
Abstract
Botulinum neurotoxins (BoNTs) are extremely potent toxins that can contaminate foods and are a public health concern. Anti-BoNT antibodies have been described that are capable of detecting BoNTs; however there still exists a need for accurate and sensitive detection capabilities for BoNTs. Herein, we describe the characterization of a panel of eight monoclonal antibodies (MAbs) generated to the non-toxic receptor-binding domain of BoNT/A (HC50/A) developed using a high-throughput screening approach. In two independent hybridoma fusions, two groups of four IgG MAbs were developed against recombinant HC50/A. Of these eight, only a single MAb, F90G5-3, bound to the whole BoNT/A protein and was characterized further. The F90G5-3 MAb slightly prolonged time to death in an in vivo mouse bioassay and was mapped by pepscan to a peptide epitope in the N-terminal subdomain of HC50/A (HCN25/A) comprising amino acid residues 985WTLQDTQEIKQRVVF999, an epitope that is highly immunoreactive in humans. Furthermore, we demonstrate that F90G5-3 binds BoNT/A with nanomolar efficiency. Together, our results indicate that F90G5-3 is of potential value as a diagnostic immunoreagent for BoNT/A capture assay development and bio-forensic analysis.
Introduction
The potential severity and lethality of botulism poisoning require that a patient presenting clinical symptoms receive antitoxin in a timely manner, before receiving a definitive diagnosis.(2,6,7) To date, the gold standard to detect BoNT in clinical samples is the mouse toxicity assay. While both sensitive and specific, this assay takes up to 96 h to complete and requires specialized personnel, facility, and reagent resources.(8) Due to adverse patient reactions that can occur and batch variation in polyclonal immunoglobulin production, it is desirable to develop rapid methods of diagnosis that permit accurate and sensitive BoNT detection and serotyping capability to ensure the correct course of treatment is initiated as soon as possible. Many efforts have been made to develop alternative, immunologically based assays for BoNT detection in clinical samples, yet these assays are often limited by the availability of high-quality antibodies.(8)
The production of monoclonal antibodies (MAbs) against BoNTs is an excellent choice for several reasons. First, MAbs are consistent between lots. Second, they are well defined and are theoretically in unlimited supply. Mammalian cell lines scale up readily and provide a means to produce clinical grade material. Third, murine hybridomas with bioactivity provide good lead molecules that can be modified by recombinant means for improved bioactivity and/or less immunogenicity, the latter, for example, by simple chimerization strategies.
Investigation of BoNT structures has provided invaluable information for alternative strategies for antibody development to the BoNTs. All known BoNTs are synthesized as inactive ∼150 kDa pretoxins composed of three ∼50 kDa functional domains. Upon secretion, the pretoxin is cleaved by proteases into an active disulfide bond-linked dipeptide consisting of a ∼50 kDa light chain and a ∼100 kDa heavy chain. The light chain contains the N-terminal catalytic domain, while the heavy chain comprises a ∼50 kDa translocation domain (HN50) and a ∼50 kDa C-terminal receptor-binding domain (HC50). The catalytic domain on the light chain is a zinc-endopeptidase that cleaves target SNARE (soluble NSF attachment protein receptor) complex proteins at the neuromuscular junction, preventing neurotransmitter release. The HC50 domain contains the receptor-binding site that targets the BoNTs to cholinorogenic nerve endings where the toxin is internalized via receptor-mediated endocytosis, and the HN50 translocation domain aids in the intracellular translocation of the enzymatically active light chain into the host cell cytosol.(9–11)
This HC50 binding domain (corresponding to residues 873-1295 of BoNT/A) can be further divided into two ∼25 kDa subdomains: the N-terminal β-barrel (HCN25 subdomain) and the C-terminal β-trefoil fold (HCC25 subdomain).(11,12) BoNTs are proposed to bind to host cell neurons via a two-receptor mechanism, whereas BoNT/A binds to host cell gangliosides followed by specific binding to the glycoprotein receptor SV2 within the synaptic vesicles.(13–15) The conserved motif H…SXWY…G for ganglioside binding is located within HCC25 of BoNT/A and BoNT/B.(11,16,17) While the specific amino acids for BoNT/A receptor binding have yet to be elucidated, the BoNT/B receptor-binding site is located within its HCC25 subdomain, and it is most likely this region that provides the BoNT/A protein receptor site as well.(14,18) Immunization with the inert HC50 domains of BoNTs provides a safe and rational approach for MAb development toward BoNTs.
