
Abstract
Histone modifications are important components of transcriptional regulation and chromatin-based regulatory processes. In addition, WD40-repeat protein and several other components are involved in these functions. Here we present the development of monoclonal antibodies (MAbs) against Arabidopsis HOS15, a WD40-repeat protein. Mice were immunized with recombinant HOS15 (rHOS15) protein for generating MAbs via the classic hybridoma production technique. We confirmed the specific activity of anti-HOS15 MAbs by tobacco transient expression assays. Based on immunoprecipitation assays, the anti-HOS15 MAb was able to detect endogenous HOS15 in Arabidopsis wild-type plants, but not in hos15 mutant plants. Finally, the anti-HOS15 MAbs are highly sensitive for detecting endogenous HOS15 protein. They can be used for immunological and immunoprecipitation assays to support other experimental strategies. In particular, the anti-HOS15 MAbs will be essential tools to investigate the role of HOS15 in the regulation of tolerance to environmental stresses in plants.
Introduction
The relative levels of hyper- or hypoacetylated histone proteins are known to be determined by the enzymatic activities of both histone acetyl transferases (HATs) and histone deacetylases (HDACs).(9) The acetylation status of histones at specific DNA regulatory sequences depends on the recruitment of histone acetyltransferases or histone deacetylases to a precise location by specific DNA binding proteins and histone interacting proteins. Acetylation of the histone tails might not only release some DNA from nucleosomes but also loosen interactions between adjacent nuclesomes, thus altering the access of transcription factors.(7) Moreover, acetylation of histone proteins regulates DNA damage repair and DNA replication-dependent chromatin assembly.(7,10) GCN5 acetylates lysine-14 of histone H3 and lysine-8 and lysine-16 of histone H4, respectively.(11) In contrast, histone deacetylases such as Rpd3p can repress gene activity based on the recruitment of proteins that bind specific promoters or methylated DNA.(9) Deacetylation of H4 Lys-16 by Sir2p promotes the interaction of Sir3p with histone H4, allowing the folding into heterochromatin to spread along the histone tails.(7) These chromatin modifiers play an essential role in controlling gene expression and in establishing epigenetic marks that may be inheritable.(12)
The existence of a direct molecular link between the histone acetylation status and transcriptional co-activators, including GCN5, PCAF, CBP/p300, and SRC-1/ACTR, shows intrinsic histone acetyltransferase activity. In addition, transcriptional repressors such as NuRD, SIN3, Groucho/Tup1, and SMRT/N-CoR associate with histone deacetylases.(13–15) Among them, the co-repressors SMRT and N-CoR exist in large protein complexes with estimated sizes of 1.5–2 MDa in HeLa cell nuclear extracts.(16) The SMRT complex includes histone deacetylase 3 (HDAC3) and transducin (β)-like I (TBL1), a WD-40 repeat-containing protein.(16) TBL1 was originally identified as a novel gene that was either partly or entirely deleted in patients carrying Xp22.3 terminal deletions. This led to the suggestion that TBL1 would be involved in the pathogenesis of the ocular albinism with late-onset sensorineural deafness phenotype.(17) In addition, TBL1 homologs exist in yeast and Drosophila.(17) In particular, Drosophila genomic sequences showed high identity (>80%) to the human TBL1 gene, suggesting that TBL1 is evolutionarily highly conserved.
In plants, histone modification and chromatin remodeling have been shown to take part in various developmental processes and to be involved in metastable changes required to maintain altered cellular and tissue properties.(12,18) Although correlations between chromatin remodeling and developmental processes in plants have frequently been demonstrated, the influence of chromatin remodeling on adaptation responses to the environment is still poorly understood. Recently, the osmotically responsive gene 15 (HOS15), a WD40-repeat protein, was identified in a forward genetic screen for mutations that alter abiotic stress signaling. HOS15 was shown to function in the control of gene expression through histone deacetylation in Arabidopsis.(19–21) Mutants of hos15 accumulated higher levels of transcripts of many stress-regulated genes and showed a hypersensitive phenotype with respect to freezing stress. HOS15 protein shared high similarity with the human TBL1 protein, a component of the chromatin repressor complex in histone deacetylation. HOS15 also interacts with histone H4, and the level of acetylated histone H4 is higher in hos15 mutants compared to that in the wild types.(19)
HOS15 plays a key role in chromatin remodeling, abiotic stress responsive gene expression, and stress tolerance in plants.(22) However, identification of other components that are a part of the repressor protein complex in which HOS15 is involved and its direct target genes remain elusive. Therefore, specific antibodies are required to detect endogenous HOS15 protein in plant tissues. Here, we expressed Arabidopsis HOS15 in E. coli. The recombinant HOS15 protein was used as the antigen to generate anti-HOS15 monoclonal antibodies (MAbs) that specifically detect endogenous HOS15 in plant tissue samples. The anti-HOS15 MAbs will be useful tools for investigating the role of HOS15 in gene regulation and chromatin structure dynamics of tolerance to various environmental stresses in plants.
