
Abstract
Histidine-rich protein-2 (HRPII) secreted by Plasmodium falciparum finds its use as a compelling marker in malaria diagnosis and follow-up. Monoclonal antibodies (MAbs) against P. falciparum HRPII are widely used in antibody-based diagnostic systems to detect HRPII protein in blood of malaria-suspected individuals. In this study, a set of five monoclonal antibodies against recombinant HRPII (rHRPII) were generated and assessed for their potential in diagnostics. Three among the five generated MAbs were of IgG1 isotype and the remaining were of IgM isotype. Probing the MAbs against proved P. falciparum infected serum and pooled control sera by immunoblotting revealed that the MAbs were successful in exposing malarial infection. Collectively, the generated MAbs have the potential to be used in immuno-based diagnostic systems uncovering P. falciparum infections.
Introduction
In P. falciparum infected individuals, the circulating HRPII is opted as a diagnostic target by various antibody-based tests, importantly, immunochromatographic technology (ICT) based rapid malaria diagnostic tests (MRDT), which are commercially available.(7–9) These MRDTs make use of monoclonal antibodies (MAbs) to sense HRPII in suspected sera and offer a great potential for diagnosis and follow-up of malaria, especially in remote areas and developing nations. When compared with traditional microscopy, these field-based tests enjoy good specificity and an uneven sensitivity, especially at low levels of parasitemia (<500 P/P/μL).(10–15) The World Health Organization (WHO) recommends a lower detection limit of 100 parasites/μL of blood for any non-microscopic rapid diagnostic test. This is similar to the level of sensitivity achieved by a well-trained, experienced microscopist.(16) The development of such capability presents a challenge, especially when the targeted HRPII protein sequence varies between the strains due to genetic heterogenicity.(17,18) One approach to accomplish the WHO recommendations is to screen an array of monoclonal antibodies, targeting a broader range of the antigen epitopes from widespread isolates.(19) Consequently, it has been a prerequisite to raise MAbs against HRPII from strains spanned across the globe.
A collection of monoclonal antibodies have been generated against HRPII for different purposes(4,20) although no reports of MAbs raised against the HRPII from Indian isolate have been documented. The aim of this study is to generate monoclonal antibodies against HRPII antigen from an Indian isolate of P. falciparum and to authenticate their worthiness for diagnosis.
Materials and Methods
All chemicals, reagents, and hybridoma media were procured from Sigma-Aldrich (Steinheim, Germany).
Cultivation of Plasmodium falciparum
An Indian isolate of P. falciparum from Malaria Research Centre (MRC, New Delhi) was used as a source of hrpII gene in this study. P. falciparum was cultured in A+ve human erythrocytes supported in RPMI media with additives (10% AB+ve human sera, 0.2% glucose, 0.1% sodium-bicarbonate, and 40 μg/mL of gentamycin), as described previously by Trager and Jenson.(21)
Cloning and expression of HRPII gene fragment
DNA was extracted from cultured P. falciparum by processing the infected RBCs with QIAamp DNA blood mini kit (Qiagen, Hilden, Germany) as per the manufacturer's instructions. Primers were designed and a portion of exon2 region of hrpII gene was amplified by PCR. The resultant PCR fragment was cloned in pQE-UA cloning and expression vector (Qiagen) in frame with the N-terminal histidine tag (His-tag), as per the manufacturer's instructions. The cloned gene was transferred into Escherichia coli M15 cells, and the successful transformants were induced to express rHRPII using 1 mM isopropyl-β-D-thio-galacto-pyranoside (IPTG). The cells harboring the recombinant protein were harvested and analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophorosis (SDS-PAGE) to confirm the protein expression.(22)
Purification of recombinant protein
Purification of rHRPII was carried out by affinity chromatography in native conditions using commercially available Ni-NTA columns (Qiagen) as per the manufacturer's instructions. Briefly the harvested pellet was lysed using ultrasound waves and the soluble recombinant protein was captured onto the Ni2+ ions in Ni-NTA resin. After a brief washing with 40 mM imidazole, the protein of interest was eluted using 350 mM imidazole. The purity of the rHRPII was examined for any contaminating proteins by SDS-PAGE analysis. Furthermore, to confirm the functionality of rHRPII, the protein was checked with commercially available MRDT system (NOW Malaria, Binax, Scarborough, Maine), which detects HRPII protein.
