
Abstract
Programmed cell death 1 (PD-1) is an immunoregulatory receptor on T cells that binds two ligands, B7-H1 and B7-DC. Although accumulating reports suggest a critical role for the B7-H1:PD-1 pathway in peripheral tolerance, the actual involvement of B7-DC has not been well confirmed. Here, we established a new MAb against mouse B7-DC (MIH37) and compared its functional properties with a previously established anti-B7-DC MAb (TY25). Binding analyses using flow cytometry demonstrated that MIH37 showed an approximately four-fold higher binding affinity to B7-DC and stronger inhibitory effects on B7-DC:PD-1 binding. In contrast to the effects of TY25, treatment with MIH37 at both sensitization and challenge inhibited hapten-induced contact hypersensitivity reactions. Furthermore, the addition of MIH37 inhibited OVA-specific T cell responses in vitro. The inhibitory effects of MIH37 were counteracted by co-blockade with PD-1 and absent in PD-1-deficient mice, suggesting PD-1-dependent action of MIH37. Our present results suggest that greater complexities of PD-1-mediated functions are induced via ligand binding for controlling immunity and tolerance.
Introduction
Thus, the actual function of B7-DC and the contribution of exact receptor(s) have not been well clarified. In this study, we established a new anti-B7-DC MAb (MIH37) and examined its binding properties. We also evaluated the effects of in vivo administration in parallel comparative studies using our previous MAb, TY25. The results demonstrated that different MAbs with differential binding properties exert opposite functions during immune responses.
Materials and Methods
Mice
BALB/c and C57BL/6 (B6) mice were purchased from Japan SLC (Hamamatsu, Shizuoka, Japan). PD-1-deficient mice on a BALB/c background(31) were kindly provided by Dr. Tasuku Honjo through RIKEN RBC (Tsukuba, Ibaraki, Japan). All mice were maintained under specific pathogen-free conditions and used at 8–16 weeks of age. All procedures were reviewed and approved by the Animal Care and Use Committee of Tokyo Medical and Dental University.
Transfectants and Ig fusion proteins
T lymphoma L5178Y and plasmacytoma J558L were maintained in RPMI 1640 medium containing 10% FBS. B7-DC-transfected L5178Y (B7-DC/L5178Y) was established as previously described.(4) PD-1-transduced J558L cells (PD-1/J558L) were generated by retroviral transduction with PD-1/pMXs-IRES-GFP using the Plat-E system as described previously(32) and cells expressing high levels of PD-1 were sorted by flow cytometry. PD-1Ig and B7-DCIg fusion proteins, which consisted of the extracellular portion of mouse PD-1 and B7-DC and the hinge-CH2-CH3 domains of the human IgG1 heavy chain, respectively, were prepared as described previously.(33) The extracellular portions of B7-DC, B7-H1, and PD-1 were amplified from full length cDNA-containing plasmid B7-DC/pCAG, B7-H1/pMKITneo, and PD-1/pMKITneo, respectively, using the following primers: B7-DC, 5′-CTC GAG GCC ACC ATG CTG CTC CTG CTG CCG ATA C-3′ and 5′-AGA TCT CTG GGG ACT TTG GGT TCC ATC CGA-3′; B7-H1, 5′-CAG CTC GAG ACC ATG AGG ATA TTT GCT GGC-3′ and 5′-AGA GAT CTC TGT TCT GTG GAG GAT GTG T-3′; and PD-1, 5′-CCG CTC GAG ATG TGG GTC CGG CAG GTA-3′ and 5′-CGG GAT CCG CAC AGC TAG GGC CCA GGC-3′.
MAbs and flow cytometry
TY25 (anti-B7-DC, rat IgG2a, κ) and RMP1-14 (anti-PD-1, rat IgG2a, κ) were generated as previously described.(4,34) A new hybridoma that produced anti-B7-DC MAb (MIH37, rat IgG2a, κ) was generated by immunizing Sprague-Dawley rats with B7-DC/L5178Y cells and fusing immunized splenocytes with P3U1 myeloma cells. MIH37 was selected on the basis of binding to B7-DC-transfectants but not to parental cells. MIH37 did not show any bindings to other B7 family molecule-transduced cells. TY25, RMP1-14, and MIH37 were purified from ascites using the caprylic acid and ammonium sulfate precipitation method. Purity was verified by SDS-PAGE analyses. Biotinylation was performed according to a standard protocol. PerCP-Cy5.5-conjugated anti-CD3 (145-2C11), allophycocyanin (APC)-conjugated anti-CD4 (GK1.5), FITC-conjugated anti-CD8 (53-6.7), and PE-conjugated anti-CD25 (7D4) MAbs were used. All MAbs were obtained from either eBioscience (San Diego, CA) or BD Biosciences (San Jose, CA). Culture supernatant from 2.4G2 (anti-CD16/CD32) hybridoma was used to block nonspecific binding. Immunofluorescence, flow cytometry, and data analysis were performed using a FACSCalibur flow cytometer using the CellQuest Pro (BD Biosciences) or the FlowJo (TreeStar, Ashland, OR) software.
