
Abstract
SURF-6 is an evolutionarily conserved nucleolar protein that is important for cell viability; however, its function in mammals still remains uncertain. The aim of this study is to generate monoclonal antibodies to human SURF-6 protein suitable for fundamental and biomedical research. The full-size human SURF-6 was expressed as a recombinant GST-fusion protein and used as an antigen to generate monoclonal antibodies, S79 and S148, specific for SURF-6. The monoclonal antibody produced by hybridoma clone S79 specifically recognizes endogenous SURF-6 by Western and immunofluorescence analyses in various cultured human cells, and by immunohistochemistry in paraffin-embedded sections of human breast cancer samples. Moreover, S79 immunoprecipitates protein complexes containing SURF-6 from HeLa cells extracts. The antibody S79 recognizes SURF-6 only in human cells; however, the antibody produced by hybridoma clone S148 can detect SURF-6 of human and mouse origin. Monoclonal antibodies to the nucleolar protein SURF-6 described in this work can be a useful tool for studies of ribosome biogenesis in normal and cancer cells.
Introduction
The nucleolar protein SURF-6 (361 amino acid residues in humans) is important for mammalian cell viability.(7) SURF-6 has a unique evolutionary conserved domain at its carboxy terminus that constitutes a novel family of eukaryotic proteins extending from human to yeast.(8,9) The Saccharomyces cerevisiae homolog of SURF-6, Rrp14/yk1082c, is a multifunctional protein, which is involved in synthesis of 35S pre-rRNA, assembly of the large ribosomal subunit, and regulation of the cell polarity.(10,11) Mouse SURF-6 has high nucleic acid binding capacities both in vitro and in situ, suggesting that mammalian SURF-6 may have functions analogous to its yeast counterpart in pre-rRNA processing.(12–15)
Recent genomic and proteomic global functional screens revealed that the SURF-6 gene is overexpressed in some types of human cancer cells and can be catalogued as a potential gene related to cancer development and progression.(16–18) In accordance with these in silico data, recently obtained results indicate that there is a higher level of SURF-6 expression in leukocytes of leukemia patients.(19) Moreover, large scale profiles of RNAi-induced-loss-of-function phenotypes reveal that depletion of SURF-6 in HeLa cells augments the number of binuclear cells.(20) These observations suggest that SURF-6 may be involved in the regulation of cell proliferation and strengthen an idea on a particular role of SURF-6 in human cancer cells.
The major aim of this work is to raise mouse monoclonal antibodies suitable for studies of SURF-6 in normal and cancer cells of human origin. Such antibodies should allow the identification of SURF-6 in human samples by Western, immunocytochemical, and immunohistochemical analyses.
Material and Methods
Cell cultures
Mouse NIH/3T3 and human HeLa, CCRF-SB, NCI-H460, U-87 MG, and K-562 cells were purchased from the Russian Collection of Cell Cultures (Institute of Cytology, Russian Academy of Sciences, St. Petersburg, Russia). NIH/3T3, HeLa, CCRF-SB, NCI-H460, U-87 MG, and K-562 cells were grown in DMEM or RPMI 1640 medium (PanEco, Moscow, Russia) according to instructions provided by the supplier with 10% fetal calf serum supplement (HyClone, Waltham, MA), 2 mM L-glutamine, penicillin, and streptomycin (250 U/mL each) at 37°C in the atmosphere of 5% CO2 and 95% O2.
Expression of human SURF-6 fused to GST
Production of the recombinant protein GST-SURF-6 has been described in detail previously.(21) Briefly, cDNA encoding human SURF-6 was amplified and cloned into the gluthatione-S-transferase (GST) fusion expression vector, pGEX-2T (Amersham Pharmacia Biotech, Little Chalfont, United Kingdom) via BamHI and EcoRI restriction sites. The GST-tagged protein was expressed in Escherichia coli strain BL21-Codon-Plus (Stratagene, Valencia, CA) and purified using gluthatione-Sepharose 4B beads (Amersham Pharmacia Biotech) yielding amounts sufficient for mouse immunization.