Immunization and generation of MAbs
A set of five female BALB/c mice weighing ∼20 g were immunized each with 50 μg of purified rHRPII emulsified in complete Freund's adjuvant through the subcutaneous route. Five booster doses with the same amount of immunogen emulsified with incomplete Freund's adjuvant were given at an interval of every 7 days through the intramuscular route. A final intraperitoneal injection of rHRPII (50 μg) was given to mice 3 days before fusion.
Cell fusion and culture
Fusion experiments were performed by slightly modifying the procedure elaborated by Kohler and Milstein.(23) Concisely, splenocytes harvested from the immunized mice were fused with Sp2/0-Ag14 myeloma cells using the fusing agent polyethylene glycol (PEG 2000). The fused cells were resuspended in selective HAT (hypoxanthine-aminopterin-thymidine) medium containing Dulbecco's modified Eagle's medium (DMEM) with 20% fetal calf serum and 1% selective supplement HAT. The resuspension was distributed in 96-well culture plates and incubated at 37°C with 5% CO2. On day 4, half the portion of media was replaced with fresh HAT medium and on day 7, HAT was entirely replaced with hypoxanthine-thymidine medium (HT).
Hybridoma screening and propagation
Hybridomas were screened for any MAbs produced by ELISA. Briefly, rHRPII at a concentration of 1 μg/100 μL/well in 50 mM carbonate buffer (pH 9.6) was coated on to a microtiter plate, and the culture soup was scrutinized for MAbs by standard ELISA procedures. The ELISA was developed with 0.4% OPD and 2 mM H2O in citrate phosphate buffer (pH 5.0) and the developed color was read at 490 nm. Selected hybridoma cell lines producing MAbs were expanded and grown on HT medium in 150 mL tissue culture flasks to collect the culture soup after 2 days.
Isotyping of murine immunoglobulins was performed by sandwich ELISA employing class- and subclass-specific anti-mouse immunoglobulins using mouse-type sub-isotyping kit (Bio-Rad, Hercules, CA), following the manufacturer's instructions.
Scrutiny of MAbs
The generated MAbs were tested for their response towards rHRPII by immunoblot reactions. Further the MAbs were investigated for their diagnostic usefulness by determining their reactivity with microscopy confirmed P. falciparum infected serum and pooled normal human sera (negative control). Briefly, the sera to be tested were diluted to the ratio of 1:1 with PBS and transferred onto the PVDF membrane after SDS-PAGE and Western blot analysis.(24) The blotted membrane was directly probed with culture soup containing MAbs from the hybridoma clones and developed with horse radish peroxidase (HRP)-DAB enzyme-substrate system using HRP-conjugated goat anti-mouse antibodies (DAKO, Glostrup, Demark) for revealing the attachment between the antigen and MAbs.
Results
Cloning and expression of rHRPII protein
PCR generated an amplicon size of approximately 900 bp, which was cloned into pQE-UA vector. The cloned amplicon expressed as a fusion protein with His-tag at its amino acid terminal was successfully affinity-purified using Ni-NTA column. The purified rHRPII relatively migrated along with the approximately 62 kDa protein standard and was found to be free from significant amounts of E. coli host proteins when examined visibly by SDS-PAGE (Fig. 1). The purified rHRPII gave a positive response with commercially available MRDT, establishing its validity (Fig. 2).