Assessment of binding properties
To assess binding affinity, B7-DC/L5178Y cells were incubated with graded doses (0.004, 0.015, 0.06, 0.25, or 1 μg/1×106 cells) of either MIH37 or TY25, followed by biotinylated anti-rat IgG and PE-streptavidin (eBioscience). Flow cytometry was performed and mean fluorescence intensity (MFI) was estimated as a representation of bound MAbs. Association constants (Ka) were determined by Scatchard plot analysis.(33) The dissociation constant (Kd) was calculated as 1/Ka. To assess competitive binding between MIH37 and TY25 to B7-DC, B7-DC/L5178Y cells were incubated with serially diluted MIH37 or TY25 (0, 0.008, 0.04, 0.2, 1, or 5 μg) as a competitor and biotinylated TY25 or MIH37 (1 μg) as a detector, followed by PE-streptavidin. To measure the inhibitory effects on binding between B7-DCIg and PD-1, B7-DCIg (0.1 μg) was mixed with graded doses of TY25 or MIH37 (0, 0.6, 1.25, 2.5, 5, or 10 μg); PD-1/J558L cells were then stained with these mixtures, followed by PE-anti-human IgFc Ab (Jackson Laboratory, West Grove, PA). To inhibit PD-1Ig binding to B7-DC, B7-DC/L5178Y cells were preincubated with graded doses of TY25 or MIH37 (0, 0.3, 0.6, 1.25, 2.5, or 5 μg) as inhibitors, and PD-1Ig (1 μg) was then added to each sample, followed by PE-anti-human IgFc Ab. All samples were analyzed by flow cytometry, and the mean fluorescence intensity (MFI) of each sample was determined. The percentage of antibody binding inhibition was calculated using the following formula:
For some experiments, PD-1/J558L cells were stained with 1 μg of biotinylated control hIgG1, B7-H1Ig, or B7-DCIg preincubated with 10 μg of isotype control, TY25, or MIH37, followed by streptavidin-PE.
Induction of contact hypersensitivity and MAb treatment
Contact hypersensitivity (CH) to dinitrofluorobenzene (DNFB; Sigma, St. Louis, MO) and anti-B7-DC and/or anti-PD-1 MAb treatment was performed as described previously.(9) Briefly, 0.5% DNFB in acetone-olive oil (4:1) was painted onto the abdominal skin on days 0 and 1, and then 0.2% DNFB was challenged on the ear on day 5. A rechallenge was performed 35 days after the initial challenge. For MAb treatment, 200 μg/body of either control rat IgG (MP Biomedicals, Irvine, CA), MIH37, RMP1-14, or both MAbs together was injected i.p. 2 h before each sensitization or challenge. Ear thickness was measured as described previously.(9) Statistical analyses were performed using a Mann-Whitney U-test. Values of p<0.05 were considered statistically significant.
Functional analyses of draining LN cells
Draining LN (dLN) cells were isolated by collagenase digestion from cervical, axillary, and inguinal LNs of sensitized and treated mice 4 days after the final sensitization.(35) Flow cytometric analyses were performed using whole LN cells. LN T cells were isolated using negative selection with a magnetic cell-sorting system (MACS, Miltenyi Biotec, Bergisch Gladbach, Germany) and a mixture of biotinylated MAbs, including anti-CD49b, anti-CD45R, anti-I-A/I-E, and either anti-CD4 or anti-CD8 MAbs, as described previously.(35) T cell purity exceeding 94% was confirmed using flow cytometry. Isolated LN T cells (3×105/well) were co-cultured with the same number of 2,4-dinitrobenzene sulfonic acid (DNBS; Tokyo Kasei Kogyo, Tokyo, Japan)-pulsed splenocytes for 4 days, and IFN-γ production in the culture supernatants were measured by ELISA as described previously.(9,35)
Evaluation of ovalbumin-specific responses
B6 mice or BALB/c background PD-1-deficient mice were subcutaneously immunized with CFA-emulsified chicken ovalbumin (OVA, 25 μg/foot pad). Ten days after immunization, dLN cells were collected from inguinal and popliteal LNs and isolated as described above. Whole LN cells (4×105/well) were cultured with graded doses (0, 10, 100, and 1000 μg/mL) of OVA in the presence of either control rat IgG, MIH37, TY25, RMP1-14, MIH37 and RMP1-14, or TY25 and RMP1-14 (20 μg/mL each) for 4 days. IFN-γ production was measured by ELISA.
