
Abstract
Caveolae are critical cell surface structures important in coordinated cell signaling and endocytosis. One of the major proteins of caveolae is caveolin 1 (Cav-1). Cellular levels of Cav-1 are associated with cancer progression. In prostate cancer cells, levels of Cav-1 are positively correlated with tumor progression and metastasis. Cav-1 can be secreted by prostate cancer cells into the microenvironment and triggers proliferation and anti-apoptosis of the tumor and tumor endothelial cells. Clinical studies have shown increased serum Cav-1 levels in patients with poor prognosis. In tissue culture and animal model experiments, blocking secreted Cav-1 by polyclonal antibodies inhibits tumor cell growth. Cav-1 is therefore a potential therapeutic target for prostate cancer treatment. In this study, we used Cav-1 knock-out mice as hosts to produce monoclonal anti-Cav-1 antibodies. A total of 11 hybridoma cell lines were selected for their ability to produce antibodies that bound GST-Cav-1 but not GST on glutathione-coated ELISA plates. Further screening with ELISAs using GST-Cav-1 fragments on GSH-coated plates classified these antibodies into four groups: N1-31 with five antibodies binds the far N-terminus between amino acids 1 and 31; N32-80 with three antibodies binds between amino acids 32 and 80; CSD with two antibodies potentially bind the scaffolding domain (amino acids 80-101); and Cav-1-C with 1 antibody binds parts of the C-terminal half. Binding affinities (Kd) of these antibodies to soluble Cav-1 ranged from 10−11 to 10−8 M. Binding competition experiments revealed that these antibodies recognized a total of six different epitopes on Cav-1. Potency of these antibodies to neutralize Cav-1-mediated signaling pathways in cultured cells and in animal models will be tested. A selected monoclonal antibody will then be humanized and be further developed into a potential anti-prostate cancer therapeutic.
Introduction
Cav-1-mediated oncogenic activities in prostate cancer are linked to two pathways: first, overexpressed Cav-1 interacts directly with cellular signaling factors to stimulate proliferation and to activate anti-apoptotic mechanisms.(15–19) In cultured prostate cancer cells, up-regulated Cav-1 binds and inhibits protein phosphatases PP1 and PP2A and results in constantly active PI3K/Akt pathway.(18,20,21) Small interfering RNA (siRNA) treatment to reduce cellular levels of Cav-1 leads to reduced β-catenin and phosphorylated low-density lipoprotein receptor–related protein 6 (LRP6) and reverses androgen insensitivity in mouse prostate cancer cells.(6,22) In animal studies, development and progression of prostate tumors in Cav-1-/- mice are significantly reduced,(23,24) and overexpression of Cav-1 in prostatic epithelial cells leads to prostatic hyperplasia.(25,26) Second, Cav-1 can be secreted by prostate cancer cells into the tumor microenvironment to promote growth and angiogenesis.(27–29) Secretion of Cav-1 was first reported from normal pancreatic acinar cells,(30) and it might be a unique mechanism adopted by prostate cancer cells to promote malignant progression. Serum Cav-1 levels are significantly higher in men with prostate cancer than in men with benign prostatic hyperplasia,(31) and elevated pre-operative levels of serum cav-1 predict decreased time to cancer recurrence after radical prostatectomy.(32) Condition medium from cultured metastastic prostate cancer cells also has detectable levels of secreted Cav-1 proteins.(20) The secreted Cav-1 (or purified recombinant Cav-1) can be taken up by prostate cancer cells or endothelia cells in vitro and, through activating Akt- and/or NOS-mediated signaling pathways, promotes cell growth and angiogenesis.(24,33,34) In animal studies, inoculating high-passage LNCaP cells with high levels of Cav-1 on one side of a mouse will promote the growth of low-passage LNCaP cells with low levels Cav-1 inoculated on the other side of the same animal. Injection of polyclonal anti-Cav-1 antibodies in prostate cancer model mice suppressed the tumor growth and metastasis.(24) The autocrine and paracrine activities of secreted Cav-1 are therefore an ideal therapeutic target for prostate cancer.
Cav-1 gene can be expressed in two isoforms, Cav-1α and Cav-1β, which are derived from alternate initiation during translation.(35,36) Cav-1α is 178 amino acids in length. Cav-1β translation starts at methionine 32, lacking the first 31 amino acids at the N-terminus of Cav-1α. While both Cav-1α and Cav-1β carry the same membrane binding and scaffolding domains, as well as the entire C-terminal acylated domain, the potential functional differences between these two isoforms have not been reported. However, phosphorylation of tyrosine 14 on Cav-1α could modulate the interaction between Cav-1 and p190RhoGAP, and cell migration.(34,37) In earlier studies, antibodies that preferentially bind Cav-1α were used to detect secreted Cav-1; it is unclear whether Cav-1β was also secreted. If both Cav-1α and Cav-1β are secreted by prostate cancer cells, it would be ideal to develop a therapeutic approach that would have the ability to neutralize/remove both isoforms. Cav-1 scaffolding domain (CSD) is responsible for interaction between Cav-1 and various signaling molecules, including receptor tyrosine kinases, serine/threonine kinases/phosphatases, phospholipases, G protein-coupled receptors, and Src family kinases.(6,8) Therapeutic agents that bind CSD would have the potential to block Cav-1 functions. Indeed, in vitro studies showed that synthetic peptides carrying CSD sequences were able to block Cav-1-mediated signaling pathways and were proposed as a potential therapeutic agent to block Cav-1 functions.(29,38,39)
In this study, we focused on secreted Cav-1 as a potential therapeutic target and to raise antibodies that would have the ability to neutralize and/or remove Cav-1 secreted by prostate tumor. Anti-Cav-1 antibodies are commercially available. However, several reports indicated that polyclonal antibodies induced against full-length Cav-1 preferentially binds Cav-1α, suggesting that the first 31 amino acids at the N-terminus of Cav-1 carry an immunodominant epitope. Monoclonal antibodies that recognize both Cav-1α and Cav-1β are also available with unknown binding specificity. One antibody raised against Cav-1 C-terminal peptide showed low binding affinities and specificity (our unpublished data). So far, no antibody specific to CSD has been reported. Since Cav-1 can be secreted by normal pancreatic cells(30) and is highly conserved among mammals, we chose Cav-1 knockout mice (129sv/ C57BL6) as our host for antibody induction, so the potential Cav-1 reactive lymphocytes will be preserved in the state of self-tolerance. A panel of antigens for immunization were designed to cover different domains of Cav-1 but mainly focused on the CSD and surrounding sequences. These antigens were expressed and purified from Escherichia coli. A group of GST-fusion proteins carrying different domains of Cav-1 were also produced to conduct the epitope mapping of each antibody. We have successfully generated 11 hybridoma cell lines that produced anti-Cav-1 antibodies recognizing six different Cav-1 domains.