Materials and Methods
Cloning of Arabidopsis HOS15
Arabidopsis HOS15 cDNA inserted to pGEX-2T plasmid DNA was used as a PCR template and the resulting HOS15 cDNA was transformed into an E. coli expression vector. For the expression of His6-tag HOS15 protein in E. coli, AtHOS15 cDNA was amplified with the following primers: forward: (5′-TTTAGAATTCATGTCTTCACTTACCTCC-3′) and reverse (5′-TAAACTCGAGCTA CATTCTGAAATCAAG-3′). The amplified HOS15 cDNA carried an EcoRI recognition site at its 5′-end and XhoI recognition site at the 3′-end. The digested pPROEX HTa vector (Invitrogen, Carlsbad, CA) and the PCR product were ligated with T4 DNA ligase (NEB, Ipswich, MA), and the ligated construct was transformed into E. coli DH5α.
Recombinant protein expression and purification
The His6-tag recombinant HOS15 protein (rHOS15) was expressed in E. coli cultured in LB broth containing 100 μg/mL ampicillin. The cultures were incubated at 37°C with constant shaking, until the optical density at 600 nm reached 0.5–0.6. The rHOS15 was induced by adding 0.6 mM isopropyl β-D-thiogalactopyranoside (IPTG) and further cultured at 37°C with constant shaking for 3 h. The bacteria were harvested by centrifugation at 6500 g for 20 min at 4°C. The cell pellet was re-suspended in 40 mL of lysis buffer (20 mM Tris-HCl [pH 7.0], 0.3 M NaCl, 8 M urea). The re-suspended cells were disrupted by sonication and solubilized at 37°C with constant shaking overnight. The soluble proteins were harvested by centrifugation at 6500 g for 20 min at room temperature (RT). The supernatant was loaded onto a cobalt conjugated bead column (Clontech, Mountain View, CA), and the column was washed with 50 times bed volumes of washing buffer (20 mM Tris-HCl [pH 7.0], 0.3 M NaCl, 8 M urea) containing 0.1% of Triton X-114 for endotoxin removal. Then the His6-tag rHOS15 protein was eluted using elution buffer (20 mM Tris-HCl [pH 7.0], 0.3 M NaCl, 8 M urea, 0.3 M imidazole).
The purified rHOS15 protein was separated by SDS-PAGE and stained with Coomassie Blue R-250 or blotted onto nitrocellulose membrane (Whatman, Maidstone, United KIngdom). The membranes were blocked with 5% non-fat dry milk in 0.1% Tween-20/PBS (PBST) and incubated with primary antibody, mouse anti-His6-tag MAb (1 μg/mL) (R&D Systems, Minneapolis MN) at RT for 2 h. After washing, the membrane was incubated with horseradish peroxidase (HRP)-coupled goat anti-mouse IgG (Jackson ImmunoResearch, West Grove, PA) for 1 h at RT and developed by using Western blot detection reagent (NeuroNex, Pohang, Korea). Photographs of developed membranes were taken by RAS4000 (Fuji, Tokyo, Japan).
Immunization and selection of hybridoma cells
The BALB/c mice were reviewed and approved by the Konkuk University Institutional Animal Care Committee. To generate monoclonal anti-HOS15 antibodies, purified rHOS15 protein was used to immunize female BALB/c mice (5 weeks of age) by subcutaneous injections. A mixture of Freund's complete adjuvant (Sigma-Aldrich, St. Louis, MO) and rHOS15 protein was used in one immunization. After 2 weeks, the mixture of Freund's incomplete adjuvant (Sigma-Aldrich) and rHOS15 protein was used for three immunizations applied in 1 week intervals. The titers of each mouse serum were determined by 1 μg/mL rHOS15 coated 96-well immune plate. Finally, the last boost of antigen was injected 3 days before a fusion.
The FO myeloma cell line was fused with splenocytes from immunized mice according to standard protocols. The hybridoma cells of positive wells were cloned by limiting dilution in aminopterin-free and HT containing Dulbecco's modified Eagle's medium (DMEM) with 20% fetal bovine serum. Single clones were screened by direct ELISA and Western blotting. A single hybridoma clone was expanded and inoculated to BALB/c mice for large-scale expression of antibodies by intraperitoneal injection.