Monoclonal antibody production
Three BALb/c female mice were subcutaneously injected with 50 μg GST-SURF-6 fusion dissolved in 0.5 mL Freund's complete adjuvant. The second and third immunizations were administered in 0.5 mL Freund's incomplete adjuvant after 7 and 14 days, respectively. Serological responses to the fusion protein were monitored by ELISA, immunoblots, and immunofluorescence in HeLa cells. Three days after the final boost the sensitized animals were sacrificed and spleens were removed. Splenocytes were fused with mouse myeloma P3X63-Ag8.653 cells with 50% polyethylene glycol 1450 and cultured in RPMI 1640 medium containing 10% fetal calf serum (FCS), hypoxanthine, and azaserine to select hybrid clones.(23) Approximately 100 clones were obtained and those that produced antibodies to SURF-6 were selected by screening each clone culturing medium by ELISA, immunocytochemistry, and immunoblots using HeLa and NIH/3T3 cells. Two selected clones, S79 and S148, were established by limiting dilutions following a standard protocol.(22)
Antibodies precipitated with 50% ammonium sulfate dialyzed against binding buffer (10 mM sodium phosphate, pH 8.0) were loaded on a column with DEAE matrix. The antibody was extensively washed with 10 volumes of column bed with binding buffer and eluted with NaCl gradient of concentrations ranging from 50 to 250 mM. The eluted fractions of an antibody were monitored by spectroscopy in 280 nm wavelength (Biologic LP Chromatography system, Bio-Rad, Hercules, CA).
Western blot analysis
5×106 cells were lysed in 200 μL buffer containing 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 10% glycerol, 0.5% Triton X-100, and protease inhibitor cocktail for general use (Sigma, St. Louis, MO) on ice. Samples were diluted in standard 5× Laemmli buffer, 50 μg of total protein was loaded per lane, run on 12% SDS-PAGE, and transferred to a nitrocellulose membrane (Millipore, Billerica, MA). Membrane was blocked in 5% non-fat dry milk in TBST buffer (20 mM Tris-HCl [pH 7.6], 0.05% Tween-20) and incubated with an anti-SURF-6 mouse antibody (1:1000) or the rabbit polyclonal SURF-6 antibodies (1:5000),(13,14) and then with anti-mouse IgG or anti-rabbit IgG conjugated with horseradish peroxidase (1:20,000, Sigma). Blots were developed using an ECL+Plus chemiluminescence kit (Amersham Pharmacia Biotech).
Immunocytochemistry
Cells were fixed with 3% paraformaldehyde in standard phosphate-buffered saline (PBS) for 20 min at room temperature, treated with 0.2% Triton X-100 in PBS for 10 min, and incubated with a SURF-6 antibody diluted 1:100–1:500 in PBS. After washing in PBS, cells were incubated with anti-mouse IgG conjugated with Texas red (Jackson ImmunoResearch Laboratories, West Grove, PA) for 30 min, stained with 0.1 μg/mL 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI, Molecular Probes, Carlsbad, CA), and mounted into Mowiol. Specimens were examined under an epifluorescence microscope Axiovert 200 (Carl Zeiss, Hamburg, Germany) using objectives 40×/0.75 PlanNeofluar and 100×/1.25 Fluar. The images were captured by a 12-bit monochromatic digital camera CoolSnap cf (Roper Scientific, Tucson, AZ) and treated using Adobe Photoshop (v 9.0, San Jose, CA).
Immunohistochemistry
Human breast cancer samples were obtained during the course of surgical operations of patients (Gertsen Research Institute of Oncology, Moscow, Russia), fixed in 10% formalin embedded in paraffin, and cut on sections following a standard procedure. To retrieve antigens, sections were incubated in an Autostainer plus reagent (Dako, Carpinteria, CA) in 10 mM citrate buffer (pH 6.0, Dako) and washed in TBS (20 mM Tris-HCl [pH 7.6]). Sections were immersed in TBS containing 3% H2O2 for 5 min to inactivate endogenous peroxidase and incubated in TBS supplemented with 2% BSA (Sigma) for 10 min and finally with the S79 antibody (diluted 1:50–1:500 in TBS) at 4°C overnight. Sections were washed in TBS four times. An UltraVision LP Large Volume Detection System kit (Thermo Scientific, Waltham, MA) was used for localization of SURF-6. Sections were studied under a Leica DMRB microscope (Hamburg, Germany) coupled to a digital camera (U-CMAD3, Olympus, Tokyo, Japan) with magnifications of 200×/400×.
