
Abstract
VP7 is a major group-specific protein of the bluetongue virus (BTV), and is therefore a candidate for use as a diagnostic reagent. In this study, BALB/c mice were immunized with BTV16, and the lymphocyte hybridoma technique and indirect ELISA screening method were employed to obtain two strains of hybridoma cells secreting specific monoclonal antibodies (MAbs) to BTV16. Eukaryotic recombinant plasmids coding for 10 segments of BTV16 separately were transfected into BHK-21 cells, respectively, followed by immunofluorescence, showing that two MAbs only reacted with BTV-VP7. Western blot analysis showed the same result. Indirect immunofluorescence results indicated that two of the MAbs present different response spectrums with BTV1∼24 serotypes. These results indicate that these MAbs may be good candidates for a specific diagnostic method and functional exploration of the VP7 protein.
Introduction
BTV is an icosahedral, non-enveloped virus and its genome consists of 10 double-stranded RNA (dsRNA) segments that code for 11 distinct viral proteins: seven structural proteins and four non-structural proteins.(6,7) Among the BTV proteins, VP7, which comprises approximately 36% of the total content of the virion, is the major group-specific protein, and is therefore a candidate for use as a diagnostic reagent.(8,9)
Here, we describe the generation and characterization of two monoclonal antibodies against VP7 protein, one of which can specifically recognize 24 serotypes of BTVs and would be useful in the detection of BTV.
Materials and Methods
Animals, recombinant plasmids, cells, and virus
Six-week-old BALB/c mice were supplied by the Centre of Experimental Animals (Harbin Veterinary Research Institute, Chinese Academy of Agricultural Sciences). Eukaryotic recombinant pCI-neo plasmids (namely, pCI-neo-BT16-VP1, pCI-neo-BT16-VP2, pCI-neo-BT16-VP3, pCI-neo-BT16-VP4, pCI-neo-BT16-VP5, pCI-neo-BT16-VP6, pCI-neo-BT16-VP7, pCI-neo-BT16-NS1, pCI-neo-BT16-NS2, pCI-neo-BT16-NS3, respectively) were conserved in our laboratory. The BHK-21 cells and the myeloma cell line SP2/0 were cultured in Dulbecco's modified Eagle's medium (Invitrogen, Carlsbad, CA) in humidified 5% CO2 atmosphere at 37°C. All cell culture media were supplemented with 10%, 56°C, 30 min heat-inactivated fetal bovine serum (HyClone, Victoria, Australia) and antibiotics (0.1 mg/mL streptomycin and 100 IU/mL penicillin). BTV1∼24, Ibaraki virus (IBAV), chuzan virus (CV), and akabane virus (AKAV) were kept in our laboratory.
Virus propagation and TCID50 assay
BTV16 virus was prepared by infecting BHK-21cells in DMEM at 37°C and 5% CO2. When 80–90% of the cells appeared CPE, the culture mixture was subjected to three rapid freeze/thaw cycles, letting virus release from the cells. Then the virus-cell mixture was centrifuged at 12,000g/min for 10 min at 4°C. The supernatant was collected and stored in −80°C, which was used as immunogen and coating antigen in the following ELISA. Meanwhile, TCID50 assay was carried out to determine viral titer of BTV16.
Immunization of mice and generation of monoclonal antibody
Four female BALB/c mice were immunized by four injections of 0.6 mL BTV16 virus-infected cell supernatant per mouse at 14-day intervals. The mice were bled and tested for serum antibody titers to BTV16 by ELISA. The mouse with the highest serum antibody titers was euthanized and its spleen collected. B lymphocytes were fused with mouse myeloma cells, according to the method of Kohler and Milstein.(10) The resulting hybridoma cells were plated in 96-well plates and cultured in HAT selection medium (DMEM containing 20% fetal bovine serum, 100 μg/mL streptomycin, 100 IU/mL penicillin, 100 mM hypoxanthine, 16 mM thymidine, and 400 mM aminopterin). The hybridoma supernatants were screened by an indirect ELISA method against BTV16. Positive clones were subcloned and rescreened.