Direct ELISA
For direct ELISA, a Maxisorp plate (SPL) was coated with 1 μg/mL of rHOS15 protein and incubated overnight at 4°C. Non-specific protein binding sites were blocked with 3% BSA in PBS for 1 h at RT. The plate was washed with PBS containing 0.1% Tween-20 (PBST). Sera from immunized mice, 10-fold serially diluted in PBS, and cell culture supernatants were added and incubated at RT for 2 h in the coated plate. After washing with PBST, horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG antibody (Jackson ImmunoResearch) was added for 40 min at RT. After washing, 3,3′,5,5′-tetramethylbenzidine base substrate (TMB) was added. The color reaction was stopped by adding 0.1 mL of 1 M H2SO4 to each well, and the plate was examined at 450 nm in an ELISA reader.
Western blot analysis
The rHOS15 protein was boiled for 5 min and loaded on the 10% SDS-PAGE. Samples were electro-transferred onto a nitrocellulose membrane and blocked with 5% skim milk in PBST. Membrane was probed with ascitic fluid of hybridoma or anti-His6-tag antibody as primary antibodies, and the bound antibodies were detected by HRP-conjugated goat anti-mouse IgG antibody (Jackson ImmunoResearch) at dilution of 1:10,000 followed by ECL substrate solutions (Amersham Biosciences, Uppsala, Sweden). Photographs of developed membranes were taken by RAS4000 (Fuji).
For transient expression of AtHOS15 protein in tobacco, combinations of Agrobacterium tumefaciens (strain GV3101) bacteria containing constructs (35S::vector control and 35S::AtHOS15) were infiltrated into leaves of tobacco (Nicotiana benthamiana) plants. Three-week old N. benthamiana plants were used for A. tumefaciens-mediated transformation. At 3 DAI (day after inoculation), Agrobacterium-infiltrated leaves were collected and proteins were prepared in protein extraction buffer (50 mM Tris-HCl [pH 7.5], 150 mM NaCl, 0.5% NP40, 1 mM EDTA, 3 mM DTT, 1 mM PMSF, 5 μg/mL leupeptin, 1 μg/mL aprotinin, 1 μg/mL pepstain, 5 μg/mL antipain, 5 μg/mL chymostatin, 2 mM Na2VO4, 2 mM NaF, 50 μM MG132). The proteins were subjected to immunoblot analysis using a 1-6-G clone MAb, which recognizes transiently expressed HOS15 protein in tobacco leaves.
Immunoprecipitation of HOS15 using 1-6-G clone antibody
Plant tissues were extracted in young Arabidopsis wild type (Young WT), young HOS15 mutant (HOS15 deletion mutant), adult Arabidopsis wild type (Adult WT), and adult HOS15 mutant (HOS15 deletion mutant). These extracts and ascitic fluid containing 1-6-G clone MAb were mixed for 4 h at 4°C, and protein G agarose bead (KPL) were added. After 1 h, beads were pelleted by centrifugation at 4°C and washed thrice with PBS containing 1% Triton X-100. The washed beads were boiled for 5 min and loaded on the 10% SDS-PAGE. Samples were separated by SDS-PAGE and blotted onto nitrocellulose membrane (Whatman). Membranes blocked with 5% non-fat dry milk in 0.1% Tween-20/PBS (PBST) were incubated with rabbit anti-HOS15 IgG at RT for 2 h. After washing, the membrane was incubated with HRP-coupled goat anti-rabbit IgG (Jackson ImmunoResearch) for 1 h at RT and developed by using Western blot detection reagent (Amersham Biosciences). Photographs of developed membranes were taken by RAS4000 (Fuji).
Results
Cloning of Arabidopsis HOS15 cDNA
The AtHOS15 cDNA was cloned into the pPROEX HTa plasmid vector to produce recombinant HOS15 protein in E. coli BL21 codon+. The PCR product of AtHOS15 cDNA is 1862 base pairs in length (Fig. 1A). The pPROEX HTa plasmid vector and amplified AtHOS15 cDNA were digested using EcoRI and XhoI, respectively, and ligated by T4 DNA ligase. The ligated plasmid was transformed into E. coli DH5α. After overnight culture, colonies were screened by PCR and the products were loaded on 1% agarose gels (Fig. 1B). Colony #6 was selected and cultured for plasmid DNA purification. The purified plasmid DNA was transformed in E. coli BL21 codon+ to express rHOS15 protein.