Results
MIH37 shows a higher binding affinity to B7-DC
To compare binding properties between a previous anti-B7-DC MAb (TY25) and a new anti-B7-DC MAb (MIH37), we evaluated the number of MAbs bound to cell surface B7-DC on B7-DC/L5178Y transfectants. Constant cell numbers of transfectants were stained with graded amounts of each MAb, and MAb binding was assessed by fluorescence intensity. Staining with 1 μg of both MIH37 and TY25 exhibited comparable levels of high fluorescence intensity (Fig. 1A), and increased amounts of MAb did not further enhance the fluorescence intensity (data not shown). These results suggest that 1 μg is the saturating amount for both MAbs. Fluorescence intensities of lower amounts of staining (0.06 and 0.004 μg) with TY25 decreased in a dose-dependent manner, while the decrease in MIH37 staining was minimal. Additionally, consistently higher binding as compared with TY25 was maintained, thus suggesting higher binding affinity. Scatchard plot analyses revealed that the bound MAb of MIH37 to B7-DC was persistently higher and the calculated Kd was 0.60 and 2.33×10–9 M for MIH37 and TY25, respectively. These results suggest an approximately three- to four-fold higher binding affinity of MIH37 to cell surface B7-DC. Next, we examined competitive binding ability between MIH37 and TY25. The addition of excess amounts (5 μg) of competitive MAb prior to the saturated amount (1 μg) of biotinylated detection MAb showed almost complete inhibition in both cases (Fig. 1B). However, when different MAbs were used in a crisscross manner as competitor and detector, binding was differentially inhibited. The excess amount of MIH37 efficiently inhibited the binding of TY25, whereas the same amount of TY25 only partially inhibited the binding of MIH37. The plots of percent inhibition at the indicated competitor doses consistently showed a higher competitive binding ability of MIH37 to B7-DC. These data indicate that MIH37 possesses a higher binding ability than TY25, and both antibodies bind to the same or close epitopes on B7-DC.
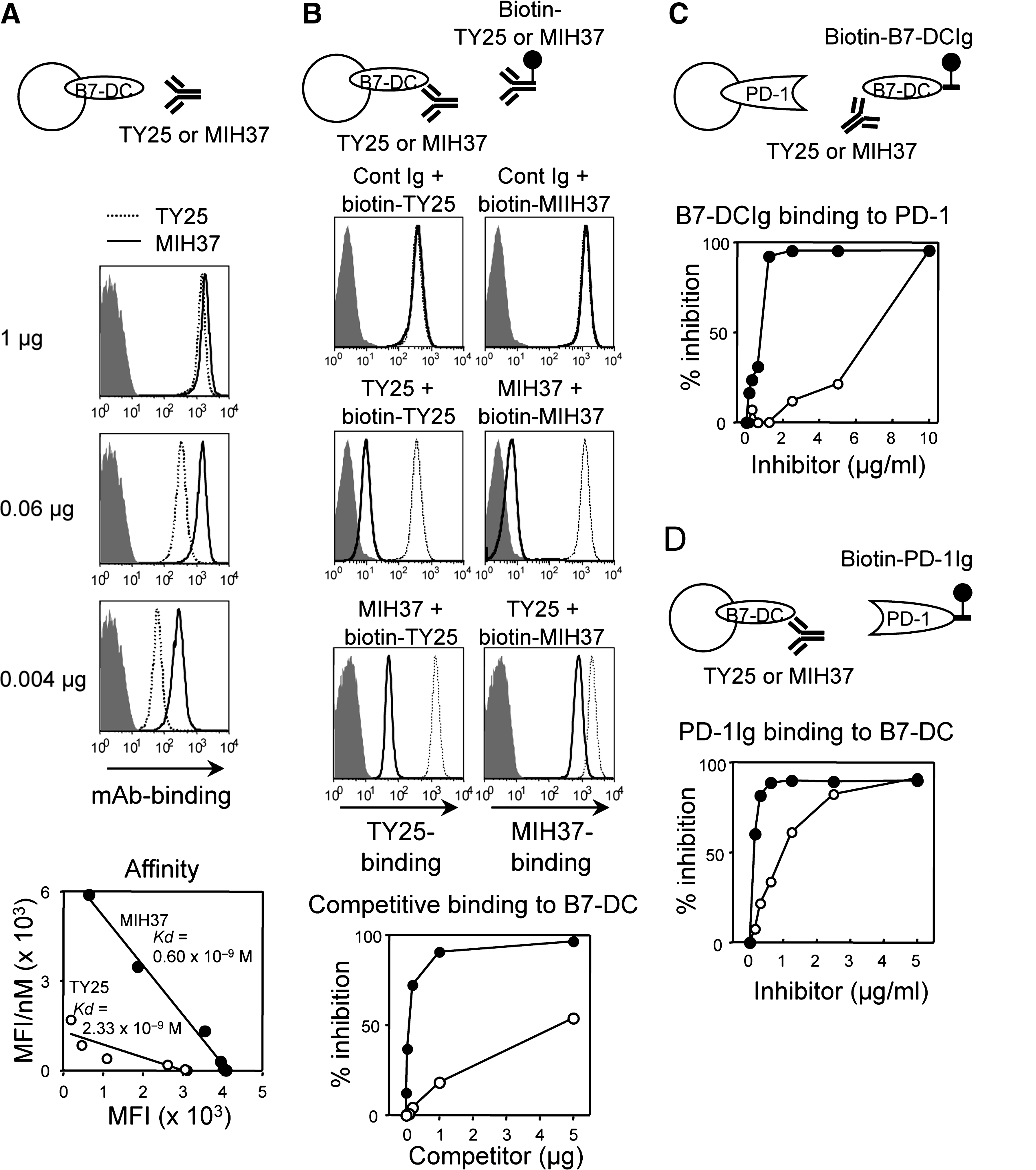
Differential binding properties between MIH37 and TY25. (
MIH37 inhibits B7-DC:PD-1 binding more efficiently