ELISA
In preliminary experiments, checkerboard titrations were performed to find the optimum concentration of coating antigen (60°C, 30 min heat-inactivated BTV16 virus-infected cell supernatant) and detect antibodies. Then, inactivated BTV16 virus-infected cell supernatant and BHK-21 cells (negative control) were diluted in phosphate-buffered saline, respectively, and 100 μL were added to each well of 96-well cell culture plates (4°C overnight).
Plates were washed with PBS (pH 7.4) with 0.1% Tween-20 between steps. 5% skim milk diluent was used to prevent non-specific binding (2 h, 37°C) and also as a diluent for the detection of antibodies. The hybridoma supernatants were incubated for 1 h at 37°C. After being washed three times, the plates were incubated for 1 h at 37°C with peroxidase-conjugated goat anti-mouse IgG (H+L) antibody (ZSGB-BIO, Beijing, China). After washing three times, immunoreactivity was visualized by means of adding substrate solution containing o-phenylenediamine (OPD). Reaction was stopped with 2 M H2SO4.(11)
Subtype identification of monoclonal antibodies
The subtype of the monoclonal antibodies (MAbs) was identified using a mouse monoclonal antibody isotyping kit (Invitrogen) according to the manufacturer's instructions.
Transfection of eukaryotic recombinant plasmids and immunofluorescence assay
Ten eukaryotic recombinant plasmids (pCI-neo), which contain one of 10 gene segments of BTV16, respectively, were transfected into BHK-21 cells according to the manufacturer's instructions for Lipofectamine™ 2000 Reagent (Invitrogen, Carlsbad, CA). Meanwhile, a plasmid containing the green fluorescent protein (GFP) gene was also transfected into BHK-21 cells as a control of transfection efficiency. After 48 h, plasmid-transfected cells were fixed on microtiter plates by incubation for 30 min at 4°C with 90% (v/v) ethyl alcohol in distilled water. Plates were also washed with PBS (pH 7.4) with 0.1% Tween-20 between steps. Then hybridoma supernatants were incubated for 1 h at 37°C. After being washed three times, the plates were incubated for 1 h at 37°C with FITC-conjugated goat anti-mouse IgG antibody (Sigma, St. Louis, MO). Microtiter plates were examined with fluorescence microscope after being washed three times.
Western blot analysis
Briefly, virus-infected and non-infected BHK-21 cell sediments were subjected to electrophoresis on 10% SDS-PAGE after treating with dithiothreitol (DTT) at 100°C for 5 min. The protein was transferred onto a nitrocellulose membrane, and non-specific binding was blocked with 5% skim milk diluent overnight at 4°C. The membranes were incubated with the hybridoma supernatants as the primary antibody, and subsequently a peroxidase-conjugated goat anti-mouse IgG (H+L) antibody (ZSGB-BIO). The color was developed using 3,3'-diaminobenzidine tetrahydrochloride (DAB) and the reaction was stopped by distilled water, followed by drying the membrane.(12,13)
Indirect immunofluorescence assay
BTV1∼24, IBAV, CV, AKAV were added to BHK-21 monolayer cells in 96-well microtiter plates. After 48 h, cells were fixed on microtiter plates by incubation for 30 min at 4°C with 90% (v/v) ethyl alcohol in distilled water. Plates were also washed with PBS (pH 7.4) with 0.1% Tween-20 between steps. Then hybridoma supernatants were incubated for 1 h at 37°C. After washing three times, the plates were incubated for 1 h at 37°C with FITC-conjugated goat anti-mouse IgG antibody (Sigma). Microtiter plates were examined with fluorescence microscope after being washed three times.(14)
Results and Discussion
Viral titer of BTV16
TCID50 assay shows the viral titer in the virus infected cell supernatant is 107.5 TCID50/mL, according to the method of LaBarre and Lowy.(15)
Generation and characterization of two novel mouse anti-BTV16 MAbs
Using virus-infected cell supernatant as immunogen, we obtained two novel mouse anti-BTV16 MAbs named IVA4 and IB7. Subtype identification showed that both IVA4 and IB7 were IgG1 subclass with a κ light chain.