Materials and Methods
Cell lines and antibodies
P3×63Ag8.653, DU145, and LNCaP cells were obtained from ATCC (Manassas, VA) and maintained in complete RPMI1640 medium supplemented with 10% fetal bovine serum. Anti-Cav-1 monoclonal antibody clone 2297 was purchased from BD Bioscience (San Jose, CA). Condition medium were prepared from sub-confluent DU145 cells according to previously published protocol.(20)
Recombinant proteins
Full-length Cav-1 cDNA was generated by RT-PCR from DU145 total RNA extracts using the following primers: Cav-1-5′ (5′-ggatcccatatgtctgggggcaaatacg-3′); Cav-1-3′ (5′-aagcttatatttctttctgcaagt-3′) and then cloned between NdeI and HindIII sites of pRSET-A vector (Invitrogen, Carlsbad, CA), or BamH1 and EcoR1 of pGEX-2T vector (GE Healthcare). Truncated Cav-1 cDNAs were further generated by PCR using full-length Cav-1 cDNA as template. The following primers pairs were used: Cav-1-101: Cav-1-5′ and Cav-1-M-3′ (5′-aagcttacagcaagcggtaaaaccagtat-3′); Cav-1-CSD: Cav-1-sca-5′ and Cav-1-M-3′; Cav-1-CSD-C: csd-c-5′ (5′-ggatcccatatgattgcagaaccagaagggaca-3′) and Cav-1-3′. Each truncated Cav-1 cDNA was cloned between BamH1 and EcoR1 of pGEX-2T vector for GST-fusion protein expression.
Three antigens used for antibody induction (full-length Cav-1, GST-CSD, and GST-CSD-C) were expressed in transformed E. coli. Briefly, transformed BL21(DE3) cells were grown until OD600 reached 0.6 and followed by protein induction at 37°C for 3 h with 0.2 mM IPTG (Sigma-Aldrich, St. Louis, MO). Inclusion bodies were then purified following our previously published protocol.(40) For soluble GST-fusion protein production, transformed BL21(DE3) cells were grown at 37°C until OD600 reached 0.6, switched to 18°C for 1 h, and induced with 0.2 mM IPTG at 18°C overnight. The entire cell extracts were mixed with GSH-conjugated Sepharose beads (GE Healthcare, Piscataway, NJ) at 4°C for 2 h, washed with 20 mM Tris-HCl (pH 8.0) and 1 M NaCl, and proteins were eluted with 100 mM glycine (pH 3.0) and neutralized immediately with 1:10 volume of 1.5 M Tris-HCl (pH 8.8).
Hybridoma cell line production
All experiments involving animals were preapproved by IACUC of Scott & White Hospital (2010-023R, Temple, TX). The protocol for hybridoma cell line production was published previously.(41) Cav-1-/- mice (129sv/ C57BL6) were from Dr. Thompson's laboratory (MD Anderson Cancer Center, Houston, TX). Briefly, groups of three mice were subcutaneously injected with 50 μg of antigen in 80 μL of PBS and 20 μL of TiterMax adjuvant (Sigma-Aldrich) on day 1. At days 15 and 29, 50 μg of antigen in 100 μL of PBS were intraperitoneally injected to boost titer. Blood samples were collected at day 36 to evaluate antibody titer induced in each animal by ELISA (see below). The mouse with the highest antibody titer was given a final boost on day 57 with intravenous injection of 10 μg of soluble antigen. Splenocytes were harvested at day 60 and fused with P3×63Ag8.653 myeloma cells to generate hybridoma cell lines. Culture supernatant from each single clone was diluted 1:10 and tested in an ELISA. All positive clones were used for isotype identification with an Iso-Gold kit (BioAssay Works, Ijamsville, MD). The identified anti-Cav-1 antibody-producing hybridoma cell lines were expanded for further antibody purification.