Disruption of BoNT binding to host cell receptors negates toxicity, and the bioactivity of neutralizing MAbs to the HC50 receptor-binding domain of BoNTs is well-established.(19–26) A limited number of neutralizing MAbs specific for the BoNT/A HC50 binding domain, HC50/A, have been characterized in detail.(20,24,27,28) Three of these neutralizing antibodies were generated by phage display, of which two were developed from a murine immune library and one was developed from a human library.(20,27) Several studies on neutralizing MAbs specific for BoNT/A indicate that an oligoclonal mixture of neutralizing antibodies provide a rational design for a potent human therapeutic.(20,25,26,29,30) Given that each of the seven BoNTs exists as multiple antigenic variants,(31) one can expect that individual MAbs may have limited in vivo protection and pools of well-defined MAbs will be required for optimal protection. There have also been reports of murine MAbs, developed using classical hybridoma technology, which neutralize BoNT/A with high potency.(25,30) Thus, the potential for development of neutralizing MAbs from murine immune sources to generate lead molecules against BoNT/A and other serotypes clearly exists.
Herein, we describe the development of a panel of MAbs generated using a recombinant non-toxic HC50 binding domain of BoNT/A and characterize the epitope of MAb F90G5-3, an antibody that binds BoNT/A with nanomolar efficiency that may serve as an important immunoreagent for diagnostic assay development.
Materials and Methods
Clostridium botulinum neurotoxin immunogens and antigens
Gene segments of BoNT/A (strain 62A; amino acids [aa] 861-1296; accession no. X52066), BoNT/B (strain Okra; aa 853-1291; accession no. M81186), and BoNT/E (strain NCTC 11219; aa 840-1252; accession no. X62683) were used to clone and express recombinant HC50/A, HC50/B, and HC50/E binding domains as previously described.(19) Antibody reactivity with whole BoNT was assessed using available commercial plates (Metabiologics, Madison, WI) in an ELISA protocol. The whole BoNT toxins correspond to BoNT/A from strain Hall A, BoNT/B from strain Okra B, and BoNT/E from strain Alaska E.
Cloning, expression, and purification of BoNT/A HC50 subdomains HCN25/A and HCC25/A
Gene segments encoding the N-terminal half (HCN25/A; aa 865-1100) and the C-terminal half (HCC25/A; aa 1077-1296) of the BoNT/A HC50 receptor-binding domain (strain 62A; aa 865-1296, accession no. M30196
Immunization of mice and generation of hybridoma cell lines
Immunization of mice and generation of the hybridoma cell lines described herein was conducted as described.(32) MAbs were first cloned from semi-solid agarose and then clones of interest underwent another 2 rounds of limiting dilution subcloning to ensure they were monoclonal. The MAb isotypes were determined using a commercial dipstick assay (Roche, Indianapolis, IN). The MAbs were manually purified from serum-free cell supernatant using HiTrap protein G columns (GE-Healthcare Bio-Sciences), according to the manufacturer's recommendations. MAb and antigen protein concentrations were conducted using BCA reagent assay (Pierce).
ELISA
Reactivity of the antibodies via ELISA was conducted following the standard operating procedures described previously by Corbett and colleagues.(32) For lower limit of detection, HC50/A was coated in a concentration range from 125 ng to 122 fg. The lower limit of detection was defined at the lowest concentration of antigen where binding was observed with a value at least four times greater than background. For competitive ELISA, 62.5 ng/mL F90G5-3 was incubated with 10 μg/mL BSA, BoNT/A HCC25/A, or HCN25/A overnight at 4°C with constant end-over-end rotation. The pre-incubated MAb was then diluted 2-fold across a 96-well plate coated with 100 ng/well HC50/A and the ELISA processed as above. Whole toxin ELISAs were conducted using plates coated with BoNT/A, BoNT/B, or BoNT/E, per the manufacturer's recommendations (Metabiologics).
SDS-PAGE and Western immunoblotting
SDS-PAGE gels and Western immunoblots were performed essentially as described previously.(33) Briefly, 2 μg of HC50/A, HC50/B, HC50/E, HCN25/A, HCC25/A, or irrelevant antigen (bovine serum albumin) were electrophoresed on a 4–20% Criterion Precast polyacrylamide gel (Bio-Rad, Hercules, CA) followed by electrophoretic transfer to Immobilon-P membranes (Millipore, Bedford, MA). The immunoblots were blocked using 0.4% BSA and washed three times with Tris-buffered saline 0.1% Tween-20 (TBS-T). Bound antibody was detected by incubation with HRP-conjugated goat anti-mouse IgG (H+L) antibody (Southern Biotech, Birmingham, AL), diluted 1:1000 with 0.1% skim–0.1% TBS-T, and developed using 4-chloro-1-napthol substrate (Sigma), according to the manufacturer's instructions.
