
Abstract
Monoclonal antibody (MAb) against trans-resveratrol (t-RSV) was obtained from hybridoma clones constructed from splenocytes of BALB/c mice immunized with carrier proteins (bovine serum albumin [BSA] and ovalbumin [OVA]) coupled with synthetic hapten mimicking t-RSV structure. The t-RSV-BSA derivate was more efficient at induction of the immune response than t-RSV-OVA. However, the use of t-RSV-OVA was advantageous during selection of hybridoma clones constructed from splenocytes of t-RSV-BSA-immunized mice. Pre-incubation of immune serum with free t-RSV inhibited the binding of antibody to t-RSV-BSA conjugate, suggesting the specific nature of antibody binding to t-RSV. Splenocytes obtained from the mouse immunized with t-RSV-BSA were used for the hybridoma construction. Expansion of the primary clones, their subsequent screening, and subcloning narrowed our search to allowed isolation of two IgG1a-producing hybridomas designated as 2H9 and 1B1. According to an indirect ELISA assay, the resulting MAb 2H9 had little or no cross-reactivity to cis-RSV. No recognition of trans-RSV-3-O-glucuronide and trans-RSV-3-sulfate was detected. The newly generated MAb against t-RSV may provide a highly valuable and cost-effective tool for the analytical assay for t-RSV in different biological and agricultural specimens.
Introduction
Since most of the biological effects of RSV develop in a dose-dependent manner, RSV quantification in biological specimens has become extremely important in modern laboratory practice. The importance of RSV quantification protocols is equally obvious for the food industry, given that RSV content in food products may become an extremely powerful marketing tool. High performance liquid chromatography (HPLC), overpressured layer chromatography (OPLC) with further fluorescent, or mass spectrometry detection of RSV isomers are among the widely accepted protocols for RSV quantification.(8,9) Moreover a rapid spectrofluorimetric method has recently been proposed to satisfy the growing demand for monitoring of RSV concentration in wine samples.(10)
Here we report a monoclonal antibody against RSV, which could be possibly used for rapid immunoassay procedures to quantify RSV content in different agricultural and biological specimens.
Materials and Methods
trans-Resveratrol (t-RSV) was obtained from Kaden Biochemicals (#621309, Hamburg, Germany). Bovine serum albumin was purchased from Amresco (#0332; Solon, Ohio). Ovalbumin (OV) and Freund's adjuvant were from Sigma-Aldrich (#A5503 and F5881; St. Louis, MO). ELISA plates were purchased from Greiner Bio-One (#762070; Frickenhausen, Germany). Liquid Substrate System for ELISA reagents was obtained from Sigma-Aldrich (#T0440). Female BALB/c mice aged 3 months were ordered from Puschino Breeding Facility (Moscow, Russia).
All other chemicals, solvents and supplies were purchased from Sigma-Aldrich, unless mentioned otherwise. All aqueous solutions including HPLC buffers were prepared using Millipore Milli Q grade water. Wine varieties were purchased from a general grocery supplier in Cambridge, United Kingdom.
Antigen generation
All glassware was oven- or flame-dried for 10 h and cooled in a nitrogen atmosphere. Antigen preparation required chemical modification of t-RSV, which was performed according to a modified protocol of Jiang.(11)
Hapten synthesis
Briefly, the introduction of a carboxyl group into the t-RSV structure required for the preparation of a t-RSV-OVA and t-RSV-BSA conjugates(11) was performed by selective O-alkylation of the hydroxyl group at the 4’-position with ethyl-6-bromohexanoate (Fig. 1). The resulting derivate (2) was isolated and fully characterized by NMR analysis. Next, a saponification of derivate 2 was accomplished with potassium hydroxide solution in methanol to obtain free acid 3, which was used for coupling with carrier proteins (BSA and OVA). The identity of derivates was confirmed with NMR on a Bruker AV-300 spectrometer (Figs. 2, 3). A brief protocol for hapten generation and the t-RSV-coupling procedure are presented below.