Antibody purification
Hybridoma culture supernatant (200 mL) was mixed with 1 mL Protein G Sepharose beads (GE Healthcare, Piscataway, NJ) at 4°C overnight. Antibodies were eluted with 100 mM glycine pH 3.0 and neutralized immediately with 1/10 volume of 1.5 M Tris-HCl (pH 8.8). Desalting column PD-10 (GE Healthcare) was used to exchange the buffer of purified antibodies into PBS. Protein concentrations were measured by Bradford assays (Bio-Rad, Hercules, CA) with BSA (Pierce, Rockford, IL) as standards and verified by reduced SDS-PAGE.
cDNA synthesis
Approximately 5×106 cells of selected hybridomas were pelleted and washed with PBS for total RNA extraction using RNeasy kit (Qiagen, Valencia, CA). cDNA of variable regions were amplified by RT-PCR using mouse Ig-primer sets (EMD Bioscience, Gibbstown, NJ) and cloned into linear pDrive TA cloning vector (Qiagen). The PCR inserts were sequenced and correct light chain variable (VL) and heavy chain variable (VH) sequences were verified by comparing with the immunoglobulin sequences database (
Enzyme-linked immunosorbent assay
100 ng of GST-fusion proteins were added to each well of a glutathione (GSH)-coated 96-well plate and incubated at room temperature for 1 h. Primary antibodies were then added at room temperature for 1 h, followed by HRP-conjugated secondary antibodies (Jackson Lab, West Grove, PA) at room temperature for 45 min. The HRP activity was detected by HRP substrate (R&D Systems, Minneapolis, MN) for 10 min and stopped by 0.5 N H2SO4. OD450 was then read in a Versamax tunable microplate reader.
Polyacrylamide gel electrophoresis and immunoblot
Protein samples were mixed with SDS sample buffer (Invitrogen) with reducing agent, incubated in boiled water for 5 min, and loaded into a 12.5% or 15% polyacrylamide gel for the analysis of antibodies or Cav-1, respectively. The gels were then stained with Coomassie Blue R-250 solution (Bio-Rad) for 30 min. After destaining, images were taken by a gel documentation system and labeled by Photoshop (San Jose, CA). For Cav-1 immunoblots, separated proteins in polyacrylamide gels were transferred onto nitrocellulose membrane (Bio-Rad) with a semi-dry transfer apparatus (GE Healthcare). Anti-Cav-1 antibodies (0.1 μg/mL) were added to the membrane and incubated at room temperature for 1 h, followed by HRP-conjugated secondary antibodies (Jackson Lab) at room temperature for 45 min. The HRP activity was detected by enhanced chemiluminicent reagent (Pierce), and images were taken by a gel documentation system and labeled by Photoshop.
Cytosolic extraction
DU145 and LNCaP cells were harvested from sub-confluent culture. Soluble cytosolic extracts were prepared by resuspending cell pellet in 2x cell pellet volume of 1% NP-40 in PBS (Fisher Scientific) on ice for 30 min and centrifugation at 14,000 g for 10 min. Soluble supernatant was stored at −80°C.
Immunoprecipitation
1 mg of purified anti-Cav-1 antibodies 2A7 or 4C9 was each conjugated to 0.5 mL of CNBr-activated Sepharose beads (GE Healthcare) following the manufacturer's protocol. 5 μL of these antibody-conjugated beads were mixed with 1.5 mL of condition medium at 4°C overnight. Beads were spun down and resuspended in SDS sample buffer (Invitrogen) with reducing agent, and proteins were analyzed by immunoblots.
Binding competition assays
0.5 mg of purified 2A7, 3C12, or 4C9 was each conjugated with 0.5 mg of HRP (Pierce) following the manufacturer's protocol. In 96-well GSH-coated plates, 100 ng GST-Cav-1 were added into each well and incubated at room temperature for 1 h. Based on binding saturation curves (see Results below), either 3 nM (Group N1-31 and N32-80) or 10 nM (4D1 and 19G3) of unlabeled anti-Cav-1 antibodies were used to pre-block the plates at room temperature for 1 h. After being washed with PBS with 0.1% Tween-20, 1 nM HRP-conjugated anti-Cav-1 antibodies were added and incubated at room temperature for 1 h. HRP activities were detected by HRP substrates (R&D Systems) and measured at 450 nm absorption with a Versamax tunable microplate reader. The reading without any pre-blocking antibody was set as 100%, and the reading with pre-blocking by the same unlabeled antibody as the HRP-antibody was set as 0%.
Results and Discussion
Induction of anti-caveolin 1 antibodies
Three bacterially expressed antigens were used to inject Cav-1-knockout mice for antibody induction: full-length Cav-1, GST-CSD, and GST-CSD-C (Fig. 1A). Hybridoma cell lines were screened by ELISA coated with GST-Cav-1 (Fig. 1B), or with GST alone as a negative control. Positive clones were expanded for antibody production, purification, and characterization.