Surface plasmon resonance
The measurement of the affinity of the MAb for BoNT/A HC was performed essentially as described previously,(34–36) using a Biacore 2000 (Biacore, Uppsala, Sweden). BIAevaluation 3.2 software was used to measure and plot the kon and koff values directly, which were then used to calculate the binding affinity (KD).
Pin peptide epitope mapping
Peptides covering the sequence of BoNT/A HC were synthesized as 15-mers, overlapping by five residues, coupled to nylon support pins in a 96-well format (Pepscan Systems, Lelystad, Netherlands). All manipulations of the pin peptide assemblies were performed by placing the tips of the pins in the wells of ELISA plates (MaxiSorp, Nalge-Nunc, Rochester, NY). The pins were blocked with 4% BSA/PBS for 1 h at room temperature, followed by incubation with MAb F90G5-3 (diluted in 2% BSA/PBS) for 1 hour at room temperature, and washed three times in 0.9% saline–0.05% Tween-20 with moderate shaking. To detect MAb binding the pins were incubated with secondary antibody and developed as above for Western immunoblot. Pins were regenerated according to the manufacturer instructions for re-use.
Mouse protection assay
The functional activity of MAb reactive with whole BoNT/A was assessed using a mouse protection assay (MPA) as previously described.(37) The MPA was conducted following the guidelines of the Institutional Animal Use and Care Committee of Thomas Jefferson University (Philadelphia, PA). The MAb-mediated protection was assessed in vivo by mixing 50 μg MAb with 5LD50 of BoNT/A toxin, incubating at ambient temperature for 1 h, followed by injection into the tail vein of 25 gm Swiss Webster mice (n=5); this was followed by monitoring percent survival for 5 days.
Results
MAb development and characterization
Spleens from mice immunized with the recombinant BoNT/A HC50/A fragment were used in two hybridoma fusion procedures. The two fusions resulted in the development of eight anti-HC50/A MAb producing hybridomas against BoNT/A HC50 receptor binding domain of BoNT/A(32) (Fig. 1). All of the MAbs, except F90G5-3 (IgG2a/κ), were of the IgG1/κ isotype. Assessment of the BoNT/A reactivity revealed only one MAb, F90G5-3, had strong reactivity with whole BoNT/A neurotoxin, and was therefore chosen for further characterization (Fig. 1). The specificity and reactivity of MAb F90G5-3 were assessed against the HC50 domains of BoNT/A, BoNT/B, BoNT/E,(32) and the BoNT/A HC50 subdomains, HCN25/A and HCC25/A (Fig. 2A and C). As we demonstrated previously in a high-throughput method development study, F90G5-3 has excellent serotype specificity for HC50/A domain with no observed cross-reactivity with HC50/B or HC50/E.(32) Indeed, the MAb was reactive with the HCN25/A subdomain and no reactivity was observed for the HCC25/A subdomain via ELISA or denaturing Western immunoblot analysis (Fig. 2A and C, respectively). The lower limit of detection of F90G5-3 for BoNT/A HC fragment in indirect ELISA was about 8 ng when assayed using ELISA conditions employed within this study (Fig. 2B).