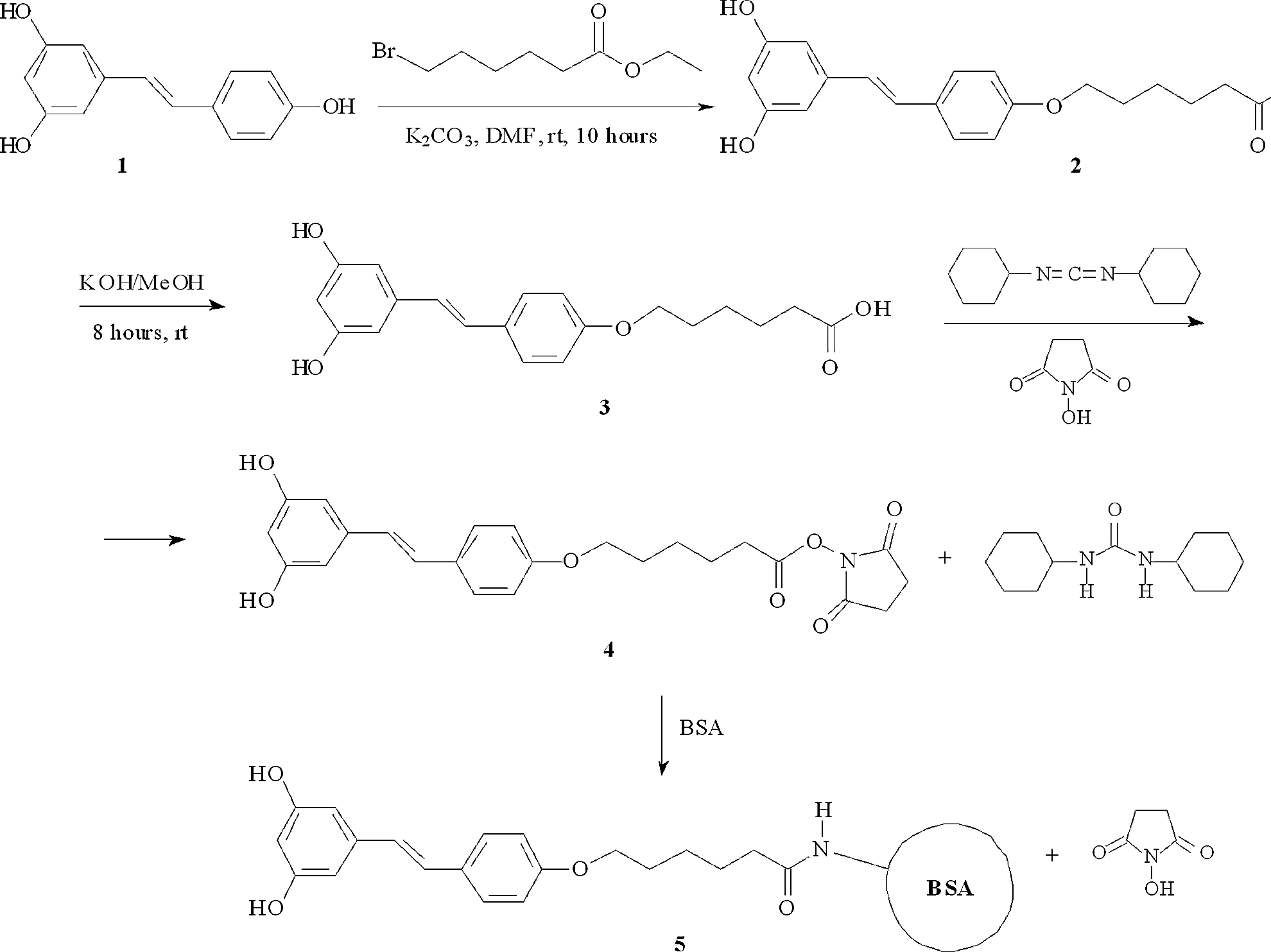
t-RSV hapten synthesis scheme (compounds 2–5).
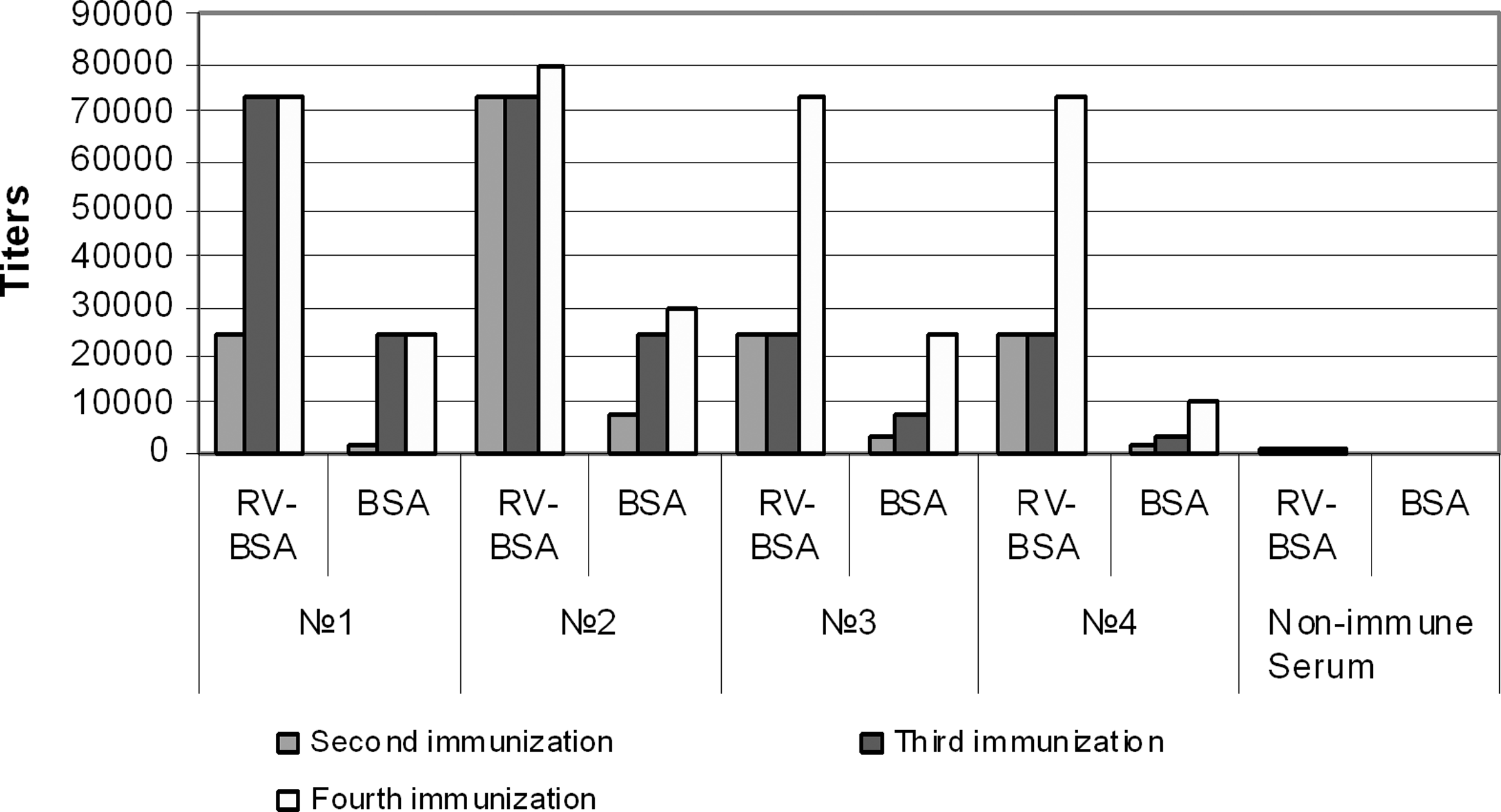
Immune response in BALB/c mice injected with t-RSV-BSA conjugate. BALB/c mice were injected with t-RSV-BSA conjugates and immune serum specimens were obtained after second, third, and fourth rounds of immunization. Antibody titers to t-RSV–BSA conjugate or BSA alone were compared in four (1–4) individual mice. Serum from non-immunized mice served as control.
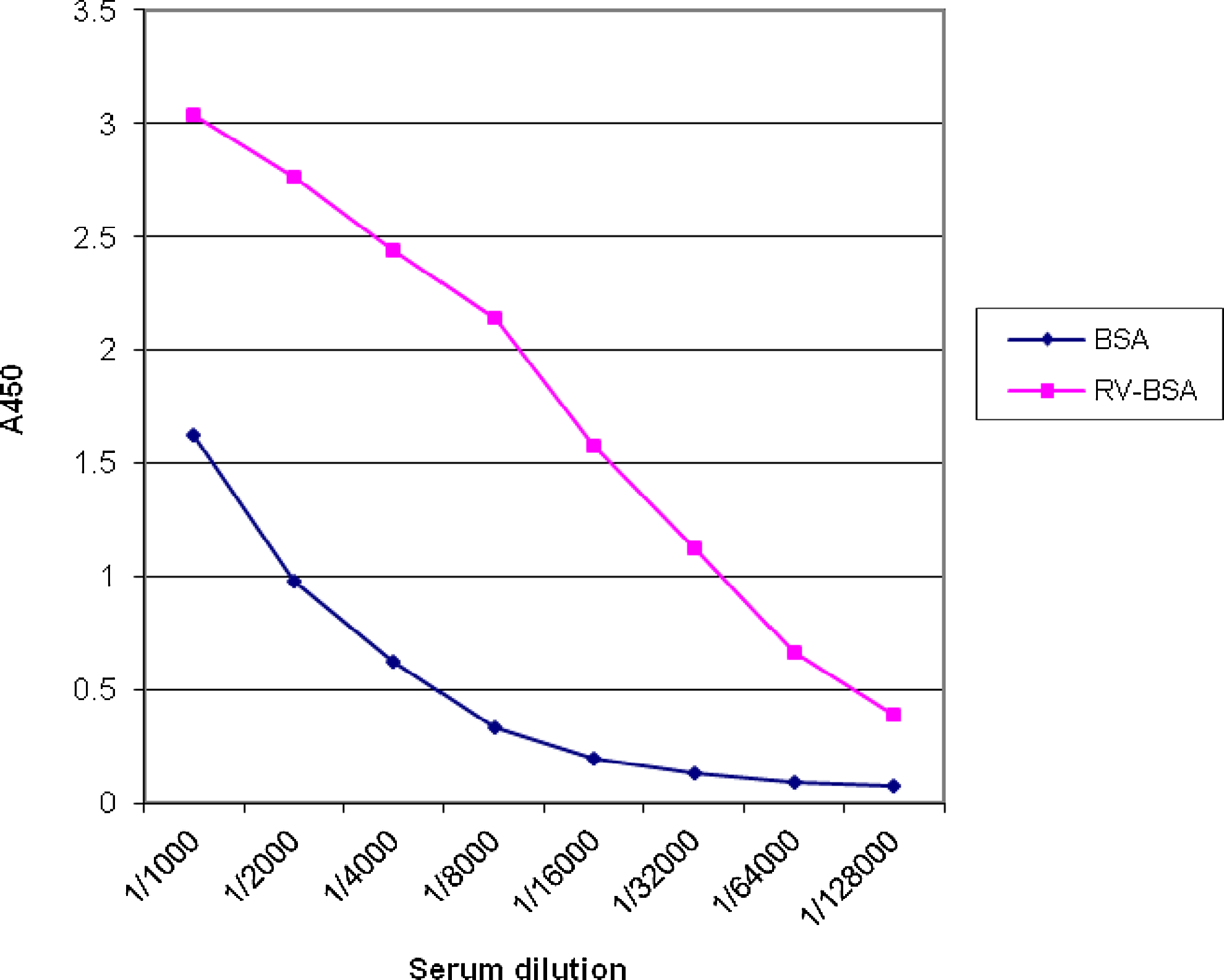
Recognition of t-RSV-BSA conjugate versus BSA alone by immune serum obtained from mouse 1. BALB/c mice were immunized and serum specimens were obtained after fourth round of immunization. Serial dilutions of the serum from mouse 1 were analyzed in the direct ELISA with t-RSV-BSA conjugate and BSA alone.