To examine blocking ability, we assessed the inhibitory effects of both MAbs on B7-DCIg binding to cell surface PD-1 (Fig. 1C), and PD-1Ig binding to cell surface B7-DC (Fig. 1D). Although preincubation of a high amount (10 μg) of either MAb with B7-DCIg almost completely inhibited B7-DCIg binding to cell surface PD-1, the inhibitory effects of TY25 rapidly decreased after titration (Fig. 1C). In contrast, MIH37 persistently displayed higher inhibitory effects. Similarly, MIH37 exerted higher inhibitory effects over TY25 in the case of PD-1Ig binding to cell surface B7-DC (Fig. 1D). These results suggest that both MAbs function as blocking antibodies for the B7-DC:PD-1 interaction; however, MIH37 more efficiently inhibits B7-DC:PD-1 binding as compared with TY25.
Treatment with MIH37 inhibits CH responses
Previously, we investigated the effects of anti-B7-H1 (MIH5, MIH6, and MIH7), anti-B7-DC (TY25), and anti-PD-1 (J43) MAb treatment in hapten-induced CH responses and demonstrated the preferential involvement of B7-H1 in PD-1-mediated regulation of CH responses.(9) Treatment with either anti-PD-1 or anti-B7-H1 MAb at sensitization significantly enhanced ear swelling, whereas the effects of anti-B7-DC (TY25) MAb treatments were minimal. We examined the effects of this new MIH37 MAb on CH responses. Notably, treatment with MIH37 at sensitization significantly inhibited ear swelling at earlier time points (24 and 48 h) (Fig. 2A, left panel). These inhibitory effects were long-lasting even when a rechallenge was performed 4 weeks after the initial challenge (Fig, 2A, middle panel). Our previous report demonstrated that treatment with anti-PD-1 MAb at challenge has minimal effects as compared with treatment at sensitization. Unexpectedly, treatment with MIH37 at challenge drastically inhibited ear swelling (Fig. 2A, right panel). These results suggest that treatment with MIH37 inhibits immune responses against hapten. We next examined the status of dLN cells 5 days after hapten sensitization and MIH37 treatment. Total dLN cell numbers in the MIH37-treated mice were comparable to those of control mice (data not shown). The percentages of CD3+ and CD8+ T cells were significantly lower in MIH37-treated mice (Fig. 2B). In addition, the percentages of CD25+ activated cells within CD4+ T cells significantly decreased in MIH37-treated mice. We have not observed any differences in a ratio of Foxp3+CD4+ regulatory T cells (data not shown). IFN-γ production from dLN T cells that were stimulated with hapten-pulsed splenocytes was also inhibited in MIH37-treated mice (Fig. 2C). These results indicate that treatment with MIH37 downregulates hapten-specific T cell activation, resulting in inhibition of CH responses.