Immunoprecipitation
6×106 HeLa cells were resuspended in 8 mL of ice-cold buffer A (10 mM Tris-HCl [pH 7.4], 10 mM NaCl, 2 mM MgCl2, 0.05% Igepal CA-630, 0.5 mM DTT, 1 mM PMSF) containing complex protease inhibitor cocktail (Sigma) and homogenized by passing through a 0.4 mm syringe needle five times. Released nuclei were pelleted by centrifugation at 700 g for 10 min at 4°C, resuspended in 0.5 mL of ice-cold buffer B (50 mM Tris-HCl [pH 7.4], 10 mM NaCl, 2 mM EDTA, 0.3% Igepal CA-630, 10% glycerol, 100 μg/mL RNAse A, 0.5 mM DTT, 1 mM PMSF, and protease inhibitor cocktail) and incubated on a rocker for 30 min at 4°C. NaCl was added to the final concentration of 0.4 M, and the sample was incubated on a rocker for 30 min at 4°C. Cellular debris was removed by centrifugation at 16000 g for 10 min at 4°C. Extract was diluted in buffer C (50 mM Tris-HCl [pH 7.4], 0.5 mM DTT, and protease inhibitors) to the final concentration of NaCl (135 mM). MgCl2 and DNase I (Sigma) were then added to the final concentration of 2 mM and 50 U/mL, respectively.
Nuclear extracts were incubated with 10 μg of the antibody S79 for 2 h, and then with 25 μL of protein G-sepharose (Sigma) for 1 h at 4°C on a rocker machine. G-sepharose beads were pelleted by centrifugation, washed with buffer (20 mM Tris-HCl [pH 7.4], 130 mM NaCl, 2 mM EDTA, 0.5% Igepal CA-630, 0.5 mM DTT, 1 mM PMSF), resuspended in standard 5× Laemmli buffer, and subjected to immunoblots with the polyclonal rabbit SURF-6 antibodies.(13,14)
Results and Discussion
Production of mouse monoclonal antibodies specific to human SURF-6 and their reactivity on Western blots
Antibodies against proteins required for ribosomal biogenesis are commonly used for studying a protein role in ribosome assembly and related processes.(23) To further examine a functional significance of a nucleolar protein SURF-6 in ribosomal biogenesis, we generated a non-denatured recombinant human SURF-6 protein fused with GST to raise original monoclonal antibodies to the protein. We used a GST-SURF-6 protein rather than a recombinant His-tagged SURF-6 protein because the SURF-6-GST recombinant protein can be produced and purified from bacteria under non-denaturing conditions. In addition, the isoelectric point of SURF-6 (pI 10.65) limits its yield from Ni-NTA agarose columns, which are commonly used for the purification of His-tagged proteins, due to its strong ability to bind the matrix of nickel-based columns. As opposed to the recombinant SURF-6 protein, the highly basic SURF-6 protein has a reduced non-specific binding to L-glutathione columns used to purify GST-targeted proteins. In this study we describe for the first time two mouse hybridoma clones that produce antibodies to human (antibodies S79 and S148) and mouse (antibody S148) SURF-6 protein. Both antibodies recognize SURF-6 on immunoblots in total lysates of HeLa cells with the characteristic band of 46 kDa (Fig. 1). However, only S148 recognizes SURF-6 in lysates of mouse NIH/3T3 cells, in which SURF-6 has an apparent electrophoretic mobility of 43 kDa. The raised antibodies were also capable of recognizing SURF-6 in human cells of various origin cultured in vitro, including acute lymphoblastic leukemia CCRF-SB cells, lung cancer NCI-H460 cells, glioblastoma U-87MG cells, and erythromyeloblastoid leukemia K562 cells (Fig. 2). As in HeLa cells, S79 and S148 detect one band of 46 kDa in lysates of U-87MG cells. In addition to the major band, both antibodies recognize a minor band of 42 kDa in CCRF-SB and NCI-H460 cells. Only S148 detects a similar minor band in K-562 cells (Fig. 2). The minor bands observed on immunoblots of CCRF-SB, NCI-H460, and K-562 cells may result from post-translational modifications of SURF-6. Such modifications may include SURF-6 phosphorylation.(24) Alternatively, the minor bands can be SURF-6 isoforms resulting from alternative splicing of a singular SURF-6 mRNA precursor. Further analysis of the expression and mobility of SURF-6 in different human cancer cells is required to assess the origin and biological significance of the additional bands revealed. Taken together, our results show that S79 and S148 recognize different epitopes in the SURF-6 molecule.