O-alkylation
K2CO3 (1.33 g, 9.64 mmol, 1.1 equiv) and ethyl-6-bromohexanoane (2.93 g, 13.14 mmol, 1.5 equiv) were added under argon to a solution of resveratrol (1) (2.00 g, 8.76 mmol) in DMF (10 mL). After stirring overnight, the mixture was diluted in EtOAc (100 mL) and washed with 1 N HCl (3× 50 mL). The organic layer was dried over MgSO4 and filtered. The solvent was evaporated under reduced pressure and the residue was purified by flash chromatography using CH2Cl2/EtOAc 10:1.5 as eluent to afford 0.4 g of 2 (30%).
Rf value, 0.71 (acetone/hexane, 50:50, v/v). Mp 182–183°C. 1H NMR (DMSO-d6, ppm) δ 1.21 (t, 3H, J=7.4 Hz, CH3); 1.47 (m, 2H, CH2); 1.63 (m, 2H, CH2); 1.77 (m, 2H, CH2); 2.35 (t, 3H, J=7.2 Hz, CH3); 4.00 (t, 2H, J=6.4 Hz, CH2); 4.09 (k, 2H, J=7.4 Hz, OCH2); 6.16 (t, 1H, J=2.2 Hz, Ar–H); 6.42 (d, 2H, J=2.2 Hz, Ar–H); 6.92 (m, 4H, Ar–H); 7.56 (d, 2H, J=8.4 Hz, Ar–H); 9.28 (s, 2H, OH).
Saponification
Compound 2 (0.2 g) was dissolved in a 5% solution of KOH in methanol (10 mL). The resulting mixture was stirred for 12 h at room temperature. The mixture was extracted with ethyl acetate twice (2×10 mL) to remove starting material and then the water phase was acidified to pH 2.0 using 6 N HCl and extracted with ethyl acetate (2×10 mL) again. The organic phase was evaporated on a rotary evaporator, and the residue was purified with column chromatography on silica gel using a mixed solvent (acetone/petroleum ether with a gradient of 1:2–1:1) to produce compound 5 (0.142 g, 62%).