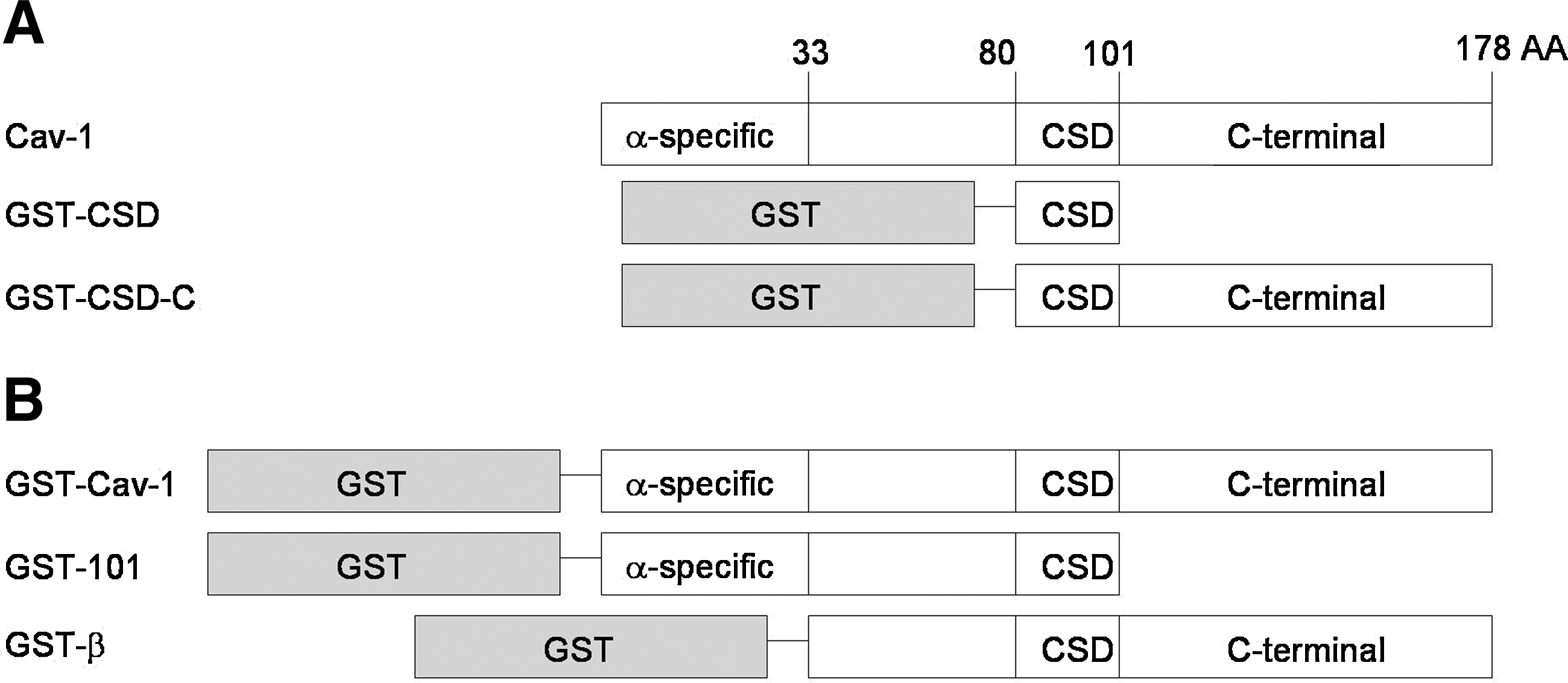
Recombinant Cav-1 proteins. (
Purified antibodies
Eleven positive clones were selected for further purification and analysis. Anti-Cav-1 antibodies were purified through binding to Protein G Sepharose beads (Fig. 2). A panel of GST fusion proteins carrying different domain of Cav-1 (Fig. 1) were used to map the binding domains of these antibodies by ELISA (Fig. 3). When full-length Cav-1 was used to immunize animals, all isolated antibodies recognize the N-terminal half of Cav-1 (Table 1). Five antibodies bind Cav-1α and the N-terminal 101 amino acid fragment but not Cav-1β (Fig. 3A), indicating a group of Cav-1α-specific antibodies. Three antibodies bind Cav-1α, Cav-1β, and N-terminal 101 amino acid fragment but not CSD (Fig. 3B), indicating a group of antibodies recognizing a part of Cav-1 between amino acids 32 and 80. Among the nine additional mice immunized with either GST-CSD or GST-CSD-C, only three positive clones were selected, suggesting CSD and the C-terminal half as poor antigens. 4D1 and 5C11 raised against GST-CSD bind Cav-1α, the N-terminal 101 amino-acid fragment, and CSD in ELISA (Fig. 3C), suggesting potential CSD-binding activities. 19G3 raised against GST-CSD-C binds Cav-1α and Cav-1β but not the N-terminal 101 amino acid fragment (Fig. 3C), suggesting a potential C-terminal domain binding activity. Based on binding domains, we separated these antibodies into four groups: N1-31 (2A7, 2H1, 3F5, 4E10, 4F10); N32-80 (3C12, 3F6, 4C9); CSD (4D1, 5C11); and Cav-1-C (19G3). The domain specificities of N1-32 and N33-80 groups also confirmed by immunoblots that N1-31 group only recognizes Cav-1α in DU145 cytosolic extracts and N32-80 group recognizes both Cav-1α and β (Fig. 4). 5C11 and 19G3 were not able to recognize denatured Cav-1 in immunoblot (data not shown), suggesting a possibility of recognizing Cav-1 structure domains. For unclear reasons, 4D1 recognized only Cav-1α but not Cav-1β in both ELISA and immunoblots.
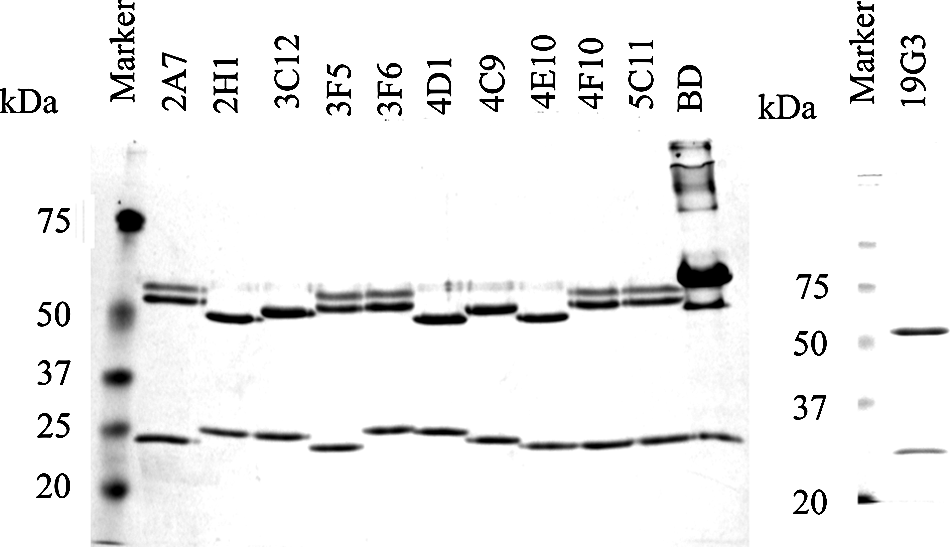
Coomassie R250-stained SDS-PAGE with 2 μg of purified anti-Cav-1 antibodies. Monoclonal antibody purchased from BD contains BSA.
