
Abstract
Peripheral blood mononuclear cells in lepromatous leprosy (LL) patients produce low levels of interferon-γ (IFN-γ) and interleukin-12 (IL-12), and these cells exhibit partial or complete deficiency in the IL-12 receptor. The behavior of the IFN-γ receptor (IFN-γR) has not been described in cells from people with leprosy. We found higher levels of mRNA for IFN-γR1 and IFN-γR2 in adherent cells stimulated with IFN-γ and Mycobacterium leprae membrane proteins from LL patients compared with healthy subjects. Flow cytometry showed no significant difference in IFN-γR1 expression between LL patients and healthy subjects. Immunoblotting detected only the mature glycosylated form of the 61–67 kDa IFN-γR2 protein in healthy subjects. In contrast, cells from LL patients showed three different expression patterns: (1) the immature deglycosylated form of the 34.8 kDa IFN-γR2 protein, (2) the mature glycosylated 61–67 kDa form, and (3) both forms. Our data indicate the existence of abnormalities in the intracellular processing and protein expression of the IFN-γR in response to specific stimuli such as IFN-γ and M. leprae membrane proteins in adherent cells of LL patients.
Introduction
L
Leprosy induces a wide range of clinical, histological, and immunological manifestations, which are used to classify the disease into five types: tuberculoid leprosy (TT), borderline tuberculoid leprosy, borderline leprosy, borderline lepromatous leprosy, and lepromatous leprosy (LL). The cellular immune response predominates over the humoral immune response in TT patients and opposite in LL patients (Ridley and Jopling 1966;NG_BIB_000_036 Ivanyi 1986; Ochoa and others 1996).
An adequate immune response against intracellular parasites involves interferon-γ (IFN-γ), interleukin-12 (IL-12), and IL-18 and their receptors. Mycobacteria-infected macrophages and dendritic cells produce IL-12 and IL-18, which synergize to activate T and natural killer (NK) cells through their specific receptors, inducing the production of IFN-γ and promoting a Th1 response (Altare and others 1998b; Nakahira and others 2002). The IFN-γ receptor (IFN-γR) comprises two α-chains (IFN-γR1) and two β-chains (IFN-γR2). Binding of IFN-γ to the IFN-γR in macrophages or dendritic cells causes the transcription of numerous genes, including those coding for IL-12 and IL-18; binding to the IFN-γR also results in autocrine activation of T and NK cells (Nakahira and others 2002; Aguet and others 1988).
Patients with tuberculosis and other diseases caused by bacilli Calmette–Guerin (BCG) or atypical Mycobacteria, such as Mycobacterium fortuitum, Mycobacterium chelonae, Mycobacterium avium, have low levels of IFN-γ and IL-12, as well as down-regulated expression of receptors of these cytokines (Newport and others 1996; Altare and others 1998a; Jouanguy and others 2000).
Peripheral blood mononuclear cells (PBMC) from LL patients produce low levels of IFN-γ and IL-2 (Islas and others 1987; Fafutis and others 1999). This is a critical event for the production of IL-12, which remains low, and IL-12 concentration does not achieve a normal level, even after exogenous addition of IFN-γ (Libraty and others 1997).
IFN-γ production can be restored in PBMC from LL patients through the synergistic actions of exogenously added IL-12 and IL-18 (López and others 2008). However, LL patients show partial or complete deficiency in the IL-12 receptor (Kim and others 2001; Ohyama and others 2005).
The IFN-γR2 is expressed in many cell types, and is particularly important in monocytes–macrophages because it is needed to control intracellular infection by parasites, especially by Mycobacteria (Celada and others 1985; Altare and others 1998a).
mRNA and protein of IFN-γR1 and IFN-γR2 are down-regulated in macrophages from mice infected with M. avium (Hussain and others 1999). However, little is known about the expression of IFN-γR mRNA and proteins in leprosy. The aim of this study was to investigate the IFN-γR at both the protein and mRNA levels in adherent cells from LL patients.
Materials and Methods
Patients
Ten patients—two women and eight men (age, 23–63 years)—were recruited from the Dermatologic Institute, Guadalajara, México. All patients were classified as having LL according to the international criteria of Ridley and Jopling and MB according to World Health Organization (WHO) criteria (Ridley and Jopling 1966; Jacobson and Krahenbuhl 1999). All patients demonstrated a positive bacilloscopy (+++) and were treated with multidrug therapy recommended by the WHO (2 years clofazimine, rifampicin, and dapsone) (WHO 1982). Drug treatment was stopped ∼6 months before this study. At the time of cessation of the drug treatment, patients were considered cleared of leprosy infection. The control group included 10 healthy subjects (HS) matched for age and gender who were not related to the patients.