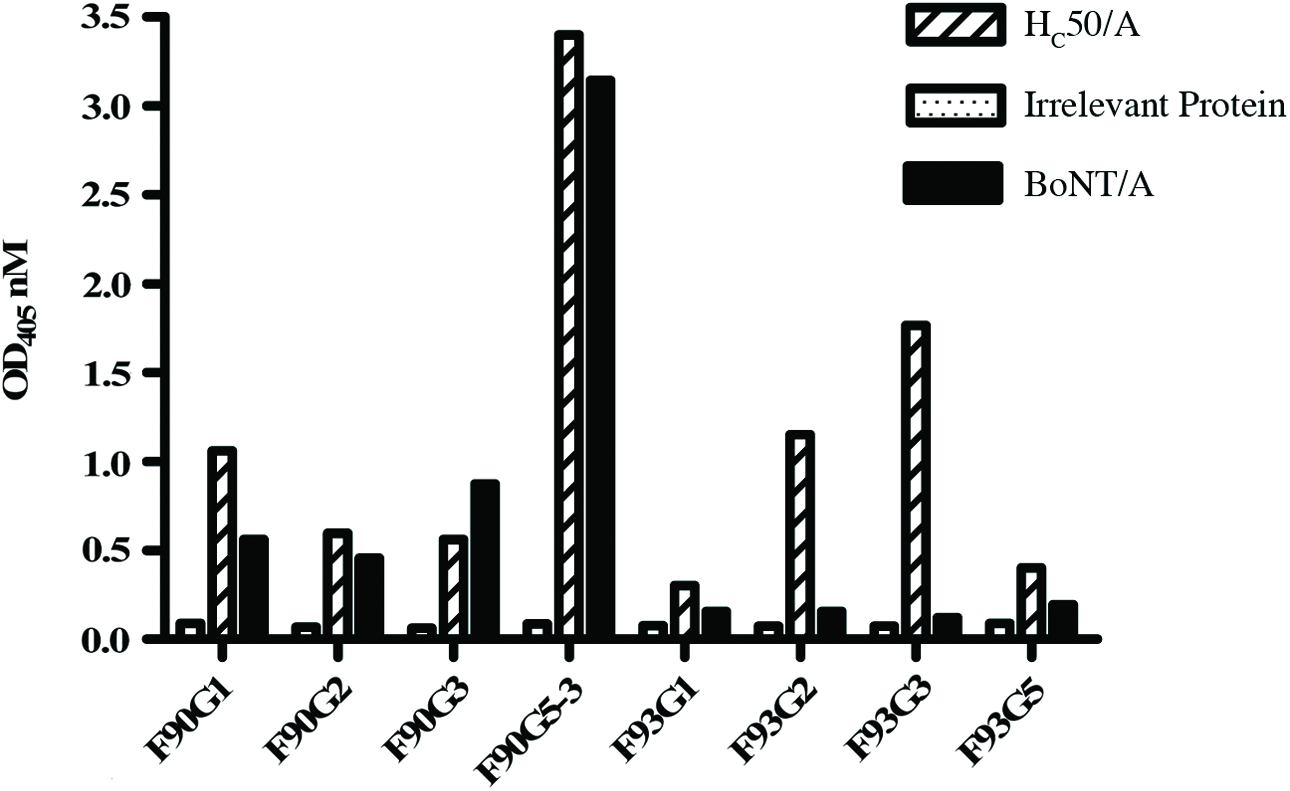
Reactivity of MAbs developed against the HC50 binding domain of BoNT/A. Anti-HC50/A MAb reactivity assessed by ELISA, as described previously.(32) One clone, F90G5-3, elicited strong reactivity with whole BoNT/A and was chosen for further characterization.
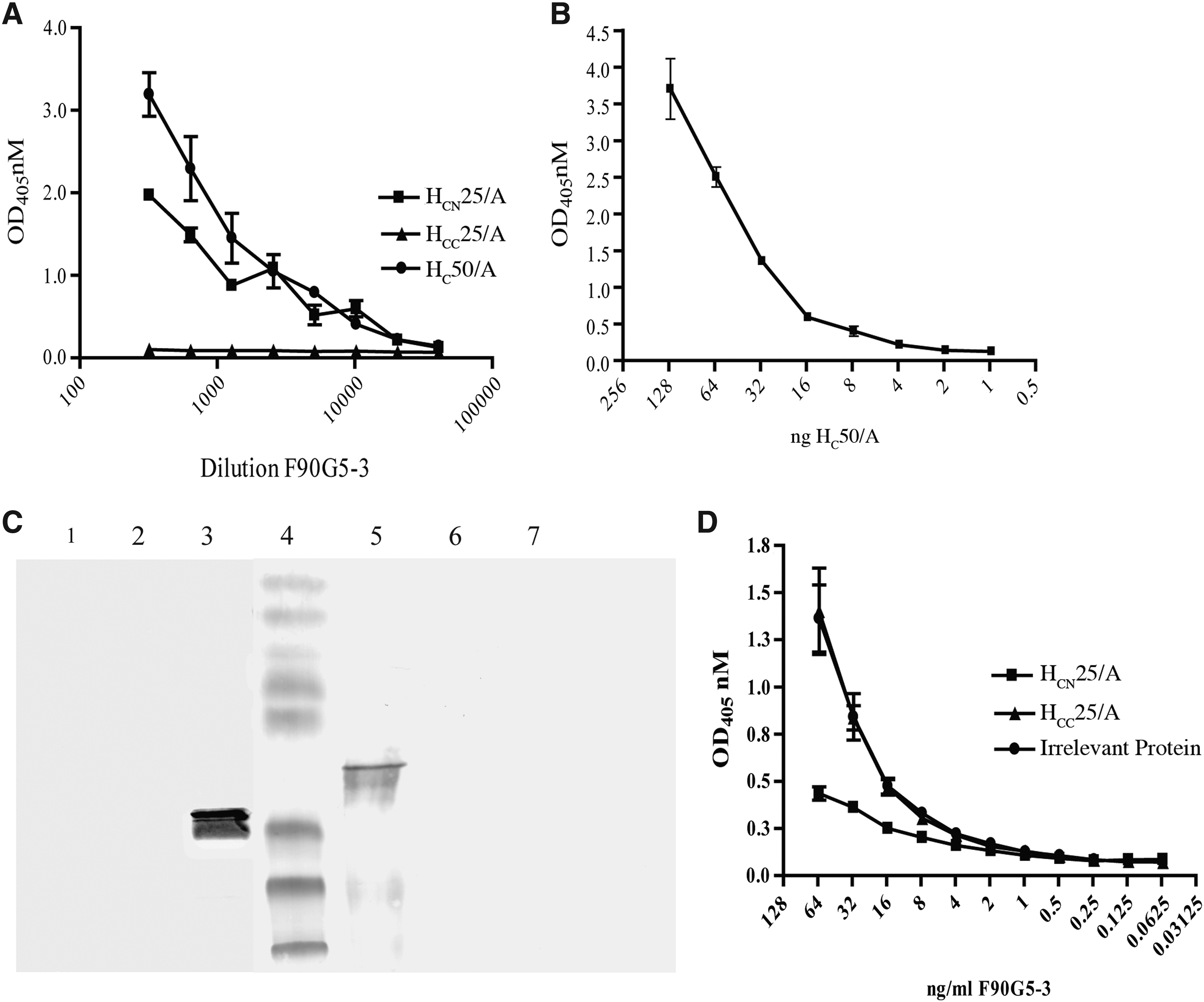
Reactivity and specificity of MAb F90G5-3. (
Reactivity of MAb with F90G5-3 with HCN25/A (as demonstrated in ELISA and Western immunoblot analysis) indicates that the antigenic site for F90G5-3 resides in the N-terminal HCN25/A fragment. To further assess the specificity of F90G5-3 for the HCN25/A subdomain, a competition ELISA was conducted. Indeed, decreased reactivity of F90G5-3 for HC50/A was observed when the MAb was pre-incubated with HCN25/A, and no competition was observed with pre-incubation of F90G5-3 with HCC25/A or irrelevant protein (BSA) (Fig. 2D).
The reactivity of F90G5-3 with denatured HC50/A and HCN25/A also suggests that the antibody binds to a linear epitope. To further define the linear epitope recognized by F90G5-3, pin peptide mapping was conducted where overlapping pin peptides covering the sequence of BoNT/A HC50 receptor-binding domain (corresponding to BoNT/A aa residues 865-1296) were utilized. The data showed that F90G5-3 predominantly bound to the peptide epitope corresponding to amino acids 985WTLQDTQEIKQRVVF999 (Fig. 3), which is found in both the HC50/A and HCN25/A recombinant proteins.