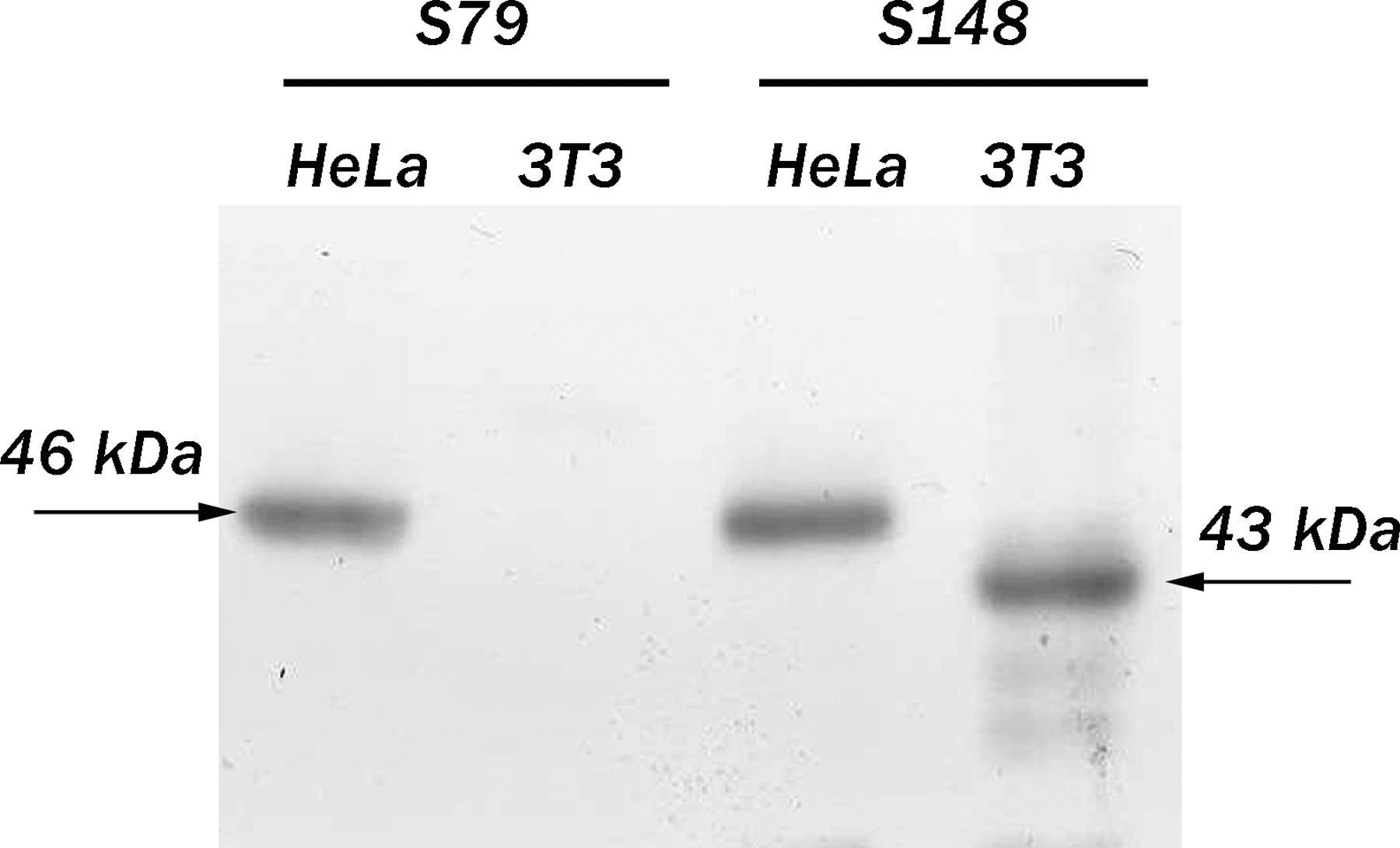
Western blot analyses of total lysates of human and mouse cells with S79 or S148 antibody. Apparent electrophoretic mobility of SURF-6 in human HeLa cells (46 kDa) and in mouse NIH/3T3 cells (43 kDa) are indicated by arrows.