Treatment with MIH37 inhibits CH responses. (
Blockade or lack of PD-1 affects the inhibitory actions of MIH37
PD-1, a known counter receptor for B7-DC, regulates T cell activation. We next examined whether absence of the PD-1 receptor affects the inhibitory actions of MIH37 in CH responses. Consistent with our previous observations, blockade of PD-1 with anti-PD-1 MAb (RMP1-14) dramatically enhanced ear swelling at all time points (Fig. 3A). Co-treatment of anti-PD-1 MAb with MIH37 did not affect ear swelling at the earlier time point (24 h), but reversed the inhibitory effects of MIH37 at 48 h. We further examined the effects of MIH37 treatment in PD-1-deficient mice. Ear swelling was maintained at a higher level in PD-1-deficient mice (Fig. 3B). Treatment with MIH37 did not inhibit ear swelling in PD-1-deficient mice at any time point. These results indicate that the inhibitory effects of MIH37 are dependent upon the regulatory functions of PD-1.
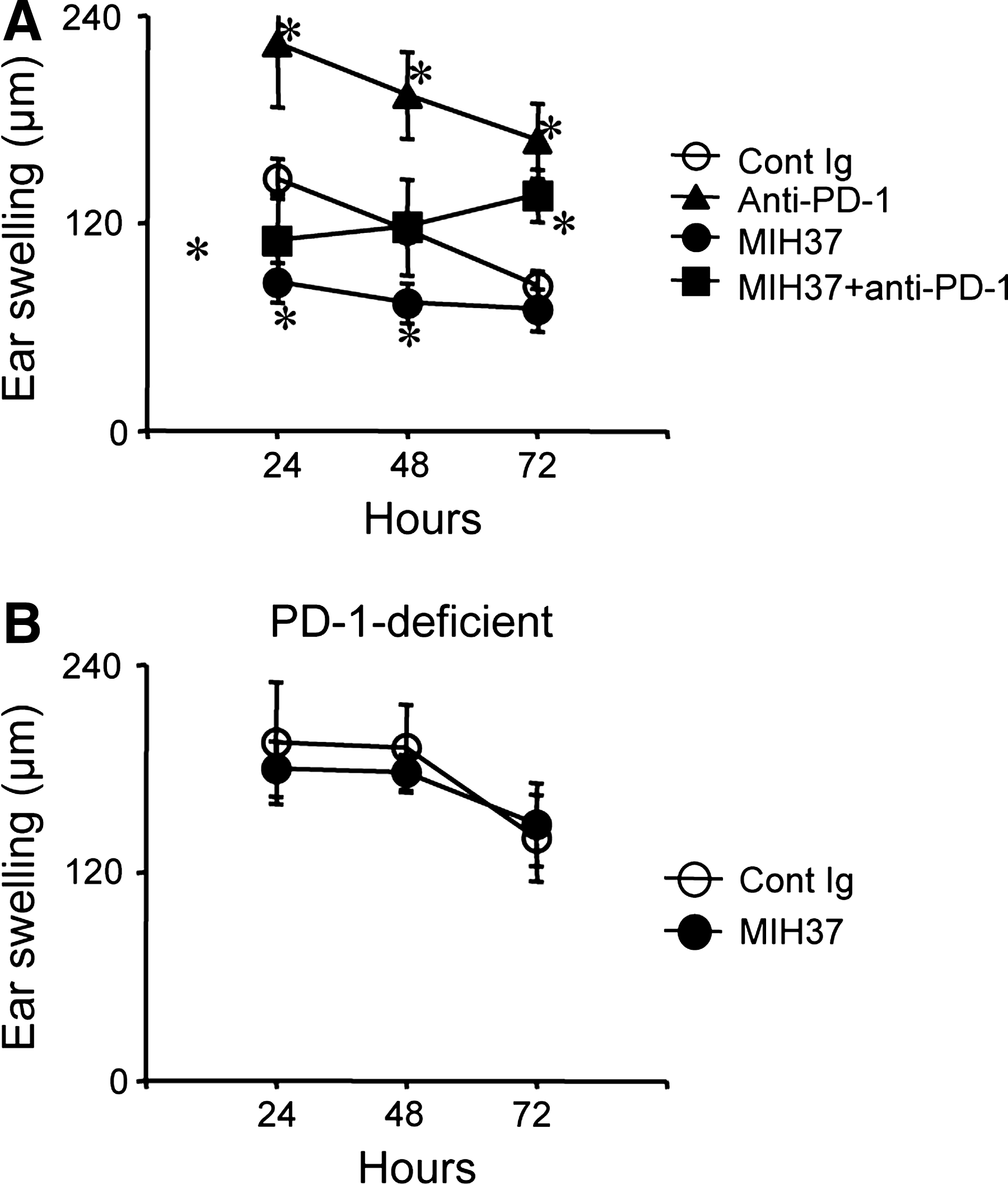
(
Effects of MIH37 on T cell responses against OVA
To further confirm whether the inhibitory effects of MIH37 can be seen in antigen-specific T cell responses in vitro, we examined T cell responses against OVA. dLN T cells obtained from OVA-immunized mice were restimulated in the presence of each MAb, and proliferative responses and IFN-γ production were measured. Consistently, the addition of MIH37 inhibited IFN-γ production in response to OVA, whereas the addition of TY25 enhanced