Analysis of the HC50/A epitope bound by MAb F90G5-3. (
The MAb F90G5-3 binds to the HC50/A fragment with nanomolar affinity. Measurement of the affinity of MAb F90G5-3 for HC50/A was performed via surface plasmon resonance analysis. The kon and koff rates were 4.8×103/Msec) and 1.5×10−4/sec, respectively, and the resultant KD was 3.5±1.1 nM. This is typical of affinity matured MAbs raised to BoNT and other toxins during T cell dependent immune responses, indicating that the HC50/A immunogen was a suitable surrogate immunogen and is able to portray the native N terminus of the toxin.(38)
The sero-specificity and endpoint ELISA titers were also determined for whole BoNTs. No cross-reactivity with BoNT/B or BoNT/E was observed, and F90G5-3 had an anti-BoNT/A titer of 3.1 ng/mL in whole BoNT/A toxin ELISA (Fig. 4A). The functional activity of MAb F90G5-3 was assessed using the mouse protection assay (MPA). Mice were challenged with 5LD50 of BoNT/A pre-incubated with MAb F90G5-3. Although MAb F90G5-3 was not able to neutralize all toxin activity, as death was observed in the treated mice, F90G5-3 treatment resulted in a slightly increased time to death (p<0.005, Fig. 4B).