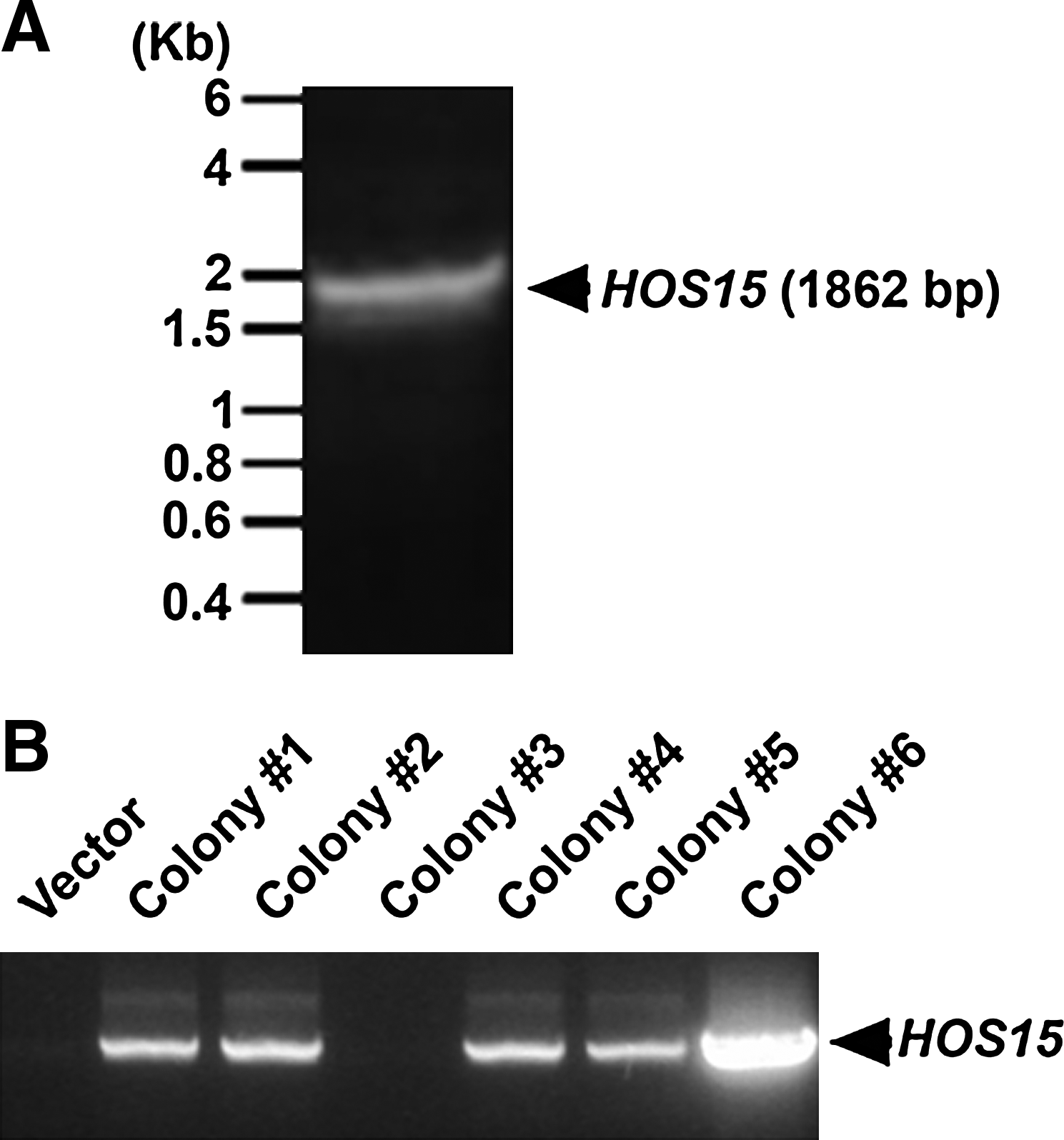
Cloning of Arabidopsis HOS15 cDNA. AtHOS15 cDNA was amplified with EcoRI and XhoI recognition sequences incorporated into primers. PCR product was loaded on 1% agarose gel (
Expression and purification of recombinant HOS15 for antigen
After expression of the rHOS15 protein in E. coli BL21 codon+, the rHOS15 protein was purified using cobalt-conjugated mini-affinity chromatography. The eluted rHOS15 protein fractions were separated on 10% SDS-PAGE and visualized by Coomassie Blue R-250 staining (Fig. 2A). As the positive control, gel-separated proteins were transferred to nitrocellulose membrane and incubated with anti-His6-tag antibody (Fig. 2B). The expected size of the His6-tag-fused rHOS15 protein was approximately 74 kDa (Fig. 2A, B). The results imply expression of rHOS15 in E. coli.