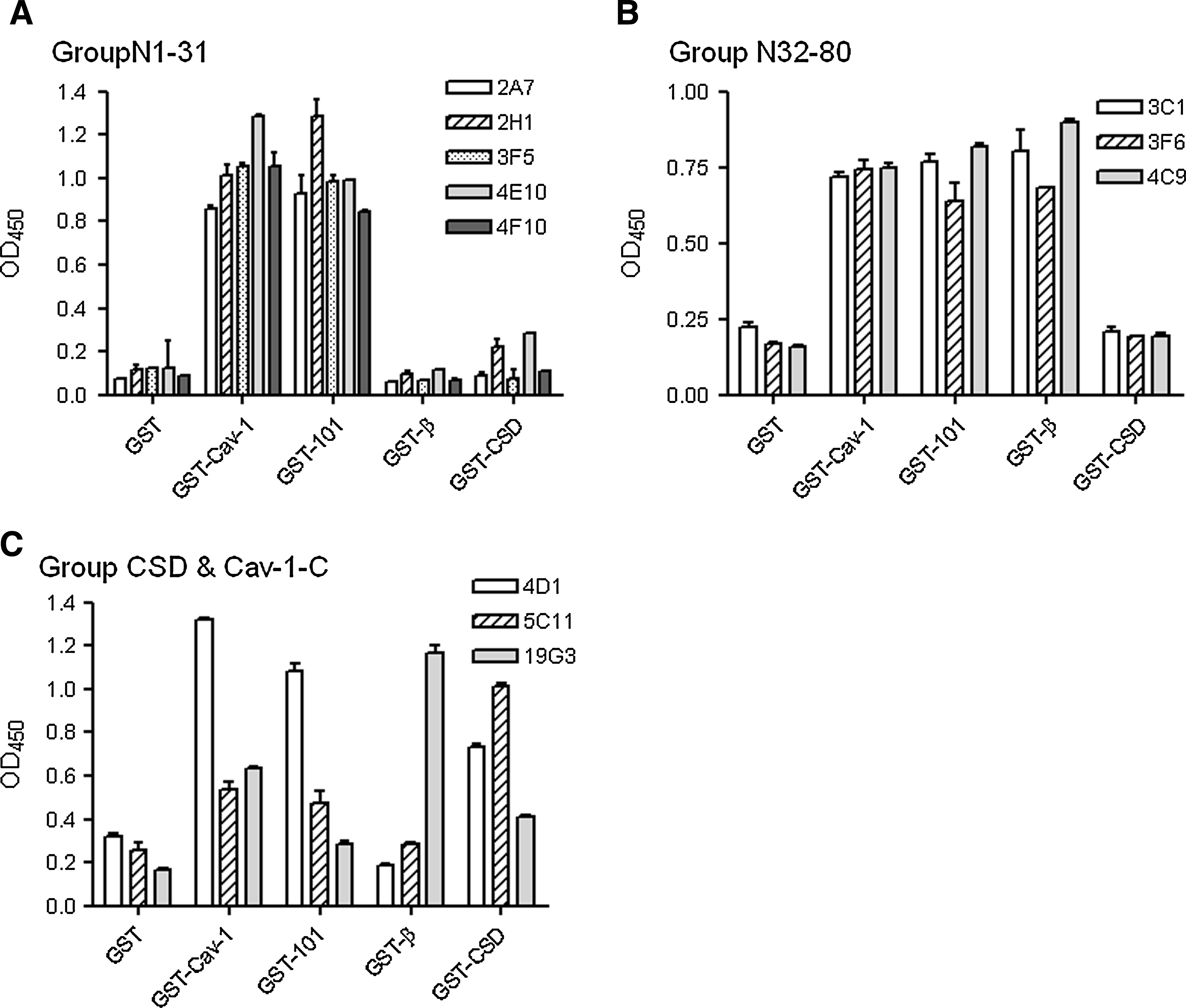
ELISA of purified antibodies against different Cav-1 domains. GSH-coated 96-well plates were incubated with different GST-fusion proteins as indicated on the x-axis. Anti-Cav-1 antibodies were separated into three epitope recognition groups: (
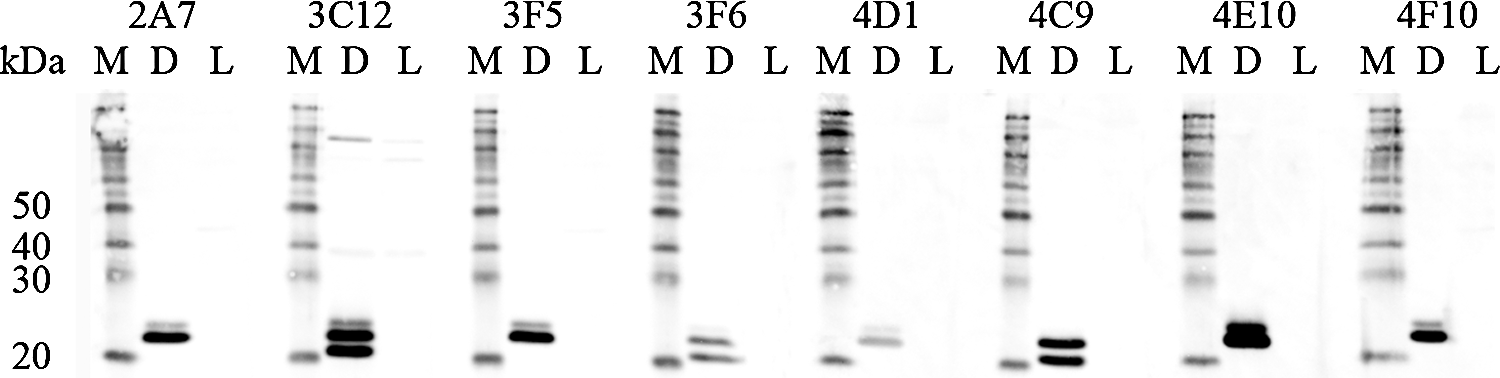
Anti-Cav-1 antibodies were tested in immunoblots with cytosolic extracts (20 μg total protein) from Cav-1+ DU145 (D) or Cav-1- LNCaP (L) cells. Biotin-conjugated protein markers (M) were used as migration indicator. The faster migrating band slightly above 20 kDa marker is Cav-1β.