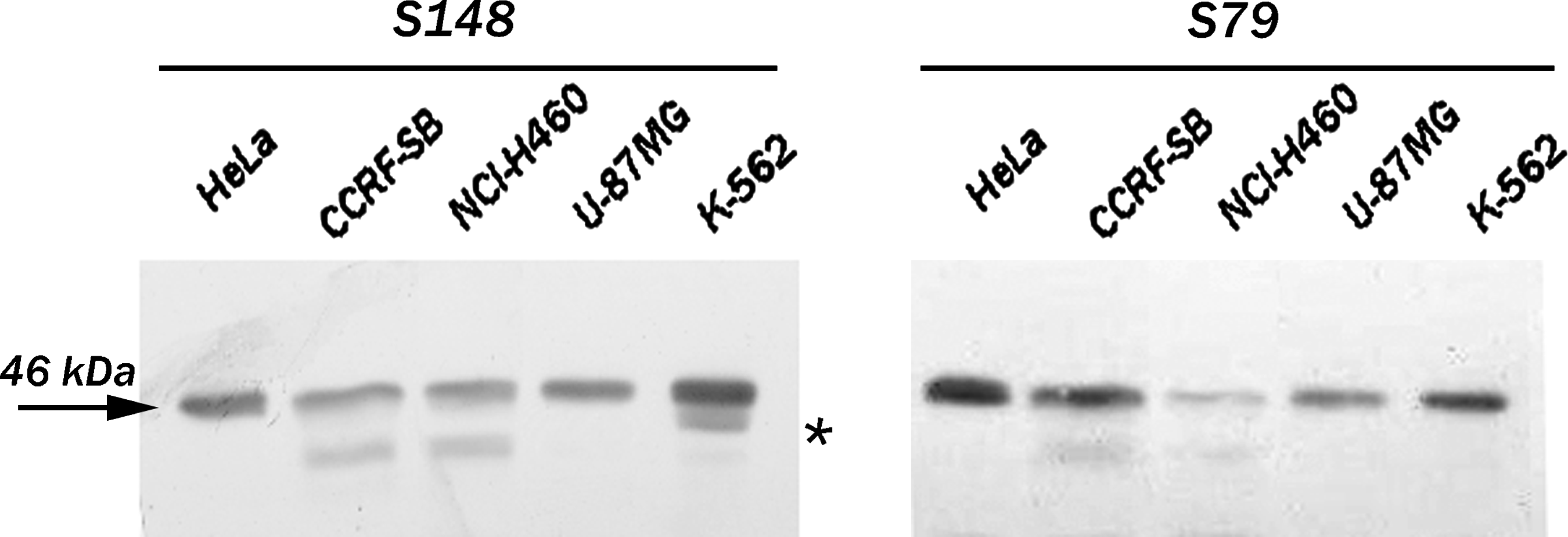
Western blot analyses of total lysates of different human cells with S79 or S148 antibody. Both antibodies recognize the major SURF-6 band (46 kDa, arrow) in HeLa, CCRF, NCI-H460, U-87 MG, and K-562 cells. Both antibodies also reveal a minor band of an apparent electrophoretic mobility of 42 kDa in CCRF-SB and NCl-H460 cells. In addition, S148 antibody also detects a band located below the major SURF-6 band in K562 cells (asterisk).
Immunocytochemical and immunohistochemical analyses of SURF-6 with S79 and S148 monoclonal antibodies
SURF-6 has been identified as a nucleolar protein using the rabbit polyclonal antibodies described earlier in mouse NIH/3T3 cells.(13,14) In the present study we show that the specific nucleolar localization of SURF-6 can be revealed with the monoclonal antibodies S79 and S148 in HeLa cells fixed with paraformaldehyde (Fig. 3) or acetone (data not shown). A weak extranucleolar staining may indicate a specific localization of SURF-6 protein in the nucleoplasm of human cells. The S148 antibody reveals SURF-6 in nucleoli of mouse NIH/3T3 cells similar to their labeling with the rabbit polyclonal antibodies (Fig. 4).(9,14)
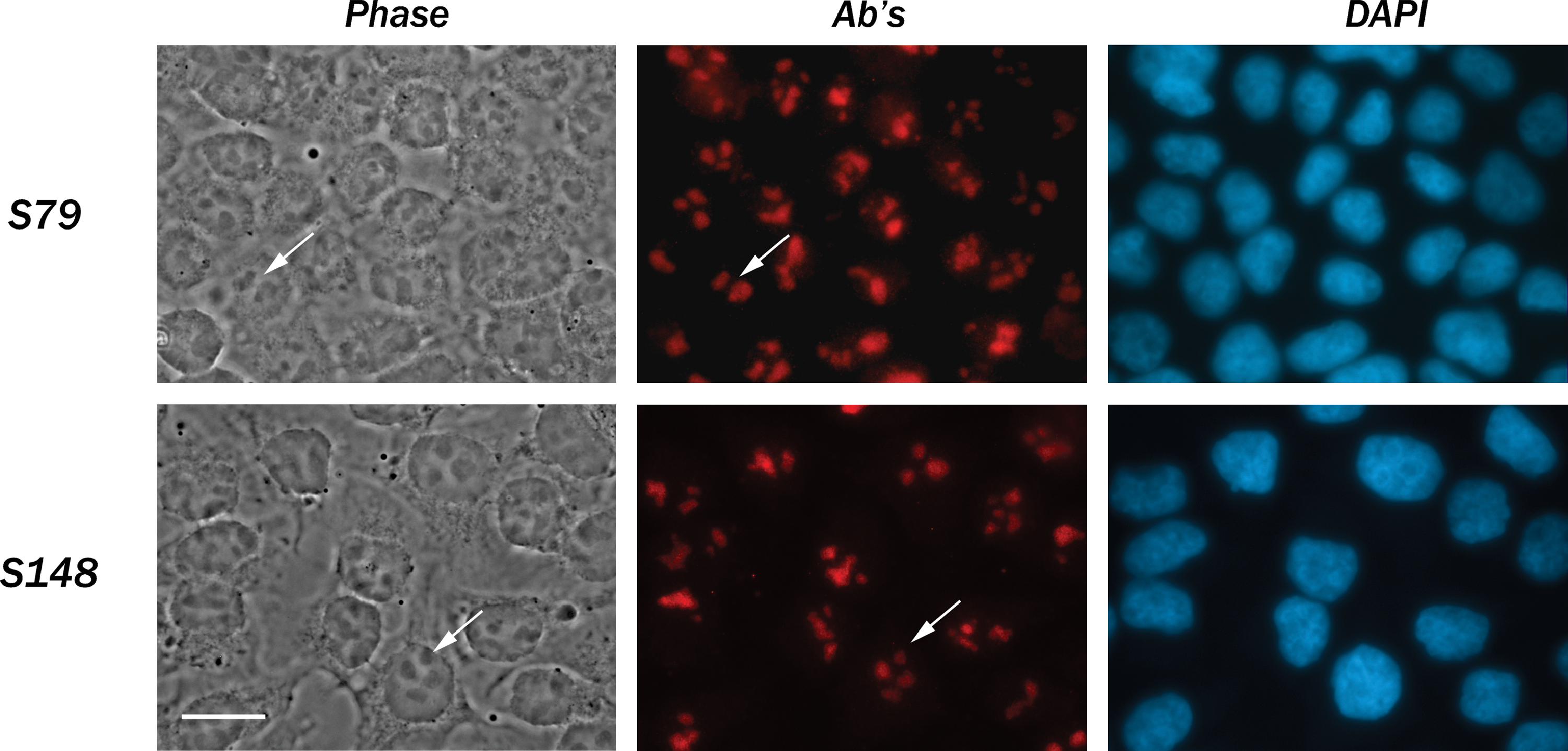
Immunofluorescence labeling of HeLa cells fixed with paraformaldehyde with the S79 or S148 antibody. Phase, phase contrast; Ab's, immunolabeling of cells with S79 or S148 antibody; DAPI, staining of nuclei with DAPI. Both antibodies label nucleoli (arrows). Bar, 10 μm.
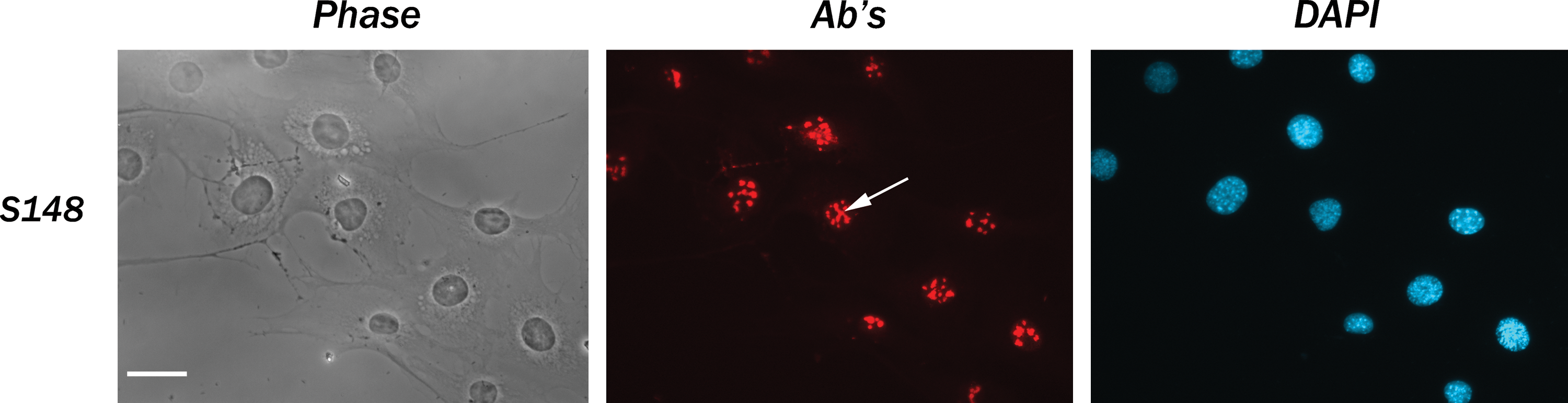
Immunofluorescence labeling of NIH/3T3 cells fixed with paraformaldehyde with the S148 antibody. Phase, phase contrast; Ab's, immunostaining with the S79 antibody; DAPI, staining of nuclei. The antibody labels nucleoli (arrow).