The study protocol was in accordance with the ethical guidelines of the Helsinki Declaration. Before blood donation, the subjects completed a medical history questionnaire and gave their written consent. The experimental protocol was approved by the Medical Research Ethics Committee at The University of Guadalajara.
Antigens and stimulus
Mycobacterium leprae membrane protein cocktail was kindly provided by Dr. John Spencer (Colorado State University, Ft. Collins, CO). Human recombinant IFN-γ (Roche, Mannheim, Germany) was used to stimulate adherent cell cultures.
Antibodies
Monoclonal antibodies (mAbs) were used for flow cytometry; these included phycoerythrin (PE)-conjugated mouse anti-human CD14 and fluorescein isothiocyanate (FITC)-conjugated CD119 (IFN-γ-R1), and their corresponding isotypes (R&D Systems Inc., Minneapolis, MN). Polyclonal rabbit antibody to the N-terminal group of the human IFN-γ-R2 (Santa Cruz Biotechnology, California, CA, USA), recognized fully glycosylated 61–67 kDa, and deglycosylated 34.8 kDa bands, was used to detect IFN-γ-R2 in Western blotting (Ishihara and others 1998; Banerjee and others 2005; Garcia-Tuñón and others 2007).
Cell preparation
Venous blood from patients and healthy subjects was collected, and PBMC were isolated by differential centrifugation over Ficoll–Hypaque (Sigma Chemical Co, St. Louis, MO)(Böyum 1988).
Immediately after isolation, PBMC (20 · 106) were cultured in Petri dishes (Nunc, Roskilde, Denmark) and incubated in RPMI-1640 for 2 h at 37°C in 5% CO2 and 95% air (Invitrogen: Life Technologies) supplemented with 10% heat-inactivated FBS, 1% antibiotic–antimycotic solution (Invitrogen: Life Technologies). Nonadherent cells were removed, and the adherent cells were removed from the bottom of the Petri dishes and resuspended in RPMI-1640 (Libraty and others 1997). Finally, 20 · 106 adherent cells were incubated for 24 h with 200 IU/mL of IFN-γ or 25 µg/mL of membrane proteins from M. leprae.
Quantitative real-time PCR of IFN-γR1 and IFN-γR2 mRNA
To quantify the mRNA of IFN-γR1 and IFN-γR2, total RNA was extracted with Trizol (Invitrogen: Life Technologies) (Chomcynski and Sacchi 1987) from 1 · 106 adherent cells after 20 h of culture with IFN-γ and M. leprae membrane proteins. Complementary DNA (cDNA) was obtained from 1 µg of total RNA using MML-V reverse transcriptase enzyme (Invitrogen: Life Technologies).
Primers and probes for real-time PCR for the target genes IFN-γR1 (NM_000416, GI 4557879) and IFN-γR2 (NM_005534, GI 5031782) were dyed with FAM (Applied Biosystems, Foster City, CA) and the housekeeping gene (18S RNA) with VIC (Applied Biosystems).
Each sample was analyzed in triplicate in a multiplex PCR. In brief, 2 µL of each cDNA sample was mixed with 1· of Universal Master Mix (Applied Biosystems), 1· of primer and probe of target gene, and 1· of housekeeping gene (Applied Biosystems) in a final volume of 50 µL. Amplification was carried out in an ABI Prism 7500 (Applied Biosystems) under the following conditions: 2 min at 50°C, 10 min at 95°C, and 40 cycles of 15 s at 95°C and 60 s at 60°C. To quantify the mRNA expression, the values are expressed as fold expression of the target gene relative to the expression of the reference control according to the formula 2−ΔΔCT arbitrary units (au).
Flow cytometry
After 24 h of culture, the adherent cells were harvested, mixed with AB-negative inactivated human serum to block the Fc receptors, and incubated at 4°C (Sellmayr and others 2001). Ten microliters of the mAbs, CD14-PE and CD119-FITC, or their respective isotype controls was added, and the cells were incubated for 30 min at room temperature in the dark. Finally, the adherent cells were fixed with 0.5% formaldehyde and were analyzed in a flow cytometer (EPICS XL-MCL, Beckman Coulter). The results are expressed as the index of the average fluorescence of the CD14+ CD119+ adherent cells.