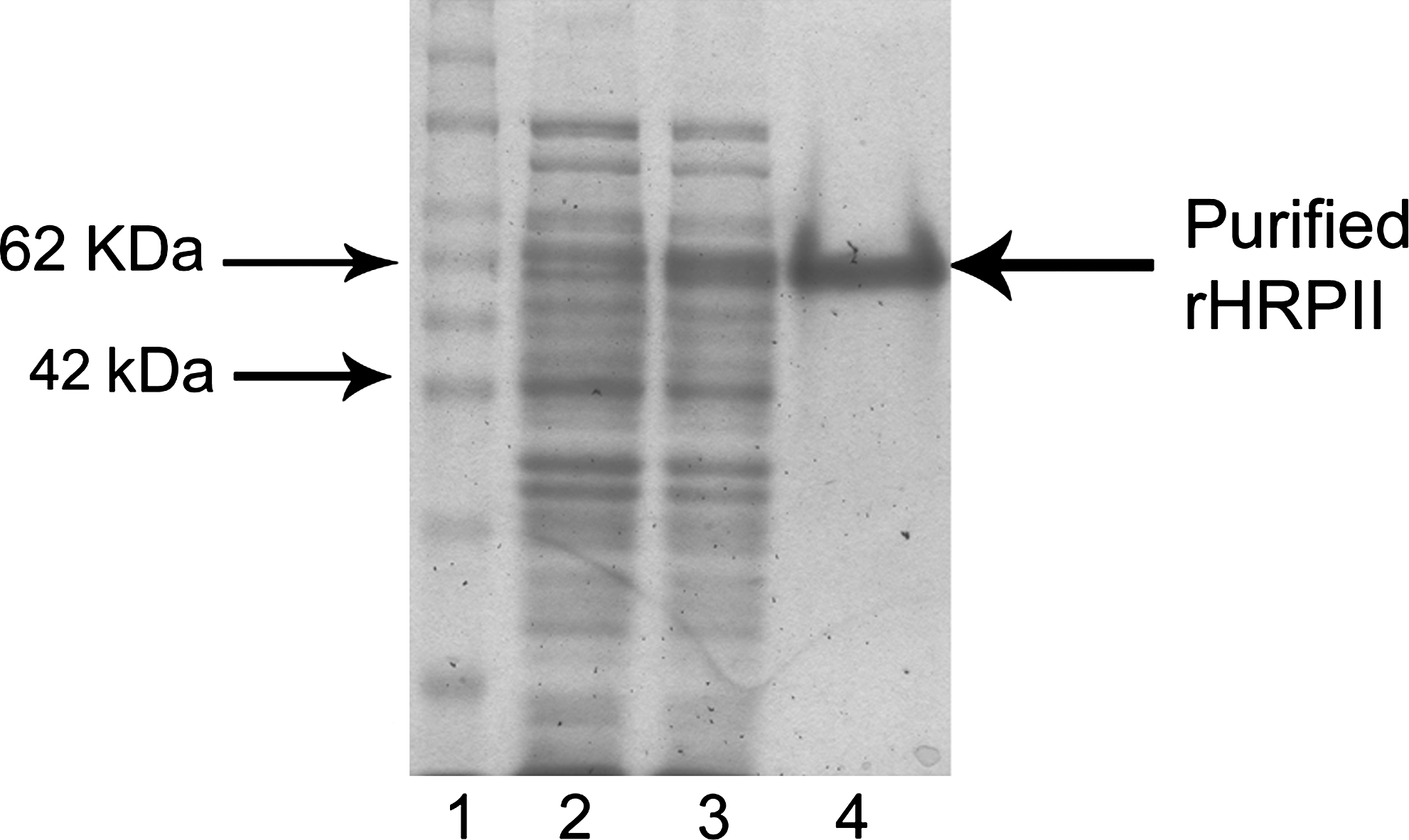
Purification of recombinant HRPII. Lane 1, protein marker; lane 2, uninduced clone; lane 3, induced clone; lane 4, purified rHRPII.
Reactivity of purified rHRPII with MRDT system. This system detects HRP of P. falciparum and lactate dehydrogenase (LDH) of other Plasmodium sp.
Generation and characterization of murine MAbs
The fusion experiments generated five clones and the generated MAbs were renamed MHRP1 to MHRP5. Three of the MAbs (MHRP1 to MHRP3) were of IgG1 isotype while the other two were of IgM isotype. When the generated MAbs were investigated for their reactivity against rHRPII, all the MAbs reacted with rHRPII antigen in immunoblot reactions (Fig. 3, Table 1).

Reactivity of generated MAbs with purified rHRPII. Lane 1, protein marker; lane 2, MHRP1; lane 3, MHRP2; lane 4, MHRP3; lane 5, MHRP4; lane 6, MHRP5.
Reactivity with human sera
The intention in raising MAbs is to exercise their potency for diagnostic purposes. When examining them for their worthiness with the available P. falciparum infected serum sample by immunoblotting, all the MAbs clearly identified a protein of approximately 62 kDa, which corresponds with the expected size of secreted HRPII. Another protein with an approximate MW of 50 kDa was also identified by the MAbs (Fig. 4). In the control immunoblot, none of the MAbs recognized a protein from normal human sera (Fig. 5, Table 1).

Reactivity of generated MAbs with pooled P. falciparum infected sera. Lane 1, protein marker; lane 2, MHRP1; lane 3, MHRP2; lane 4, MHRP3; lane 5, MHRP4; lane 6, MHRP5.

Reactivity of generated MAbs with pooled P. falciparum infected sera. Lane 1, protein marker; lane 2; MHRP1; lane 3, MHRP2; lane 4, MHRP3; lane 5, MHRP4; lane 6, MHRP5.