Immunoreactivity of the human-specific anti-SURF-6 antibody S79 was further examined in formalin-fixed and paraffin-embedded samples obtained from a patient with breast cancer. As seen in Figure 5, S79 recognizes SURF-6 in nucleoli on sections of breast cancer samples thus indicating that S79 is suitable for immunohistochemical research.
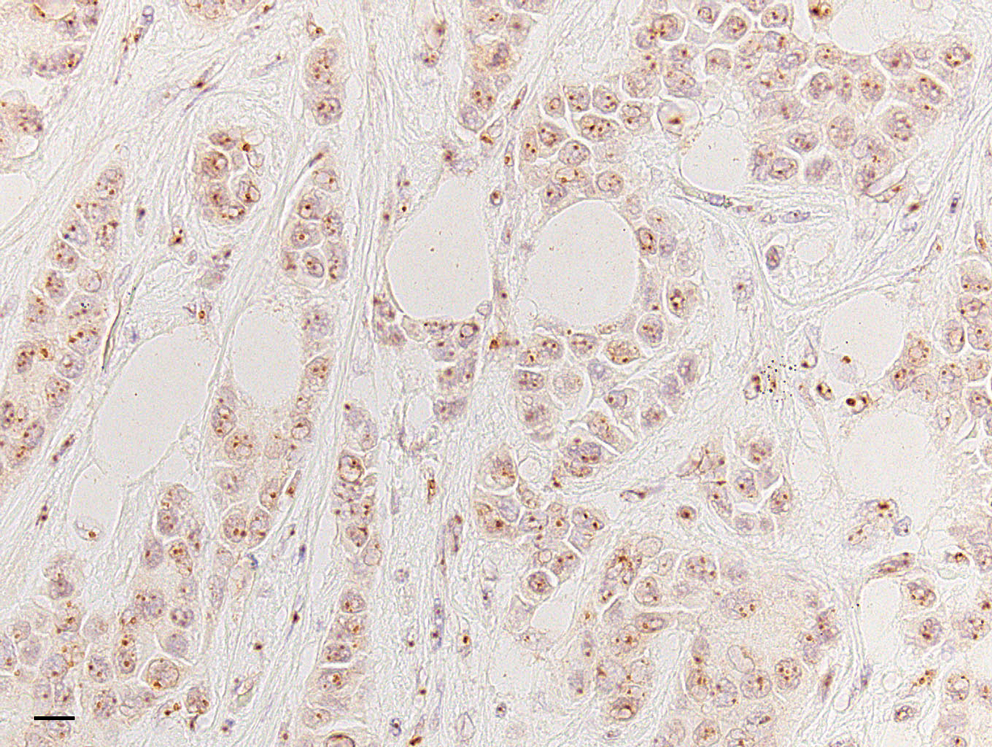
Immunohistochemical labeling of formalin-fixed and paraffin-embedded samples obtained from a patient with breast cancer with the S79 antibody. DAB was used as a substrate; the section was counterstained with hematoxylin to contrast nuclei. Nucleoli are intensely labeled. Bar, 10 μm.
Immunoprecipitation of protein complexes containing SURF-6 with monoclonal antibody S79
The antibody S79 was also tested for its capacity to immunoprecipitate SURF-6 from HeLa cell extracts enriched in nuclear and nucleolar proteins. DNase I and RNase A treatments of the extracts were applied because of strong nucleic acid binding capacities of SURF-6 in vitro and in situ.(12,14) Immunoprecipitated proteins were resolved on 12% SDS-PAGE (Fig. 6, lane IP). The gel staining with Coomassie Blue G-250 revealed bands corresponding to the heavy and light chains of mouse IgG (Fig. 6, asterisks) and several proteins including SURF-6 (Fig. 6, lanes IP and IB). These observations show that the S79 antibody can precipitate a complex of proteins, which are the most likely SURF-6 protein partners in HeLa cells. As known, the yeast homolog of SURF-6, Rrp14/yk1082c, interacts with a number of proteins that are required for ribosomal biogenesis (SSF1, EBP2, RRP1), control of the cell polarity, and cell cycle progression (Gic1, Gic2, CDC42, CKA2).(9,10) Based on these results, we concluded that the S79 antibody may be applied in immunoprecipitation experiments, which aim to identify SURF-6 interacting proteins in human cells.