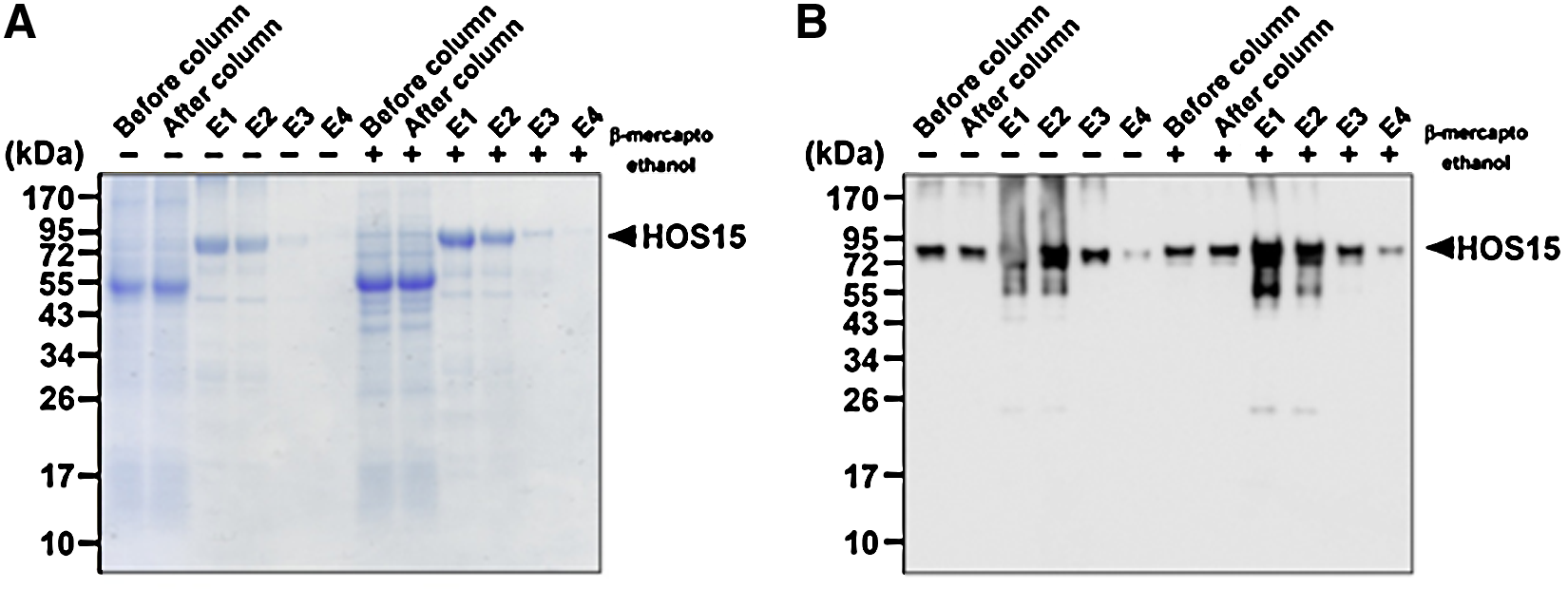
Preparation of recombinant His6-tag HOS15 protein for antigen. His6-tag rHOS15 protein was purified by mini-cobalt conjugated affinity chromatography. The eluted rHOS15 protein was separated by 10% SDS-PAGE under reduced (β-mercaptoethanol) or non-reduced condition and visualized by Coomassie Blue R-250 staining (
Production of MAbs against rHOS15 protein
To generate anti-HOS15 monoclonal antibodies (MAbs), we used His6-tag purified rHOS15 expressed in the BL21 codon+ strain. Three BALB/c mice were immunized with the purified rHOS15 protein and titrated with sera from the immunized mice (Fig. 3A). Mouse #2 showed the highest titer. Therefore, mouse #2 was selected and fused using splenocytes from the mouse, as described in the Materials and Methods section. For the screening of antibody producing hybridoma clones, rHOS15 was used for coating ELISA plates. We screened 108 hybridoma clones by direct ELISA and found three positive hybridoma clones (Fig. 3B). These three hybridoma clones (1-6-G, 4-8-F-1, 8-3-C-6) were adapted in HT-free DMEM with 10% fetal bovine serum.