Cell extracts
For protein analysis, 5 · 106 adherent cells and HeLa cells (used as a positive control for IFN-γR2 expression) were cultured in a polystyrene flask (Nunc) and stimulated with 200 IU/mL of IFN-γ for 24 h in the culture conditions described earlier. The cells were harvested and resuspended in cell extraction buffer comprising 10 mM Tris–HCl, 100 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1 mM NaF, 20 mM Na4P2O7, 2 mM Na3VO4, 1% Triton X-100, 10% glycerol, 0.1% SDS, 0.5% deoxycholate, 1 mM PMSF, and protease inhibitor cocktail (Sigma Chemical Co.) according to the manufacturer’s instructions. The cell suspension was centrifuged at 12,000g and the cell extract was collected and stored at −70°C until analyzed by electrophoresis.
Immunoblotting
The concentration of protein extracted from adherent cells and HeLa cells was quantified using the Bradford method (Bradford 1976). From each sample, 40 µg of protein was boiled in reducing sample buffer (10% SDS, Tris–HCl pH 8, 50% glycerol, 0.01 mM 2-β-mercaptoethanol, and 0.01% bromophenol blue) and resolved on an SDS–7.5% polyacrylamide gel. Proteins were transferred to a PVDF membrane (Bio-Rad Laboratories, Inc., Hercules, CA). The membrane was blocked with 5% non-fat-dried milk and incubated overnight at 4°C with the primary antibody to IFN-γR2 (1:200) in TBS buffer. A mouse/rabbit chemiluminescent Western blotting kit from Roche Diagnostics (Mannheim, Germany) was used to visualize the proteins.
Statistical analysis
The results are expressed as mean ± SD. Because the data were not distributed normally, they were analyzed by the nonparametric Mann–Whitney U-test using SPSS 10.0v software. A P value < 0.05 was considered significant.
Results
IFN-γR1 and IFN-γR2 mRNA expression
The IFN-γR1 mRNA level was higher in adherent cells from the LL patients than from the HS. The values in cells from LL patients were 6.8 ± 2.2 au when incubated in the presence of medium, 9.7 ± 4 au when incubated with IFN-γ, and 8 ± 2.8 au when incubated with M. leprae membrane proteins. The data obtained for HS were: 5 ± 2.3 au in cells incubated with medium, 6.1 ± 2.3 au in cells incubated with IFN-γ, and 3.5 ± 1 au in cells incubated with M. leprae membrane proteins.
Only the values for adherent cells stimulated with IFN-γ and M. leprae membrane proteins differed significantly between LL patients and HS (Fig. 1A, Table 1).
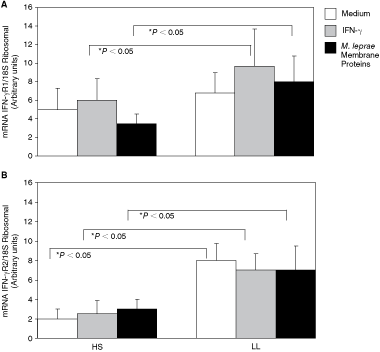
Expression of the mRNA IFN-γR1 and IFN-γR2 by real-time PCR. Quantitative analysis by real-time PCR and by retrotranscription of messengers from IFN-γR1 (
*P < 0.05 lepromatous leprosy patients versus healthy subjects.
The level of IFN-γR2 mRNA was also significantly higher in adherent cells from LL patients than from the controls. The values in cells from LL patients were 8 ± 1.7 au when incubated with medium, 7 ± 1.7 au when incubated with IFN-γ, and 7 ± 2.5 au when incubated with M. leprae membrane proteins. The corresponding values in HS were 2.0 ± 1 au (medium) 2.5 ± 1.4 au (IFN-γ), and 2.6 ± 1 au (M. leprae membrane proteins) (Fig. 1B, Table 1).
Fluorescence intensity of IFN-γR1 expression
The mean fluorescence intensity (MFI) for the expression of IFN-γR1 on the adherent cells membrane did not differ significantly between LL patients and HS. In cells from LL patients, the values were 17.6 ± 2.8 in cells incubated with medium, 16.5 ± 3 in cells incubated with IFN-γ, and 17.4 ± 2.6 in cells incubated with M. leprae membrane proteins. The corresponding values for the HS were 13.2 ± 2.2 (medium), 13.6 ± 2.2 (IFN-γ), and 13.6 ± 2.3 (M. leprae membrane proteins) (Figs. 2 and 3).