Since we are selecting an antibody that can neutralize Cav-1 in human blood stream, antibody characterization was performed against GST-Cav-1 fusion proteins on GSH-coated ELISA plate, so the Cav-1 portion of the fusion protein is exposed to the aqueous phase. While antibodies in N1-31 and N32-80 groups show similar binding affinities (Kd at 10-50 pM), antibodies in CSD and CSD-C groups have much lower affinity to GST-Cav-1 (Fig. 5, Table 1). 5C11 is not able to bind all of the coated GST-Cav-1 and reach plateau at a lower level (Fig. 5C), suggesting that 5C11 can only recognize a sub-group of Cav-1. Due to the selected antigen recognition and the relatively lower Cav-1 binding affinities, 4D1, 5C11, and 19G3 were not considered for further development into therapeutic agents. However, 19G3 with its unique C-terminal binding activities might be used in the future to develop sandwich ELISA for Cav-1 detection.
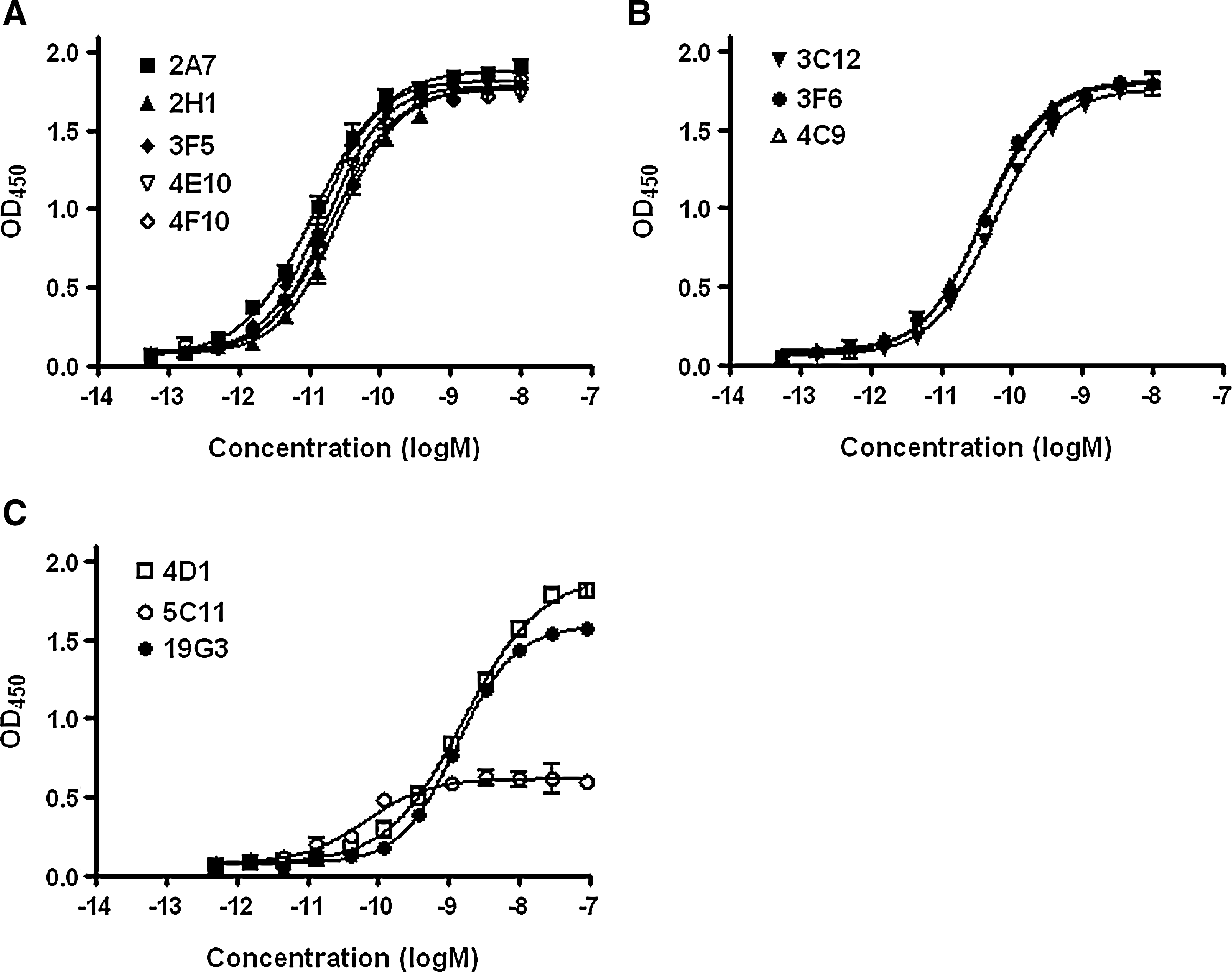
Binding affinity of purified anti-Cav-1 antibodies against immobilized GST-Cav-1 on GSH-coated plates. (
CDR mapping
Selected hybridoma cell lines were subjected to RT-PCR to isolate cDNA of the variable regions. Complementary determining regions (CDRs) were identified and compared (Table 2). All four analyzed sequences in N1-31 group have almost identical CDR sequences, and the overall variable region sequences are 91–99.5% identical (data not shown). This strongly suggests that they recognize the same Cav-1 epitope. Conversely, the three antibodies in N32-80 group showed lower levels of similarities among all the CDRs, suggesting the recognition of different epitopes within the amino acid 32-80 region. CDRs of 19G3 indicated the potential of yet another totally different binding domain.