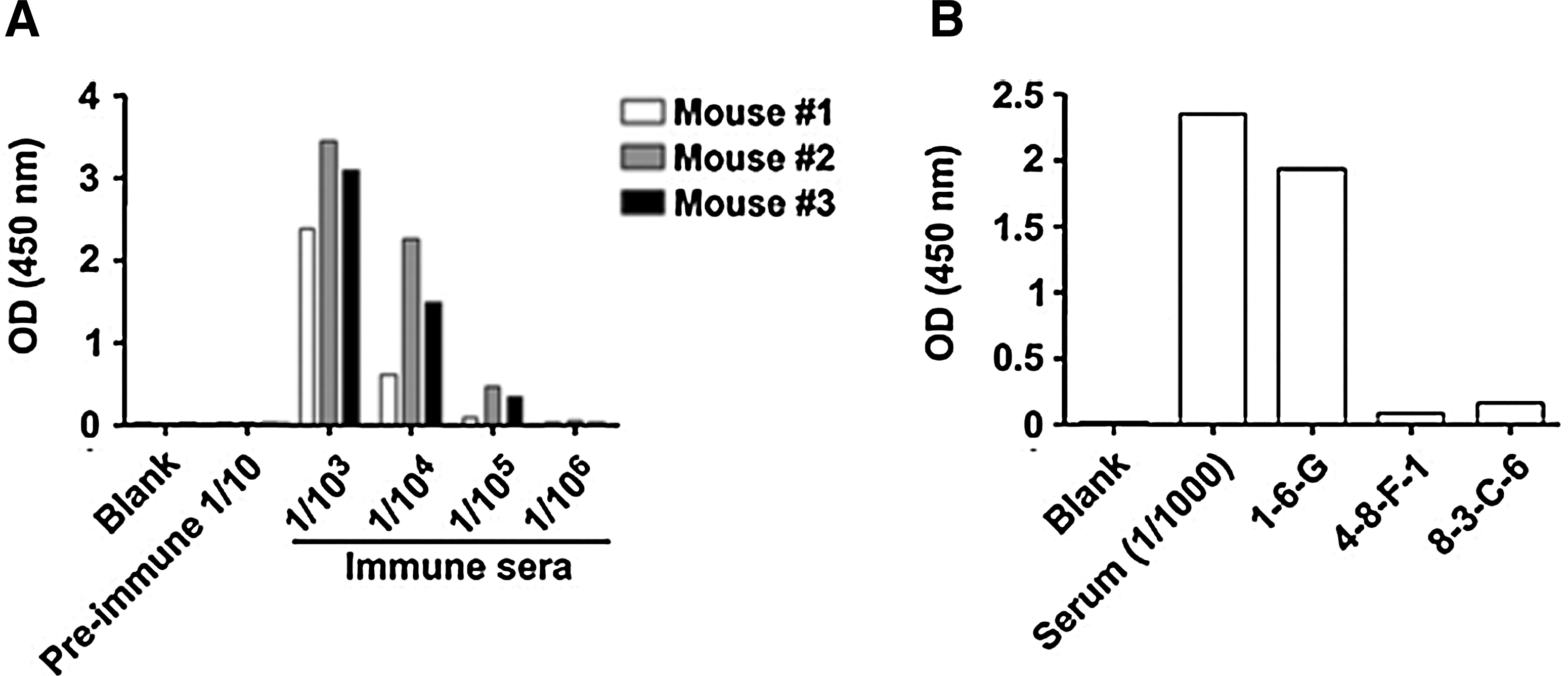
Immunization of rHOS15 to BALB/c mice and screening the hybridoma clones of anti-HOS15. Titers to rHOS15 were analyzed in serum samples (
Western blot analysis with selected anti-HOS15 MAbs
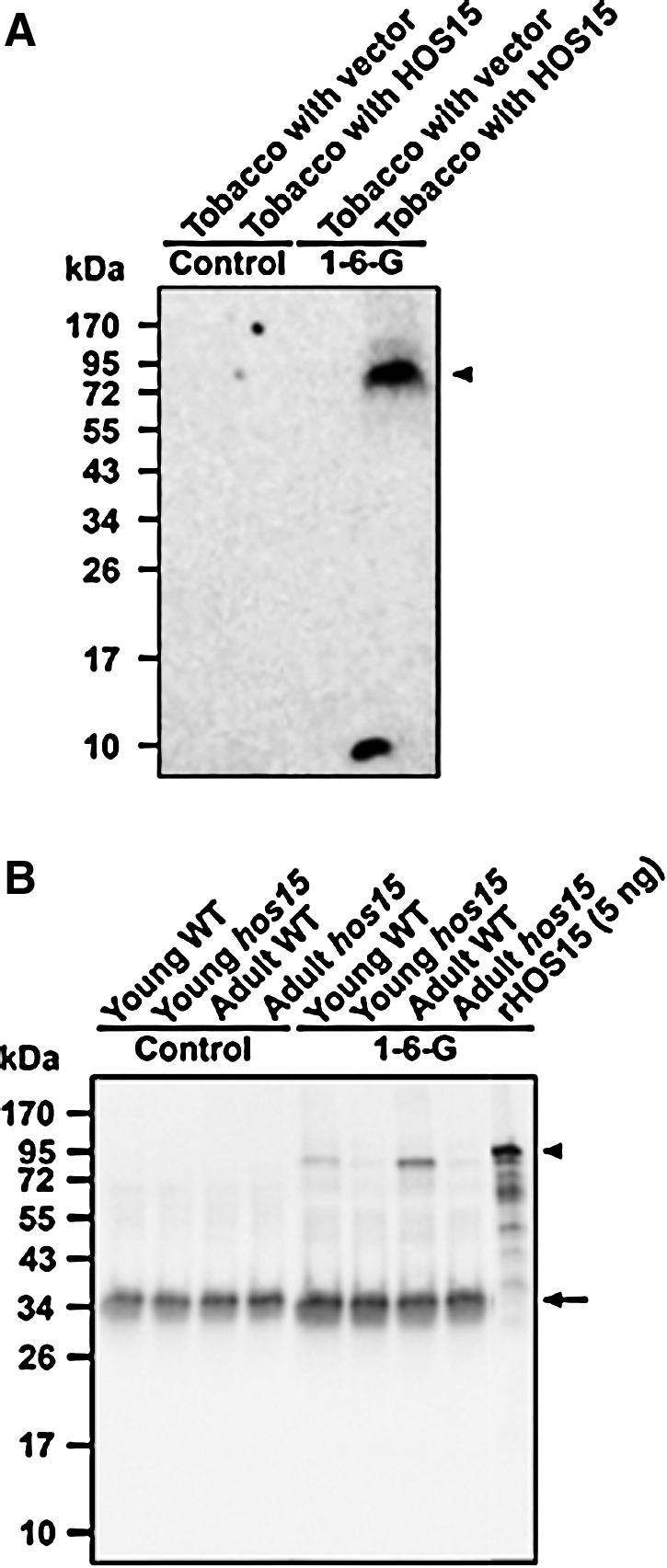
We performed Western blot analysis with rHOS15 protein as the antigen. The MAb clone 1-6-G recognized rHOS15 protein (50 ng/lane) (Fig. 4A) whereas other MAbs (4-8-F-1 and 8-3-C-6) failed to recognize rHOS15 in Western blot analysis under reducing condition (Fig. 4B, C). As control, we used anti-His6-tag antibody to detect 50 ng of rHOS15 and to show that equal amounts of rHOS15 were loaded in each lane (Fig. 4D). Therefore, the results suggested that MAb clone 1-6-G of HOS15 provided an excellent candidate for an anti-HOS15 MAb to detect endogenous AtHOS15 protein in Arabidopsis plants. In addition, Western blot analysis by a tobacco transient assay system was used (Fig. 5A). Agrobacterium tumefaciens (strain GV3101) containing two different constructs [35S::vector control (tobacco with vector) and 35S::AtHOS15 (tobacco with HOS15)] cells were infiltrated into leaves of tobacco (N. benthamiana) plants. The AtHOS15 protein was transiently expressed in tobacco leaves. After 3 days, Agrobacterium-infiltrated leaves were extracted with protein extraction buffer. The proteins were subjected to immunoblot analysis using a 1-6-G clone MAb. The 1-6-G clone MAb specifically detected the transiently expressed HOS15 protein in Agrobacterium-infiltrated tobacco leaves (Fig. 5A).

Immunoblot analysis with anti-HOS15 MAbs against rHOS15 protein. Two different concentrations of rHOS15 were subjected into SDS-PAGE gel and transferred to membrane. The membranes were probed with MAb clones 1-6-G (
Detecting the endogenous HOS15 with 1-6-G MAb. (
Detection of endogenous HOS15 in plant tissues using 1-6-G clone MAb
To examine the specificity of 1-6-G clone MAbs in detecting endogenous HOS15 in different plant tissues, immunoprecipitation was carried out with plant tissue lysates of young Arabidopsis wild type (Young WT), young hos15 deletion mutant (Young hos15), adult Arabidopsis wild type (Adult WT), and adult hos15 deletion mutant (Adult hos15). The immunoprecipitation samples were probed with mouse anti-HOS15 monoclonal antibody (1-6-G clone MAb). As shown in Figure 5B, the mouse anti-HOS15 monoclonal antibody detected endogenous HOS15 in young and adult Arabidopsis wild types, but no signal was observed in the leaves of either young or adult HOS15 mutants. The specificity of 1-6-G clone MAb to endogenous HOS15 protein showed no cross-reaction to other proteins. The result suggests that MAb clone 1-6-G specifically recognizes the endogenous HOS15 protein in Arabidopsis.