this response (Fig. 4A). When we examined combinational effects with anti-PD-1 MAb at 1000 μg/mL of OVA, the addition of anti-PD-1 MAb together with MIH37 cancelled the inhibitory effects of MIH37 and enhanced IFN-γ production (Fig. 4B). The addition of anti-PD-1 MAb alone markedly enhanced IFN-γ production. Enhanced IFN-γ production by TY25 was not affected by further addition of anti-PD-1 MAb. In a similar experiment using T cells from PD-1-deficient mice, IFN-γ production was not affected by the addition of either MIH37 or TY25 (Fig. 4C), suggesting that the opposite effects of both anti-B7-DC MAbs are dependent on PD-1. Our results demonstrate that MIH37 and TY25 show opposite effects on T cell responses against OVA, but both effects are dependent on PD-1.
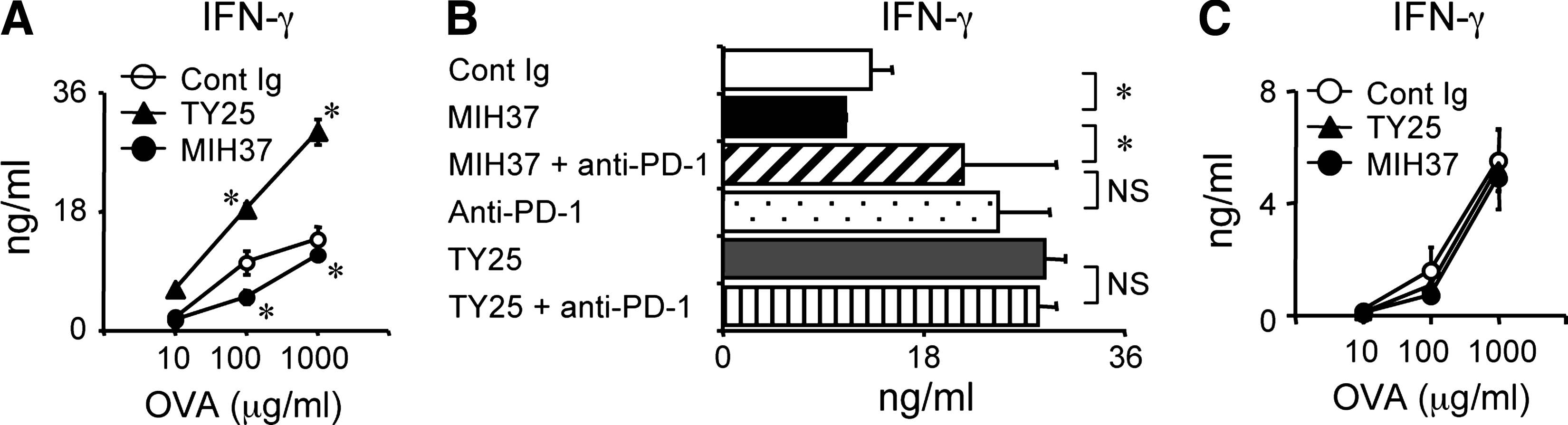
MIH37 inhibits in vitro T cell responses against OVA. dLN cells from B6 (
Discussion
In this study, we demonstrated that a newly established anti-B7-DC MAb, MIH37, possesses an approximately four-fold higher binding affinity to cell surface B7-DC as compared with TY25. In addition, it exerts stronger inhibitory effects on B7-DC:PD-1 binding. Despite differential binding properties to B7-DC, epitopes on B7-DC appear to be close or identical because the saturated amounts of each MAb mostly inhibit the binding of another. Nevertheless, we have shown that the administration or addition of these MAbs in vivo or in vitro shows opposite effects on T cell-mediated immune responses.