Binding competition
To further define the Cav-1 binding domains of these antibodies, a binding competition ELISA was established. While all five antibodies in N1-31 group were able to block the binding of HRP-labeled 2A7 to GST-Cav-1, none of the antibodies in N32-80, CSD, or Cav-1-C group showed significant effect on 2A7 binding (Fig. 6A). It is interesting to find that 3F5 and 4E10 are able to block HRP-2A7 binding better than 2A7 itself, suggesting a lower off-rate of 3F5 and 4E10. It is also interesting to note that 4E10 has a higher degree of CDR sequence variation among antibodies in N1-31 group (Table 2). Since 4E10 has the potential to form a more stable complex with antigen, it would be an ideal candidate in the N1-31 group to be further developed.
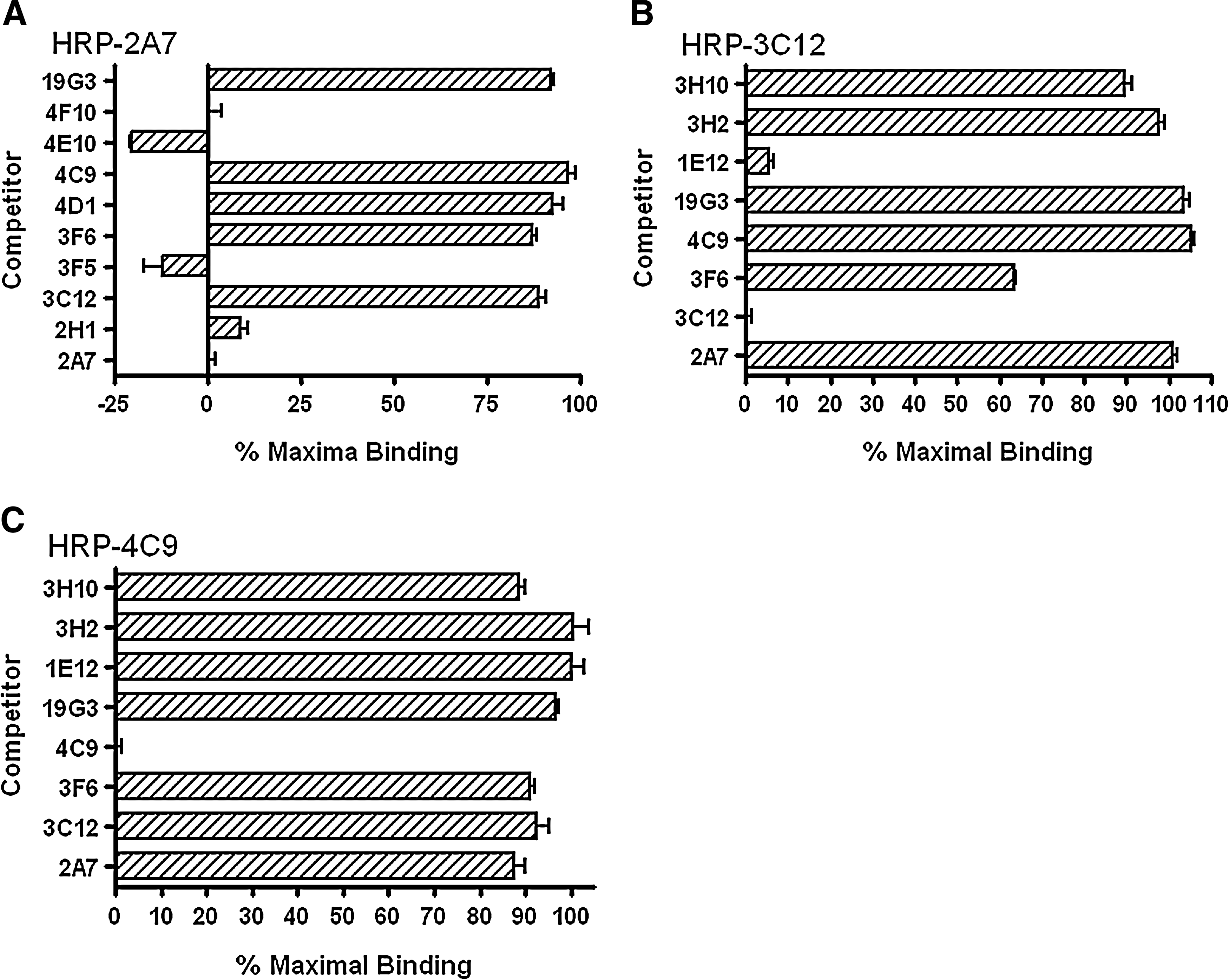
Binding competition assays. Purified (
Binding of HRP-labeled 3C12 to GST-Cav-1 can only be blocked by non-labeled 3C12 and partially blocked (∼40%) by 3F6, but not affected by the pre-blocking of 4C9 or other antibodies outside N32-80 group (Fig. 6B). This suggests that 3C12 and 3F6 recognize partially overlapped domains. Interestingly, binding of HRP-labeled 4C9 to GST-Cav-1 can only be blocked by 4C9 itself, but not by any other anti-Cav-1 antibodies (Fig. 6C). This suggests that 4C9 recognizes a unique Cav-1 domain. To narrow the domain within the amino acid 32 to 80 regions further, three monoclonal antibodies raised against synthetic Cav-1 peptides were used in binding competition assay. 3E2 and 3H10 were induced by peptide from amino acids 34 to 48, and 1E12 from 51 to 64. These three antibodies were produced by Dr. Thompson's group at MD Anderson Cancer Center (Tahir and Thompson, unpublished data). While 1E12 can block HRP-3C12 binding to a similar level of 3C12 itself, pre-blocking of 3E2 or 3H10 showed no effect (Fig. 6B). This would narrow the 3C12 binding domain down to amino acid 51-64 region. However, pre-blocking by 3E2, 3H10, or 1E12 had no effect on HRP-4C9 binding (Fig. 6C), suggesting that 4C9 binds amino acid 64-80 domain immediately outside the CSD. Since antibody that binds directly on CSD is not currently available, 4C9 with high specificity and affinity might be the best candidate to neutralize/sequester secreted Cav-1.
To summarize the domain mapping results (Fig. 7), the N-terminal 31 amino acids of Cav-1 carry a dominant antigen domain and all five Cav-1α-specific antibodies recognize that same domain with high affinities. Conversely, CSD and Cav-1 C-terminal half are not very immunogenic. The three antibodies in the N32-80 group are able to bind both Cav-1α and Cav-1β at relatively high affinities.

Anti-Cav-1 antibodies domain mapping.
Immunoprecipitation and co-immunoprecipitation