Rf value, 0.82 (HOAc/EtOAc=5/95). Mp 196–197°C. 1H NMR (CD3OD, ppm) δ 1.52 (m, 2H, CH2); 1.65 (m, 2H, CH2); 1.75 (m, 2H, CH2); 2.34 (t, 3H, J=7.2 Hz, CH3); 3.98 (t, 2H, J=6.2 Hz, CH2); 6.17 (t, 1H, J=2.2 Hz, Ar–H); 6.45 (d, 2H, J=2.2 Hz, Ar–H); 6.91 (m, 3H, Ar–H); 6.94 (d, 2H, J=16.1 Hz, Ar–H); 7.43 (d, 2H, J=8.8 Hz, Ar–H). Active ester 4 was synthesized by the treatment of free acid 3 (100 mg) with dicyclocarbodiimide (DCC, 78 mg) and N-hydroxysuccinimide (80 mg) in 1.5 mL of DMF at room temperature. The precipitate (dicyclohexyl urea) was filtered out and the mother solution was used for the conjugate synthesis.
Conjugation
The final conjugate 5 was obtained by adding compound 4 (100 mg) in dimethylformamide to an aqueous solution (0.1 M NaHCO3) of bovine serum albumin (250 mg) with further incubation of the resulting mixture (t-RSV-BSA) at 4°C for 10 h. In the same manner the second conjugate was prepared starting from 100 mg of 3 and 250 mg of ovalbumin (t-RSV-OVA). Conjugation products were dialyzed for 28 h against 0.02 M NH4HCO3 with a further switch to 0.01 M NH4HCO3. Dialyzed solutions were centrifuged at 10.000 g and passed through a Sephadex G15 column balanced with 0.01 M NH4HCO3. Final products were lyophilized and kept under nitrogen. The efficiency of conjugation was measured and the resulting t-RSV conjugates were characterized using fluorescent spectroscopy as described by others.(12) It was assumed from the beginning and confirmed once an ELISA had been developed that t-RSV conjugates are stable for at least 1 month when stored at −20°C.
Standard solutions
A stock solution of t-RSV was made in 50% methanol and water and kept in small aliquots at −80°C in darkness. The frozen solution was thawed by 1 h exposure to room temperature. Unused portions of aliquots were discarded. Working solutions of t-RSV were made by dissolving in PBST supplemented with 10% ethanol. For the ELISA protocol, 0.5% BSA was added to the working solutions. ELISA calibration specimens were made by spiking PBST-ethanol with increasing amounts of t-RSV within 2 h of the scheduled assay.
Antibody generation
All animal procedures were approved by the local Animal Research Ethical Committee at Moscow Institute of Virology. Solutions of t-RSV conjugates (t-RSV-OVA and t-RSV-BSA) for animal use were prepared in 0.9% NaCl on the day of injection. Mice were immunized three times with a 3-week interval between the first and second injections of conjugates and a 2-week interval between the second and third injections of t-RSV conjugates. The first immunization was performed by intraperitoneal injection of 100 μg of t-RSV-BSA or t-RSV-OVA conjugate with complete Freund's adjuvant (1:1 vol/vol). The second immunization was done subcutaneously by injection of 200 μg of t-RSV conjugate with Freund's incomplete adjuvant. 100 μg of t-RSV conjugates with Freund's incomplete adjuvant were used for the third immunization. Immunized mice were boosted intraperitoneally with 200 μg of t-RSV conjugates with no adjuvant added 2 weeks after the third immunization.
Sera evaluation
Serum specimens obtained from the immunized mice were routinely screened using indirect ELISA starting from the second round of immunization.
Titer check
Greiner Bio-One 96-well ELISA microplates were coated at room temperature for 2 h and at 4°C overnight, with 100 μL t-RSV-BSA, t-RSV-OVA, or BSA and OVA alone (5 μg/mL in 0.1 M carbonate buffer, pH 9.5). Plates were washed 3 times with PBST. Control sera from non-immunized mice and sera from immunized mice were diluted with PBST supplemented with 1 mg/mL BSA. 100 μL serial dilutions ranging from 1:1000 to 1:128,000 were pipetted into the wells of the ELISA plates, with further incubation of the plates at 37°C for 1 h. After three washings with PBST, addition of goat peroxidase-labeled antibody against mouse IgG was made. Further incubation of the plates at 37°C for 1 h was followed by three PBST washes and addition of 100 μL/well of tetramethylbenzidine (TMB) chromogenic substrate. After 30-min incubation at room temperature, the color reaction was stopped by adding 50 μL/well 1 N H2SO4. The plates were read at 450 nm (Multiscan EX, Thermo Scientific, Waltham, MA). The titers were measured for each individual mouse after every immunization.
Hybridoma construction
After 72 h following the final intravenous boost, the spleens of anesthetized mice (positive responders) were removed and dispersed and the resulting splenocytes were fused with Sp-2 cells and incubated in HAT medium according to the conventional protocol.(13) Positive wells were inspected microscopically for cluster formation and the supernatants were tested for antibody presence by indirect ELISA. Positive clones were subcloned by limiting dilution protocol using spleen feeders. Antibodies were isotyped with a mouse monoclonal antibody isotyping reagent.