Discussion
Malaria caused by P. falciparum is a serious disease in developing and third world countries. The pathogen secretes a series of histidine-rich proteins, of which the parasite encoded water-soluble HRPII is secreted out by the infected RBCs as an extracellular antigen. Recent studies have established the genetic polymorphism in P. falciparum hrpII gene, resulting in the heterogenicity of HRPII protein across the strains. Hence the available epitopes recognized by MAbs vary significantly among the strains, leading to sensitivity issues in tests employing MAbs.(25,26)
In the present study we generated a set of five monoclonal antibodies against the recombinant form of HRPII from an Indian isolate. We examined the reactivity of the MAbs against the rHRPII and P. falciparum infected serum to establish their validity and worthiness, respectively.
As the hrpII gene is heterogenic between isolates, primers were designed to bind with the conserved regions of the gene, and a portion of the exon2 region encoding the histidine-rich sequences were amplified. Recombinant HRPII protein constructed against these histidine-rich sequences was affinity-purified in native conditions and the developed product was authenticated by its reactivity in MRDT.
To ensure that the MAbs are generated against the epitopes of rHRPII, immunoblot reactions were performed against the recombinant protein. All the MAbs identified the rHRPII with the same amount of strength, establishing the affinity towards the target epitopes.
To establish the worthiness of the generated MAbs for diagnostic purpose, we probed them against a microscopically confirmed P. falciparum infected human sera transblotted onto the PVDF membrane. All the generated MAbs detected an ∼62 kDa protein in the P. falciparum infected serum with varying strength. The 62 kDa protein is considered to be the secreted HRPII, based on the predicted molecular weight. An advantage with immunoblot is that the cross-reactivity exhibited by the antibody, if any, over non-specific proteins can be determined. Here, an ∼50 kDa protein was also recognized in P. falciparum infected sera. This is due to the presence of more or less similar epitopes shared between the HRPII and the lower MW protein.
Parra and colleagues(27) identified a 72 kDa protein of human origin when probing P. falciparum infected human plasma with anti-HRPII MAbs. This exposes the epitope sharing between the human serum proteins and HRPII, which emerge as cross-reactivity in immunoblot reactions with MAbs. To rule out the lower MW protein as a non-specific interfering protein of human origin, we probed the generated MAbs against pooled sera from normal human individuals. The generated MAbs did not detect any protein from human sera. This not only highlights the specificity exhibited by the MAbs towards the epitopes of HRPII but also emphasizes that the lower molecular weight protein is not of human origin—here it may be from P. falciparum.
P. falciparum infected RBCs in humans secrete a multiplet of HRPII species with MW spanning across different kilo Dalton, which are visualized as a “ladder” in immunoblots.(27) Our immunoblot reactions with positive serum did not produce the multiplet bands, yet the absence of any cross-reactivity with the serum proteins support our view that the protein is not of human origin. The second likely explanation for the low MW second protein is histidine-rich protein-3 (P. falciparum-HRPIII). HRPIII protein is a P. falciparum specific secretary protein that shares epitopes with P. falciparum HRPII.(28,29) Monoclonal antibodies against HRPII were found to cross-react with HRPIII and this contributes to the detection of HRPIII in immunoassays targeting HRPII. This criss-cross response of antibodies sum up the sensitivity by detecting HRPIII along with HRPII protein.(25) These two possibilities further stress that our MAbs had recognized the epitopes of P. falciparum origin related to the HRPII protein. The identification of 62 kDa HRPII and a lower MW protein of P. falciparum origin along with the absence of cross-reactivity with human serum protein confirm that the generated MAbs successfully differentiated P. falciparum infected serum from normal human serum, proving their diagnostic potential.
Eradication of this fatal version of malaria is met with barricades, predominantly due to a vague knowledge of the parasites' life inside the host and inept diagnostic tools. This limitation creates a snag in developing better drugs, vaccines, and diagnostics. Antibodies against HRPII of P. falciparum find their use in decoding the various colossal domains—from exploration to diagnostics in the battle against malaria.(30,31) The MAbs generated in this study were proved to be worthy for the diagnosis of P. falciparum infection and can be used to improve the existing monoclonal antibody based immunoassays, especially MRDTs.
Footnotes
Acknowledgments
The authors are thankful to Dr. Vijayaraghavan (Director, DRDE, Gwalior), Dr. M.P. Kaushik (DRDE, Gwalior), and Dr. C.R. Pillai (National Institute of Malaria Research, New Delhi) for their support in this study.
Author Disclosure Statement
The authors have no financial interests to disclose.