hXBP-1 PAbs revealed an ∼55 kDa band from the recombinant protein (Fig. 1). It was found that hybridoma hXBP-1.1 and hXBP-1.2 could blot a band of the same molecular weight as PAbs tested, whereas there were no positive signals when hXBP-1.3 was used to blot the GST-hXBP-1 (Fig. 1).

Western blot analysis of reactivity of MAbs with recombinant hXBP-1 proteins. Western blot analysis with a rabbit anti-hXBP-1 PAbs revealed an ∼55 kDa band from the recombinant protein (lane 1). Two of the ELISA-positive hybridomas (hXBP-1.1 and hXBP-1.2) specifically blotted a band of the same molecular weight as PAbs tested (lanes 2 and 3).
Screening of MAbs against endogenous hXBP-1 protein by Western blot analysis
We used the same methodology to test the reactivity and specificity of the hXBP-1 MAbs in Ramos cell nuclear extracts that endogenously express XBP-1. Cultured Ramos cells were harvested and prepared using lysis buffer. Cell lysates (30 μg) were separated by SDS-PAGE and transferred onto PVDF membrane. After exposure to PAbs or three ELISA-positive MAbs, there was a band of ∼29 kDa specifically recognized by PAbs, hXBP-1.1 and hXBP-1.2 (Fig. 2).

Western blot analysis of reactivity of MAbs against endogenous hXBP-1 protein. Western blot analysis with a rabbit anti-h hXBP-1 PAbs revealed ∼29 kDa band (lane 1). Two of the ELISA-positive hybridomas (hXBP-1.1 and hXBP-1.2) specifically blotted a band of the same molecular weight as PAbs tested (lanes 2 and 3).
Application of MAbs in immunocytochemistry