Specificity evaluation
An indirect competitive ELISA assay was used to evaluate the specificity of immune response in the mice, as well as immunoglobulins produced by hybridoma clones. ELISA plates were coated with each t-RSV conjugate or each carrier protein as described above. Serial dilutions of the competitor ranging from 15 ng/mL to 1000 ng/mL were made from a stock solution of t-RSV (1 mg/mL) in PBST supplemented with 0.5% BSA and 10% ethanol. 100 μL of the competitor serial dilution and an aliquot of immune serum were inoculated into each well of the microplate. The amount of sera introduced into the wells was constant and corresponded to the serum dilution that produced an OD450 reading of ∼1.0, as determined by the preliminary titer evaluation. The competitor and primary antibody were co-incubated for 1 h at 37°C with occasional gentle shaking of the plate. The further processing of the plates included washing and addition of TMB as described above. A competition assay for each serum specimen was done in duplicate. The readings were used to calculate inhibition value (CInh) according to:
where A450 + t-RSV is an optical density at 450 nm in the wells with t-RSV added versus the well with no addition of t-RSV (A450 – t-RSV). Similar format of indirect competitive ELISA was used for the evaluation of IgG produced by hybridoma clones.
Standard curve generation
t-RSV-BSA or t-RSV-OVA were used as coating agents for an indirect competitive ELISA as described above. Serum specimens obtained from immunized mice were used as a source of primary antibody. Increasing concentrations of non-conjugated t-RSV mixed with aliquots of sera were pre-incubated on ice for 1 h before inoculation into the wells of the ELISA microplate. All ELISA reactions for standard curve generation were performed in duplicate and were repeated three times. The mean values for the calibration curve within the linear range were fitted using linear regression analysis.
Results
Using the protocols described above we were able to conduct hapten synthesis according to the scheme shown in Figure 1 and confirm the identity of the intermediates (ester 2 and acid 3) from 1H NMR spectra in CDCl3. Free acid 3 was used for a coupling procedure with carrier proteins. The resulting conjugates were used for the immunization protocol.
Injections of t-RSV conjugates into BALB/c mice produced a detectable immune response from the second round of immunizations. All mice injected with t-RSV-BSA had measurable antibody in serum to the immunogen (Fig. 2), while only three out of seven mice injected with t-RSV-OVA had some antibody response to the conjugate (results not shown). Over several attempts t-RSV-BSA was a better inducer of the immune response than t-RSV-OVA. Moreover, Figure 2 shows that all immune serum specimens obtained from the mice immunized with t-RSV-BSA demonstrated better recognition of the conjugate than BSA alone. This may indicate that immunization with t-RSV-BSA resulted in the production of antibody against t-RSV and the linkage zone of the conjugate, allowing partial recognition of BSA.
The recognition pattern of BSA versus t-RSV-BSA by immune serum obtained from mouse 1 is presented in Figure 3. It reveals that recognition of t-RSV-BSA is much stronger than BSA alone.
Competition analysis confirmed the specific nature of antibody binding to t-RSV. As can be seen in Figure 4, pre-incubation of immune serum with free t-RSV significantly depleted binding of antibody to t-RSV-BSA conjugate. Similar analysis was done with the serum specimens obtained from two mice immunized with t-RSV-OVA. In both cases, the addition of free t-RSV did not affect binding of the antibodies with the immobilized conjugate (results not shown). In other words, the serum specimens from the mice immunized with t-RSV-OVA did not contain antibodies specific to free t-RSV. Therefore our attempts to use t-RSV-OVA in the immunization protocol were terminated.