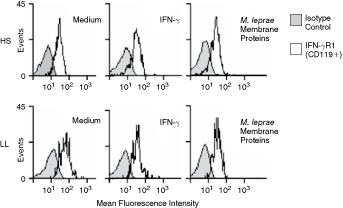
Expression of IFN-γR1 (CD119) on adherent cells from healthy subjects (HS) and lepromatous leprosy (LL) patients. Representative expression of IFN-γR1 on CD14+ adherent cells of HS (upper panels) and LL patients (lower panels) incubated with medium, IFN-γ, or Mycobacterium leprae membrane protein analyzed by flow cytometry. The shaded histogram indicates the isotype control, and the open histogram indicates IFN-γR1 in HS and LL patients.

Expression of IFN-γR1 (CD119) on CD14+ adherent cells from healthy subjects (HS) and lepromatous leprosy (LL) patients. The results are presented as the mean MFI ± SD of IFN-γR1 (FL1) on CD14+ adherent cells from HS and LL patients stimulated with IFN-γ or Mycobacterium leprae membrane protein.
IFN-γR2 protein expression
The IFN-γR2 protein expression was analyzed by Western blotting using antibodies that recognized the immature 34.8 kDa deglycosylated protein and mature 61–67 kDa glycosylated protein. We detected only the mature glycosylated form of the IFN-γR2 61–67 kDa protein in resting adherent cells from HS and HeLa cells (used as a positive control for expression of these proteins) and in cells stimulated with IFN-γ (Fig. 4A and 4B). In contrast, we found three different patterns of expression in adherent cells from LL patients: (a) the immature 34.8 kDa deglycosylated protein, (b) the mature 61–67 kDa glycosylated form, and (c) both the immature and mature forms (Fig. 4).

Expression of IFN-γR2 in adherent cells from lepromatous leprosy (LL) patients, healthy subjects (HS), and HeLa cells analyzed by Western blotting. The protein extracts from HeLa and adherent cells stimulated with IFN-γ were separated by SDS–7.5% polyacrylamide gel electrophoresis and then transferred to a PVDF membrane for immunoblotting. The detected bands corresponded to the molecular weights of 34.8 kDa and 61–67 kDa as shown by reaction with the rabbit polyclonal anti-human IFN-γR2 antibody. HeLa cells stimulated with IFN-γ (lane 1), mature glycosylated form of the IFN-γR2 61–67 kDa protein in adherent cells from HS (lanes 2, 3, 6, and 7), cells from LL patients showed three different expression patterns of IFN-γR2: (a) the immature deglycosylated form of the 34.8 kDa IFN-γR2 protein (lanes 12 and 13), (b) the mature glycosylated 61–67 kDa form (lanes 10 and 11), and (c) both forms (lanes 4, 5, 8, and 9).
Discussion
Leprosy is a chronic granulomatous infection caused by M. leprae. Based on the important role that IFN-γ and its receptor plays in mycobacterial diseases, the goal of this study was to analyze the expression of the IFN-γR and its protein and mRNA levels in adherent cells from LL patients.
IFN-γR1 is constitutively expressed and is thought not to be regulated by external stimuli (Farrar and Schreiber 1993). However, we found that CD14+ adherent cells from LL patients had higher levels of IFN-γR1 mRNA and protein in the resting state and after stimulation with IFN-γ or M. leprae membrane proteins compared with cells from healthy subjects. To our knowledge, no one has reported on the incremented levels of IFN-γR1 mRNA and protein in adherent cells from LL patients stimulated with IFN-γ.
The mycobacterial membrane fraction of M. leprae contains a number of immunologically important antigens that are recognized by cell-mediated and humoral responses in leprosy patients, including the 35 kDa major membrane protein I (MMP-I, ML0841) and bacterioferritin, the major membrane protein II (MMP-II, ML2038) (Brightbill and others 1999; Hertz and others 2001; Krutzik and others 2003). This fraction is particularly enriched for lipoproteins, a class of molecules that has been implicated in innate immunity against mycobacterial infection, particularly involving lipoproteins that stimulate the Toll-like receptor 2 (TLR-2) molecule on antigen-presenting cells. Dendritic cells (DC) from healthy individuals pulsed with MMP-II produced IL-12 p70 and up-regulated the surface expression of MHC class I, class II, CD86, and CD83, all of which are molecules that are important in the acquired immune response to infection (Brightbill and others 1999; Hertz and others 2001; Krutzik and others 2003). In addition, TLR-2 signaling was involved in DC maturation induced by MMP-II (Maeda and others 2005). Thus, the membrane fraction of M. leprae appears to have the ability to activate antigen-presenting cells, such as DCs, and to be involved in stimulating innate immunity, both of which would be important against mycobacterial infections such as tuberculosis and leprosy.