hXBP-1.1, hXBP-1.2, and hXBP-1.3 were tested in immunocytochemistry respectively. pCMV-4-hXBP-1 transfected CHO cells or pCMV-4 transfected CHO cells were grown in RPMI 1640 containing 10% FCS and harvested by centrifugation. The cell pallets were fixed, dehydrated, and made into paraffin block. Then the paraffin block was sectioned for immunocytochemistry. As shown in Figure 3, hXBP-1.2 antibody recognized endogenous XBP-1 in pCMV-4-hXBP-1 transfected CHO cells, which is visualized by strong DAB staining in the cytoplasm (Fig. 3A) in comparison with that in pCMV-4 transfected CHO cells
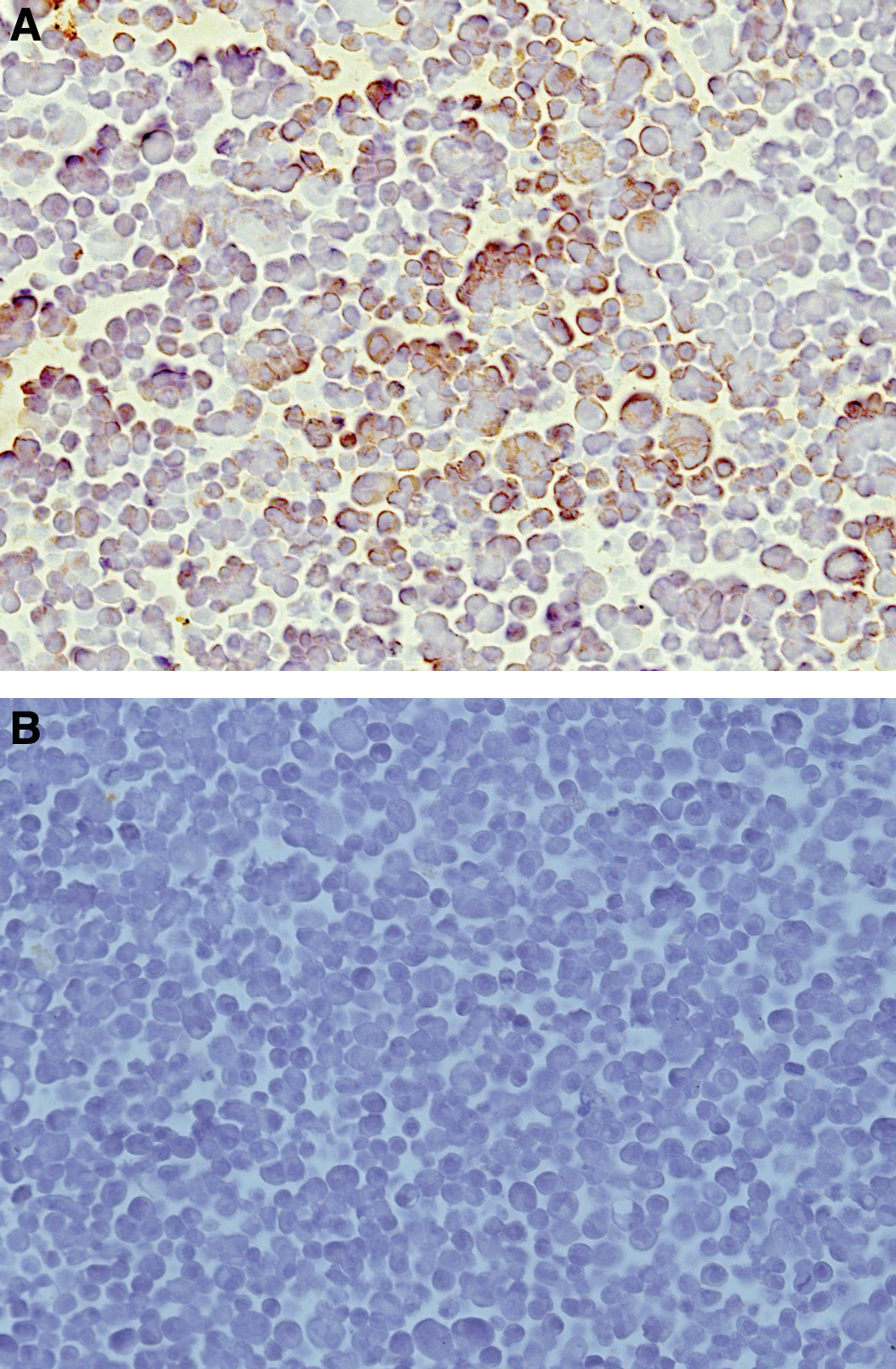
CHO cells immunostaining for XBP-1.
Localization of XBP-1 immunoreactivity
XBP-1 was expressed in six of eight HCC samples. The positive signals localized in the cytoplasm, with no immunostaining in the nuclei. Moderate to strong cytoplasmic staining was detected in well-differentiated HCC tissues (Fig. 4A), whereas the poorly differentiated HCC tissues exhibited weak or negative immunostaining for XBP-1. The negative control showed no positive staining (Fig. 4B).
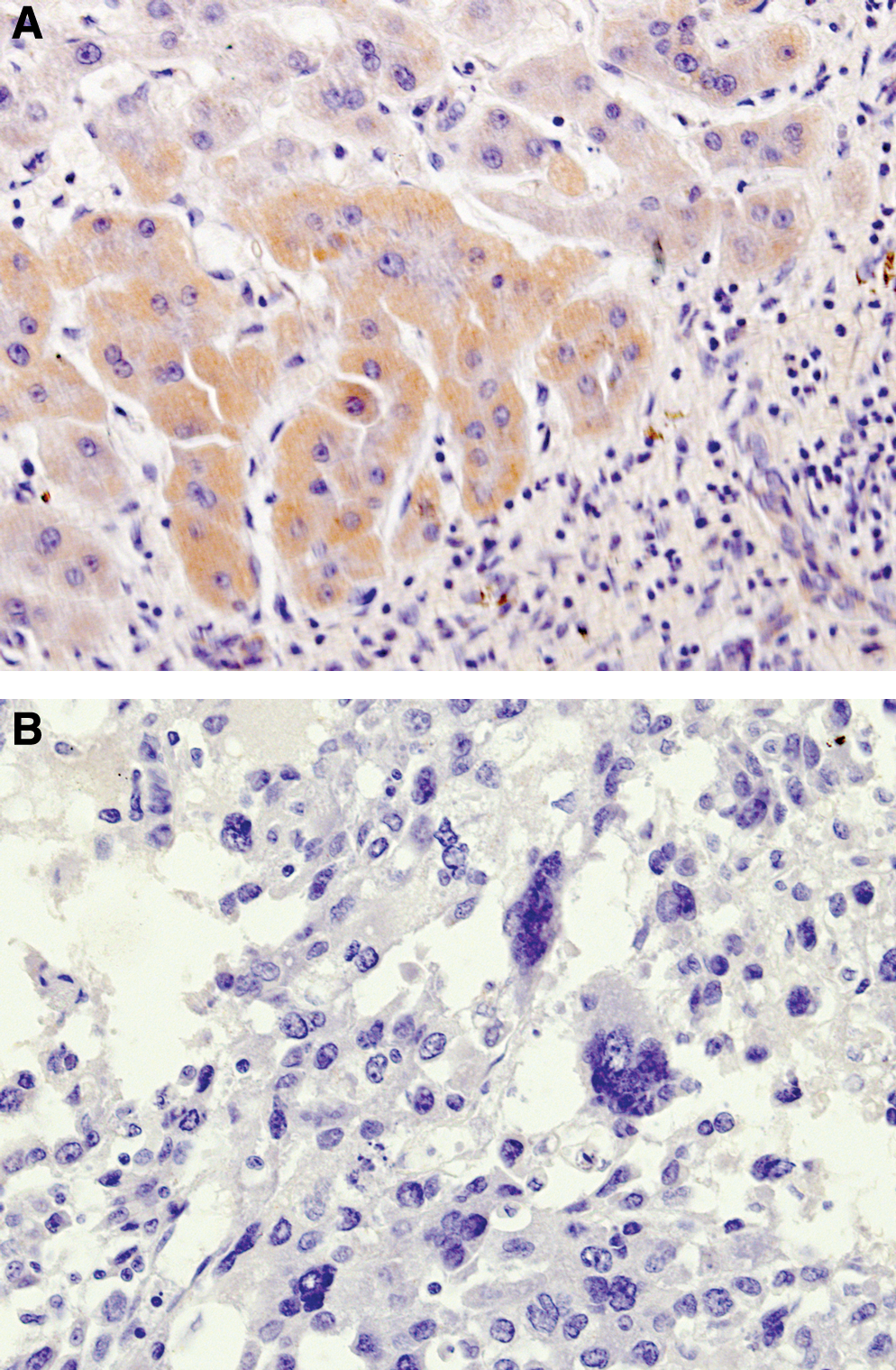
Immunohistochemical detection of XBP-1 in human HCC tissues using anti-XBP-1 MAb (hXBP-1.2).
Discussion
XBP-1 was first identified as interleukin-6 target genes in human multiple myeloma cells in 1999.(12) Later it was found that XBP-1 is one of the major transcription factors in plasma cell differentiation, and its loss leads to an almost complete absence of plasma cells and circulating immunoglobulins.(13) Further study showed that XBP-1 signaling defects led to aberrant expression of the plasma cell transcription factors IRF4 and Blimp-1, and altered levels of activation-induced cytidine deaminase and sphingosine-1-phosphate receptor, which resulted in B-cell failure to signal effectively through the B-cell receptor.(14)
In addition to its role in the terminal differentiation of B lymphocytes into plasma cells, the XBP-1 gene is activated in response to endoplasmic reticulum stress signals, which generate a nuclear protein that acts as a potent transactivator and regulates the expression of genes related to the unfolded protein response (UPR).(15,16) UPR allows cells to tolerate conditions of endoplasmic reticulum (ER) stress caused by misfolded proteins. It has been proven that XBP-1 is a critical transcriptional activator during UPR and is responsible for regulating the function of genes in tumor cell survival.(2) Meanwhile it has been found that over-expression of XBP-1 in tumor cells could confer drug resistance by preventing drug-induced cell-cycle arrest and mitochondrial permeability and apoptosis, while downregulation of XBP-1 increases the sensitivity to killing by hypoxia.(17)
For the important roles of XBP-1 in B cell differentiation, UPR, and tumorigenicity, novel tools are needed to explore its expression, distribution, and functions during physiological and pathogenic processes. In this study, a set of XBP-1 MAbs has been raised. Among them, two could be used in Western blot analysis, and one could be used in immunocytochemistry and immunohistochemistry. These MAbs provide new powerful tools for the investigation of the expression profile of XBP-1 protein and the study of XBP-1 function.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (nos. 30772074; 30872319).