Discussion
Adaptations to counteract deleterious effects of environmental changes have evolved in all organism kingdoms. Such adaptations are generally genetic, based on gene regulation programs. It is also increasingly realized that adaptations can be plastic and reversible through equally genetically based programs that lead to chromatin remodeling. This important mechanism in the control of gene expression modulates the epigenetic state of cells. Major targets of epigenetic mechanisms are histone proteins, which are protein components of chromatin, and also DNA methylations that are marks for recognition and further modifications. Changes of histone proteins affect gene expression by RNA polymerase and contact levels of adaptor molecules.(13–15,23) Arabidopsis HOS15, a WD-40 repeat protein, is localized to the nucleus and functions as a repressor involved in the control gene expression mechanisms important to environmental stress tolerance through elimination of an acetyl group in the protein tail of histone H4.(19,20,24) In addition, HOS15 plays a central role in tolerance to abiotic stresses as well as plant growth.(19,21,25–28) Thus, we developed anti-HOS15 MAbs and these anti-HOS15 MAbs could be used for an effective and universal method to investigate the function of HOS15 in the stress tolerance mechanism of plants.
In this study, we cloned the Arabidopsis HOS15 cDNA into a protein expression vector and expressed rHOS15 protein (Figs. 1 and 2). The His 6 -tag rHOS15 protein was expressed in E. coli BL21 codon+, which resulted in very high yields of rHOS15 protein. The rHOS15 protein was present predominantly insoluble in inclusion bodies and could be solubilized in 8 M urea solutions. The solubilized rHOS15 protein was passed through a mini-cobalt affinity column and then further purified by HPLC. Finally, the purified recombinant HOS15 protein was used as an antigen to generate MAbs in mice (Fig. 2). Given the difficulty in generating highly specific monoclonal antibodies against plant proteins, Northern blot analysis has often been used as a preferred tool. However, monitoring the level of transcription is often irrelevant because proteins are the real reactors in the cells of living organisms. Protein studies are essential for solving signaling mechanisms and responses, and in this respect specific antibodies are required to advance our understanding.
We used a classical immunization protocol of PEG1500. As a result, we were able to generate three different hybridoma clones producing MAbs against rHOS15, which was identified by direct ELISA (Fig. 3B). Although three anti-HOS15 MAbs were positive with direct ELISA, only the 1-6-G MAb clone recognized rHOS15 (74 kDa) (Fig. 4). It seems likely that the two non-reactive clones might react with lower affinity to rHOS15 or to recognize the native, non-denatured rHOS15 protein. In order to examine the specificity of the 1-6-G clone MAbs to endogenous HOS15, we used HOS15-deleted Arabidopsis (mutant) and wild-type plants in immunoprecipitation. The endogenous HOS15 in wild type was pull-down by the 1-6-G clone MAb and detected by rabbit anti-HOS15 (Fig. 5B). Intriguingly, HOS15 protein was present in adult wild-type plants at much higher concentrations than in young wild-type plants. The result indicates that HOS15 protein may have functions associated with processes important during reproductive growth or during late stages of development. In addition, our results also document that the specificity of the anti-HOS15 MAb to endogenous HOS15 provides a very clean signal that shows extremely low cross-reaction to other proteins in Arabidopsis.
Since plants produce new cells to continue growing during and after adaptation (acclimation) to a new environment, we can expect chromatin-mediated metastable genetic changes to be involved in the process.(29,30) For example, an Arabidopsis cold-induced transcription factor that binds to C/DRE elements, CBF1, has been shown to associate with histone acetyltransferase proteins (HATs).(31) In order to study the response of endogenous HOS15 to environmental cues, it is essential that we examine the expression level of endogenous HOS15 protein under various stress conditions.
Collectively, the direct ELISA assay isolated three anti-HOS15 MAbs recognizing rHOS15 protein, and further immunoprecipitation assay revealed the 1-6-G clone MAb, which is the only clone able to immunoprecipitate endogenous HOS15 in plant tissues. The anti-HOS15 MAbs provide clear advantages for the detection of the native form of HOS15 in plants. This newly developed anti-HOS15 MAbs will be useful to study the biological activity of HOS15 in growth, reproduction, and tolerance to various environmental stresses.
Footnotes
Acknowledgments
We thank Dr. Hans J. Bohnert for his critical reading and insightful comments. This work was supported by grants from Brain Korea 21 Program and Korea Science and Engineering Foundation (KOSEF) funded by the Korea government (MOST: R01-2006-000-10837, WCU: R33-2008-000-10022-0, and KRF-2008-313-C00644) (to SK); and the World Class University Program (R32-10148), the Next-Generation BioGreen 21 Program (SSAC, PJ008025), Rural Development Administration, Republic of Korea (to D-JY).
Author Disclosure Statement
The authors have no financial interests to disclose.