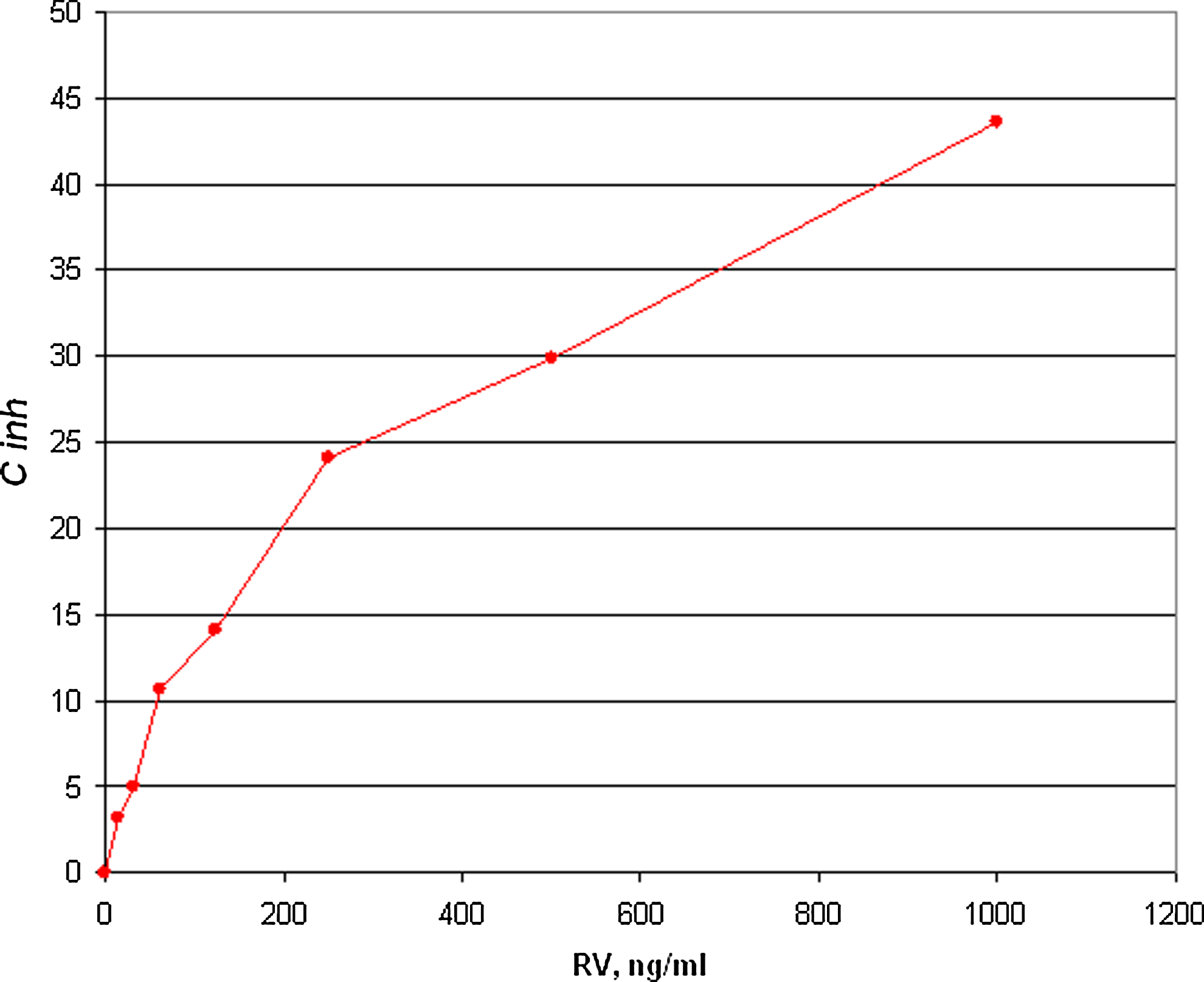
Effect of t-RSV on t-RSV-BSA conjugate recognition by immune serum from mouse 1. BALB/c mice were immunized and serum specimens were obtained after fourth round of immunization. An indirect competitive ELISA was performed.
Splenocytes obtained from mouse 1 injected with t-RSV-BSA were used for the hybridoma construction. 767 primary hybridoma clones were established and tested by indirect ELISA with t-RSV-BSA and BSA alone. Expansion of the clones and their subsequent screening narrowed our work to 16 independent clones that were capable of recognition of t-RSV-BSA with little or no cross-reactivity to BSA alone. Further expansion of the clones limited our selection to eight cell lines, which were stable enough to maintain antibody production. Finally, the cloning protocol employed allowed us to narrow our work to two IgG1a-producing hybridomas designated as 2H9 and 1B1 (Fig. 5).