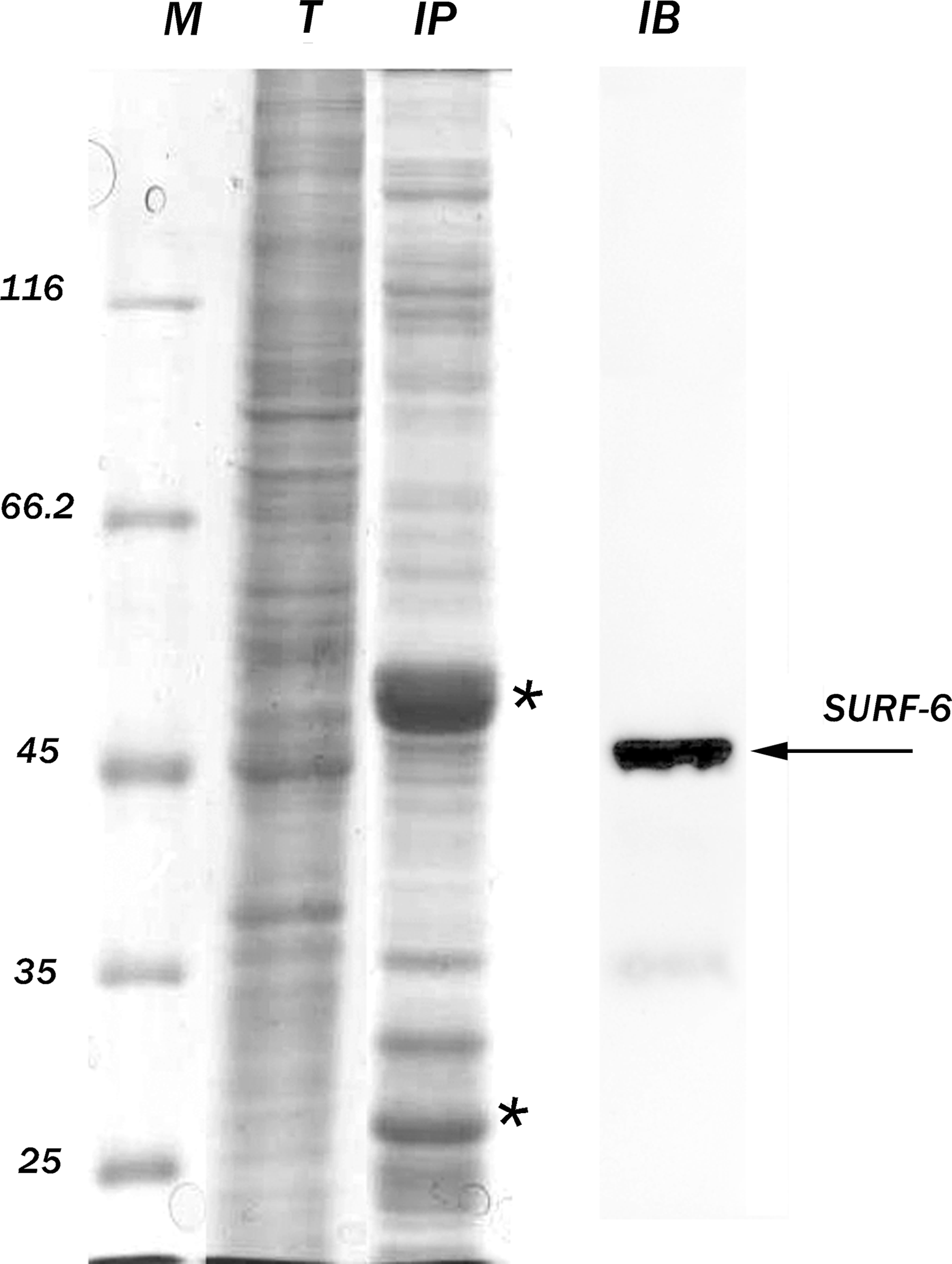
Immunoprecipitation of protein complexes from HeLa cell extracts with the S79 antibody. M, protein markers (kDa); T, SDS-PAGE of extracts enriched in nuclear proteins after staining with Coomassie blue; IP, precipitated proteins; IB, immunoblot of the precipitated proteins developed with the rabbit polyclonal SURF-6 antibody.(13) Asterisks indicate the heavy and light chains of mouse IgG. Arrow indicates the major band corresponding to SURF-6.
Footnotes
Acknowledgments
This study was supported by the Russian Ministry of Sciences and Education (grants 14.740.11.0116 and 14.740.11.0171). The authors are grateful to Dr. O. Zharskaya for help with the immunocytochemical experiments.
Author Disclosure Statement
The authors have no financial conflicts to disclose.