Reactivity and functional activity of MAb F90G5-3 with whole BoNT. (
Discussion
We used recombinant HC50/A to develop a MAb to BoNT/A that binds a linear epitope in the HCN25/A subdomain (985WTLQDTQEIKQRVVF999) with nanomolar affinity. Moreover, the use of non-toxic HC50 antigens for the development of MAbs against whole BoNTs provides a safe alternative for the development of BoNT detection reagents. Within this study, only one MAb out of eight was highly reactive with whole BoNT/A and thus selected for further study.
The fact that we were able to develop antibody to this epitope utilizing the HC50/A receptor-binding domain and selecting for those that cross-react with whole BoNT/A is not surprising, as the epitope is surface exposed within the full-length BoNT/A (Fig. 3C). One can observe slight reactivity in the Pepscan (Fig. 3A) with a second peptide corresponding to amino acids 1195TNASQAGVEKILSAL1209. Given that F90G5-3 is a monoclonal antibody, this reactivity is most likely due to cross-reactivity with similar amino acid residues and thus an artefact of the in vitro pepscan assay, as this sequence is located within HC25/A sub-region of BoNT/A with which F90G5-3 does not specifically react (Fig. 2). Interestingly, comparison of the primary amino acid sequence in the region corresponding to the 985WTLQDTQEIKQRVVF999 epitope in BoNT/A to BoNT/B and BoNT/E reveals that five of 15 residues are identical between all three toxins, whereas paired comparison of BoNT/A with BoNT/B or BoNT/E shows only two of 15 and one of 15 are conserved and semi-conserved between all three BoNTs, respectively (Fig. 3B). Indeed, only six residues are unique within this region in BoNT/A, indicating that BoNT/A residues TQE-K-R are critical for F90G5-3 binding and strongly suggesting that this region is under intense immune or environmental selection pressure.
The modest neutralization of BoNT/A by MAb F90G5-3 demonstrates that interaction with epitope 985WTLQDTQEIKQRVVF999 only slightly decreases the functional activity of the neurotoxin. This partially neutralizing epitope lies within the HCN25/A subdomain of HC50/A, a region with greater primary sequence homology in comparison to the HCC25 subdomain, which is more antigenic and important for host cell ganglioside binding.(11,16,39) In the schematic representation of BoNT/A, one can observe the residues important for host cell ganglioside receptor binding are located on an adjacent surface to the F90G5-3 epitope (Fig. 3C, orange space fill). The interaction with the host cell gangliosides is the first low affinity binding interaction that brings the toxin in close proximity to the host cells in the dual receptor model of binding.(15) Antibody bound to the F90G5-3 epitope (Fig. 3C, red space fill) may cause steric hindrance to partially occlude the ganglioside-binding site and thus provide an explanation for the delayed time to death observed within this study. While the amino acid residues that interact with the host cell protein receptor have yet to be clearly elucidated, Garcia-Rodriguez and colleagues have recently co-crystallized neutralizing antibody fragments (CR1 and AR2) with BoNT/A, thus identifying critical contact residues for neutralization.(40) The amino acid residues of BoNT/A identified as the structural binding epitope of neutralizing CR1 are depicted in the pink color space fill, and the residues identified as part of the functional epitopes of both neutralizing MAbs CR1 and AR2 are depicted in the cyan space fill (Fig. 3C).(40) In this schematic, it does not appear that the epitope bound by MAb F90G5-3 interferes with these key neutralizing residues.
Most significantly, MAb F90G5-3 was specific for HC50/A and bound to a linear epitope (985WTLQDTQEIKQRVVF999) within the HCN25/A subdomain with nanomolar affinity. A search of the Immune Epitope Database (
Footnotes
Acknowledgments
This work was supported by the Public Health Agency of Canada, project CRTI 03-0021TD from the Chemical, Biological, Radiological, Nuclear and Expositive (CBRNE) Research and Technology Initiative (J.D.B.) and a National Institute of Allergy and Infectious Diseases contract (N01-AI-30050 to L.L.S.). Thanks are extended to M. Garbutt, the staff of the Monoclonal Antibody Section, and Emerging Bacterial Pathogens Division at the National Microbiology Laboratory for their expert technical help.
Author Disclosure Statement
The views and opinions expressed herein are those of the authors only, and do not necessarily represent the views and opinions of the Public Health Agency of Canada, the Government of Canada, or the CRTI.
The authors have no financial interests to disclose.