Previously, we demonstrated that PD-1-mediated regulation is involved in the induction phase of CH responses and that B7-H1 is dominantly involved in this regulation.(9) The administration of TY25 at sensitization slightly enhanced CH responses, although this effect was not significant. In contrast, here we show that similar treatment with MIH37 inhibits ear swelling at earlier time points (24 to 48 h). Inhibition of T cell responses by MIH37 treatment was also confirmed by decreases in dLN-T cell expansion, CD25 induction, and IFN-γ production against hapten-pulsed antigen-presenting cells. Marked opposite effects between MIH37 and TY25 were observed in OVA-specific T cell responses in vitro. These results indicate dual co-stimulatory and co-inhibitory roles of B7-DC.
To explain this phenomenon, we initially hypothesized the existence of another receptor other than PD-1 for B7-DC that transduces positive co-stimulatory signals; an undiscovered secondary co-stimulatory receptor for B7-DC has been previously suggested.(27,28,30) However, our results from studies using PD-1-deficient mice or co-blockade of PD-1 with MIH37 have revealed that the inhibitory effects of MIH37 are also dependent on PD-1 (Figs. 3 and 4). Surprisingly, co-administration of anti-PD-1 MAb and MIH37 did not affect the inhibitory effects of MIH37, but rather diminished the enhancing effects of anti-PD-1 MAb. Consistently, similar results were obtained in experiments of OVA-specific T cell responses in vitro. These results indicate that B7-DC-mediated co-stimulatory functions require PD-1 and that the blockade of B7-DC-mediated co-stimulation by MIH37 also neutralizes regulatory functions via PD-1. These results suggest that B7-DC-mediated co-stimulatory functions and PD-1-mediated regulatory functions are closely linked.
What and how control positive and negative functions via the PD-1 receptor? We have not observed any evidence that MIH37 directly induces signaling via B7-DC, and the previous reports demonstrating B7-DC-mediated reverse signals were all retracted. B7-H1 and B7-DC have been shown to cross-compete for PD-1 binding and to exhibit comparable affinities, but differential association and dissociation properties.(36) It has been shown that B7-H1 interacts with CD80 for inducing inhibitory signals in T cell activation.(37) B7-H1 binds to both PD-1 and CD80 with similar association and dissociation mechanisms.(36) Thus, the blockade of B7-DC:PD-1 interaction by the MIH37 with higher affinity seems to affect B7-H1:PD-1 and B7-H1:CD80 interactions. This may modulate contribution of co-signal receptors including PD-1, CD80, CTLA-4, and CD28, since these functionally opposite co-signal receptors compete and accumulate to the immunological synapse and the final availability of receptors are determined by ligand affinity and expression levels.(38–40) Although we cannot clarify the precise mechanisms, our results suggest greater complexities of PD-1-mediated functions induced via ligand binding for controlling immunity and tolerance.
Footnotes
Acknowledgments
We thank T. Honjo (Kyoto University, Japan) for kindly providing the PD-1-deficient mice; T. Kitamura (Institute of Medical Science, University of Tokyo, Japan) for kindly providing retroviral vectors and a packaging cell line Plat-E; and A. Yoshino and S. Miyakoshi for cell sorting. This study was supported in part by Grants-in-Aid for Scientific Research from the Ministry of Education, Culture, Sports Science and Technology of Japan (22021016, M.A.) and from Japan Society for the Promotion of Science (20249075, M.A.).
Author Disclosure Statement
The authors have no financial conflicts to disclose.