The overexpression of mRNA IFN-γR1 in adherent cells from LL patients observed in our study probably reflects an effect on the signaling mechanism, as mentioned earlier.
In another hand, the immature form of IFN-γR2 (34.8 kDa) has been detected in HeLa cells and human monocytes (Hemmi and others 1994; Soh and others 1994; Sakatsume and others 1995). This form goes through post-translational modification by the addition of carbohydrates, and its mature form in monocytes and HeLa cells weighs between 61 and 67 kDa (Bach and others 1996).
The expression of IFN-γR2 mRNA and protein is regulated by several stimuli including IFN-γ by itself in T cells from peripheral blood (Bach and others 1995; Sakatsume and Finbloom 1996).
We found significantly elevated IFN-γR2 mRNA levels in resting adherent cells from LL patients and in cells stimulated with IFN-γ or M. leprae membrane proteins compared with cells from healthy subjects. This suggests that, in patients with LL, the expression of IFN-γR2 mRNA is up-regulated when cells are exposed to M. leprae membrane proteins or IFN-γ.
We detected the mature glycosylated form of the IFN-γR2 protein (61–67 kDa) in HeLa cells (positive control in Western blotting) and in adherent cells from HS. In contrast, adherent cells from LL patients exhibited three different expression patterns: (a) the immature deglycosylated 34.8 kDa form of the IFN-γR2 protein, (b) the mature 61–67 kDa glycosylated form, and (c) both forms. We note that these forms were detected both in cells with and without IFN-γ stimulation.
A recent report showed that missense and other in-frame mutations can cause deleterious effects at the cellular level through different mechanisms, as when new carbohydrates are added to proteins. One patient with Mendelian susceptibility to mycobacterial disease showed a complete deficiency of the IFN-γR2. This deficiency may reflect an in-frame IFN-γR2 mutation (382–387 dup), which may be associated with the loss of gene expression, loss of surface expression, or non-functional receptor (Vogt and others 2007). Several mutants of the IFN-γR2 with the 382–387 dup mutation result in proteins of low molecular weight that fail to mature in a normal way.
Proteins of high molecular weight may also be produced as a result of excessive N-glycosylation (Vogt and others 2007; Vogt and others 2008).
We found both the mature form (61–67 kDa) and the immature form (34.8 kDa) of IFN-γR2 in adherent cells from LL patients. However, we did not sequence the gene that codes for IFN-γR2 protein in LL patients. A detailed analysis of the gene sequence for the IFN-γR2 protein will identify whether mutations exist that result in abnormalities of the glycosylation sites.
Getting all the most significant immunologic abnormalities of LL patients cells reported in the literature for trying to understand and explain the anergy presented by the patients in this pole of the disease, it has been published: (a) INF-γ low level in PBMC (Fafutis and Cols 1999) that recover under synergic action of exogenous addition of IL-12+IL-18 (López and others 2008), independently that the T cells of LL patients show total or partial deficiency of IL-12 receptor (Kim and others 2001). (b) Low levels of IL-12 are expressed on LL patients, which are not able to recover still after a INF-γ exogenous addition (Libraty and others 1997) provably due to a failure on the INF-γR expression in monocytic–macrophagic series.aq4
In the present study, we found abnormalities in the INF-γR expression to transcriptional or translational level, such as IFN-γR2 glycosylation in adherent cells of LL patients. Bioassays are required to understand the functionality of IFN-R2 and to determine whether the abundance of mRNA for this protein associated with LL results in increased protein expression on changes in receptor function.
Leprosy induces a wide range of clinical, histological, and immunological manifestations. It is mandatory to make studies that include the mRNA and proteins expression involved with the immune response against M. leprae through the spectrum of the disease, in order to have knowledge about the relationship between the immunologic abnormalities and the clinical manifestations in leprosy disease.
Footnotes
Acknowledgments
This work was supported by CONACyT Grant 31121. We want to thank Dr. John Spencer and Dr. Patrick Brennan (Colorado State University) for their kindly providing the M. leprae proteins, because without all this there was no way to realize this work.
Author Disclosure Statement
No competing financial interest exists.