To select an antibody to be further developed into therapeutic agent, such antibody would have to be able to bind all the secreted Cav-1. The N1-31 group showed the highest binding affinities among all the anti-Cav-1 antibodies, ideal candidates to achieve highest efficacy. However, N1-31 group will not be able to recognize Cav-1β, if present. The possibility that Cav-1β is also secreted by prostate cancer cells has not been examined before. Traditionally, secreted Cav-1 was detected in serum-free condition medium by concentrating the medium by ∼100-fold and analyzing by immunoblots. In order to examine the possible presence of Cav-1β, condition medium from DU145 or LNCaP was first immunoprecipitated with 4C9-conjugated Sepharose beads to pull down both α and β forms, if present, and then analyzed by immunoblots with 4C9 antibody (IP-Western). Indeed, both α and β forms can be found in DU145 condition medium (Fig. 8A, lane 2) but neither was in low passage LNCaP medium (Fig. 8A, lane 3).

Immunoprecipitation and co-immunoprecipitation of Cav-1α and Cav-1β. (
It is known that secreted Cav-1 forms multimeric complexes.(30) To know whether both α and β forms of Cav-1 can be presented in the same complexes, co-immunoprecipitation experiments were performed with 2A7- and 4C9-conjugated Sepharose beads to pull down Cav-1 from DU145 condition medium. The bound Cav-1 proteins were analyzed by SDS-PAGE and immunoblot with 4C9 antibody. As shown in Figure 8B, both 2A7 (lane 4) and 4C9 (lane 5) can pull down both α and β forms at a similar ratio. This suggests that both forms of Cav-1 are present in the same complexes, and antibodies in both N1-31 and N32-80 groups would have the same potential to deplete all secreted Cav-1.
Conclusion
The autocrine and paracrine activities of secreted Cav-1 in prostate cancer have been shown to be ideal therapeutic targets. The ability of polyclonal anti-Cav-1 antibodies to suppress prostate cancer cell growth in culture and in mouse models has encouraged the development of antibody-based therapeutic agents. In this study, new monoclonal antibodies were raised against Cav-1 that will be further developed into anti-prostate cancer therapeutic agents. As summarized in Figure 7, we generated 11 monoclonal antibodies that recognize six different Cav-1 epitopes. Additional in vitro assays and animal model studies will be done to directly compare the Cav-1-neutralizing activities of a few selected anti-Cav-1 antibodies. The most effective antibody will then be subjected to humanization, retested in functional assays, and produced in larger quantities for pre-clinical and clinical studies.
Footnotes
Acknowledgments
This work was supported in part by Scott & White Startup Fund (JSL). Work done in MD Anderson Cancer Center (TC Thompson's lab) is supported by NIH/NCI R01 CA68814.
Author Disclosure Statement
TC Thompson holds the patent for therapeutic use of anti-Cav-1 agents. The other authors have no financial interests to disclose.