In order to confirm the specificity of the two Mabs, we conducted transfection of eukaryotic recombinant plasmids, immunofluorescence, and Western blot to test to which particular protein of BTV16 the MAbs bind. The transfection and immunofluorescence assay revealed that green fluorescence was only observed in the well in which pCI-neo-BT16-VP7 plasmid was transfected, but not in the other nine wells, within which the remaining nine gene segments of BTV16 were transfected (Fig. 1). Further, Western blot results showed that hybridoma supernatants of the two MAbs were only reactive with VP7 of BTV16, and not reactive with other proteins of BTV16 and BHK-21 cell proteins (Fig. 2). These two independent tests jointly confirmed that these two MAbs were mouse anti-BTV16 VP7 MAbs.

Transfected eukaryotic recombinant plasmids (pCI-neo) of BTV16 into BHK-21 and immunofluorescence. (
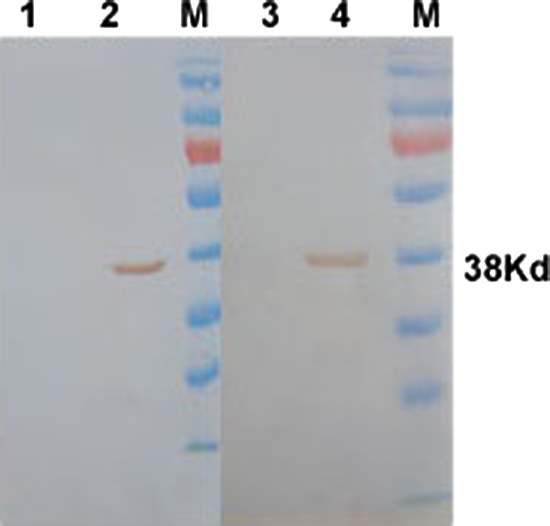
Western blot analysis of MAbs IB7 (lanes1,2) and IVA4 (lanes 3,4) against BTV16-VP7 (lanes 2,4) and BHK-21 cell lysate (lanes 1,3). M, PageRuler™ Prestained Protein Ladder (Fermentas, Burlington, Canada).
Response spectrums of MAbs
Indirect immunofluorescence results indicated that the two MAbs present different response spectrums with BTV 24 serotypes. IVA4 can react with BTV1∼24 serotypes, while IB7 reacts with BTV24 serotypes except for BTV 7, 15, 19 serotypes. And both MAbs have no cross-reaction with IBAV, CV, AKAV.
VP7 protein of BTV is the major group-specific protein, and is therefore an ideal immunogen for generating group-specific MAbs. Afshar and colleagues have reported group-specific MAbs against the core proteins of BTV, but whether they have cross-reaction with other viruses belonging to Reoviridae or transmitted by Culicoides species was not reported.(16) Zhou and Afshar reported MAbs to epizootic hemorrhagic disease virus of deer (EHDV) and BTV using recombinant VP7 of EHDV serotype 1 as immunogen, which had strong specificity to EHDV but poor group specificity to BTV.(17)
In this study, instead of using recombinant BTV-VP7 protein generated from a prokaryotic expression system, we used BTV as an immunogen to immunize BALB/c mice and screen MAbs. In this way, the MAbs obtained in this study would have a higher affinity with viral particle of BTV. The MAb IVA4 shows specific reaction with all 24 serotypes of BTV, while no cross-reaction was seen with IBAV, CV, which belong to the family Reoviridae, and AKAV, which is transmitted between hosts by bites of Culicoides species, possessing good group specificity. Therefore, this MAb may be a good candidate for the establishment of a specific diagnostic method for BTV such as immunofluorescence assay (IFA) and ELISA, after being labeled with fluorescein isothiocyanate or horseradish peroxidase.
This study was supported by Special Fund for Agro-scientific Research in the Public Interest (20120356); Fundamental Research Funds for Central Public Welfare Research Institutes (ZGKJ201105), Fundamental Research Funds for Central Public Welfare Research Institutes (0302012016).
Footnotes
Author Disclosure Statement
The authors have no financial interests to disclose.