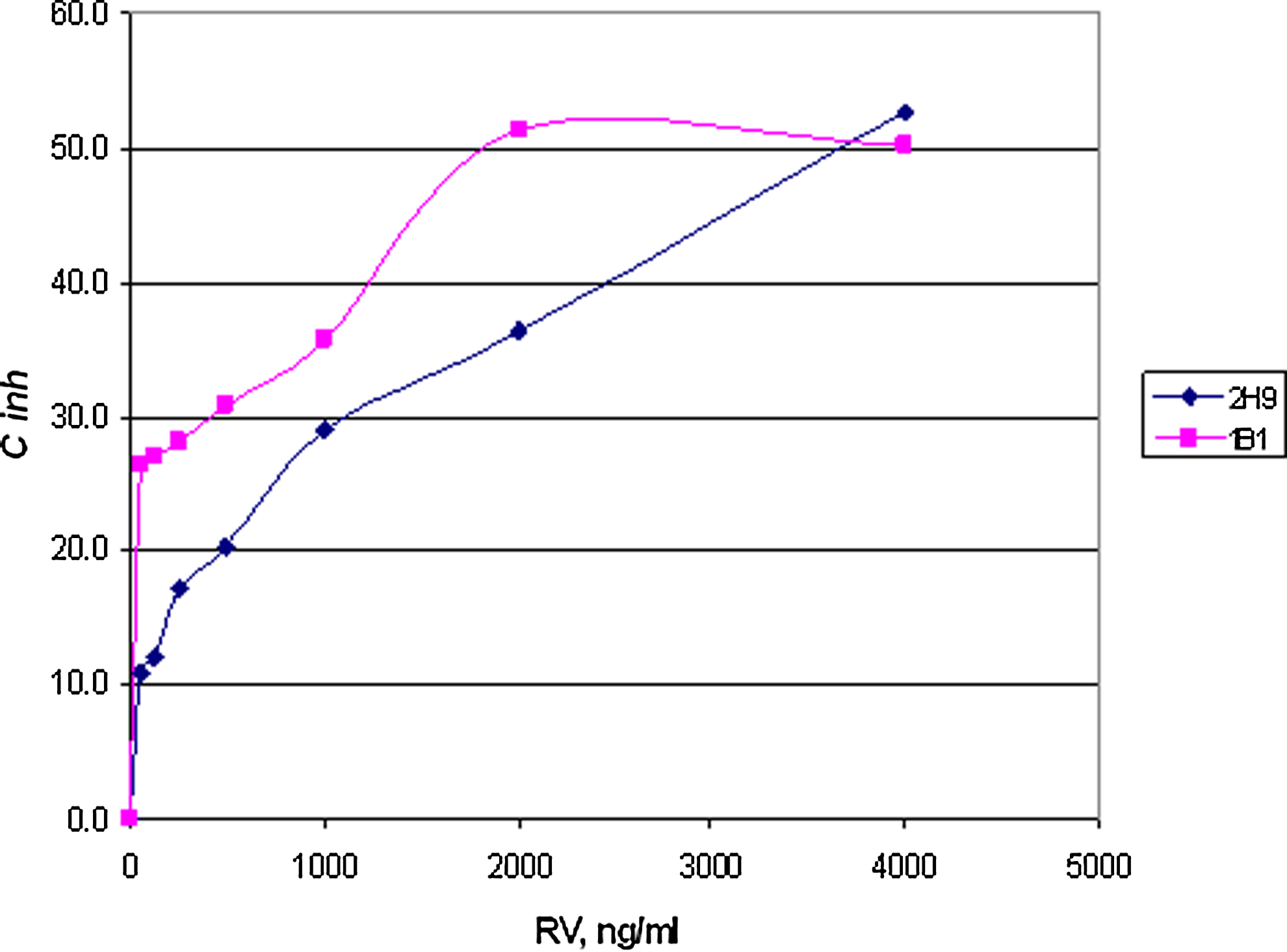
Effect of t-RSV on t-RSV-BSA conjugate recognition by MAbs 2H9 and 1B1 in the direct ELISA with t-RSV-BSA. Hybridoma clones were constructed using splenocytes from mouse 1. Resulting MAbs 2H9 and 1B1 were tested in an indirect competitive ELISA with t-RSV-BSA conjugate.
The newly generated MAbs were further tested in an indirect ELISA assay in which MAb 2H9 displayed better consistency and reproducibility on multiple occasions. MAb 2H9 had little or no cross-reactivity to cis-RSV. No recognition of trans-RSV-3-O-glucuronide and trans-RSV-3-sulfate was detected.
Discussion
The majority of analytical methods developed for measurement of low-molecular weight food constituents are based on extremely sophisticated instrumental techniques such as gas chromatography and high pressure liquid chromatography coupled with mass spectroscopy. These methods represent a “gold standard” for many analytical protocols, including RSV analytical assays.(15) Besides the need for expensive equipment and reagents, these methods are time-consuming and require highly skilled support personnel, as well as specialized laboratory space. On the other hand, immunological methods, in particular ELISA, represent a newly emerging alternative to many conventional analytical protocols. Indeed, recent advances in immunology allow the generation of highly specific detection molecules (antibodies) against virtually any substance.(16)
In the present article, we report a successful hapten synthesis, preparation of a hapten-protein conjugate, and generation of monoclonal antibody against t-RSV, a polyphenolic compound originating in grapes and berries. The newly generated MAbs can be used for the development of a novel immunoassay that can be employed for the detection of t-RSV in different biological and agricultural specimens. t-RSV-BSA conjugate gave the best performance in the immunization protocol while t-RSV-OVA complex was less immunogenic. To our knowledge, the generation of MAb to t-RSV has never been reported before. However, the probability of a positive outcome to our study was quite high, given recent reports regarding the immunogenic properties of stilbenes. It has been reported that a close chemical analog of t-RSV, t-stilbene can be effectively used to elicit a panel of monoclonal antibodies.(17) The newly generated MAb against t-RSV may provide a highly valuable tool for the development of analytical immunoassay for t-RSV.