
Abstract
Cryptococcal meningitis is often associated with elevated IL-10 levels, which suggest a dysregulation in the antifungal immune response. β-Arrestin 2 plays a major role in desensitization of G-protein-coupled receptors involved in the immune responses, provides a scaffolding platform for modification of many signal transduction proteins, and binds Src and MAP kinases family members. This study compared the levels of β-arrestin 2 mRNA and protein in peripheral blood mononuclear cells (PBMC) of patients with cryptococcal meningitis detected. The interferon-γ (IFN-γ) serum concentration was determined with enzyme-linked immunosorbent assay (ELISA) to reveal its relationship with β-arrestin 2. The effect of modulation of β-arrestin 2 on cytotoxic activity against Cryptococcus was explored via transfection and interference of β-arrestin 2. PBMCs of patients with cryptococcal meningitis exhibited significantly elevated levels of β-arrestin 2 and a positive correlation between β-arrestin 2 and IL-10 levels existed in patients’ serum, but a negative correlation was found between β-arrestin 2 and IFN-γ expression. In conclusion, elevated expression of β-arrestin 2 in PBMCs of patients with cryptococcal meningitis correlated with a reduced cytotoxic activity against Cryptococcus. This study suggests that reduced β-arrestin 2 mRNA levels or inhibition of β-arrestin 2 activity may augment INF-γ production, and ultimately, the anti-Cryptococcus immune response of infected patients.
Introduction
T
The immune response to cryptococcal infection entails activation of the innate immune system including the potent antigen-presenting cells, mature dendritic cells (mDCs), NK cells, neutrophils, the secretion of cytokines, which aid intercellular communication, and the activation of B cells, antibody production, and T cells (Schop 2007). T-cell–mediated immune responses play a key role in the host’s response and outcome to cryptococcal infection (Singh and others 2006; Vecchiarelli 2007). Helper T cells (TH) are subdivided into 2 classes, called TH1 and TH2 (Scheinecker and others 2008). The cytokines, IL-2, IFN-γ, and TNF-α are mainly produced by TH1 cells and their predominance correlates with resistance to long-term cryptococcal infection (Netea and others 2004; Siddiqui and others 2005; Biondo and others 2008). In contrast, IL-4, IL-6, and IL-10 are mainly produced by TH2 cells and high levels are associated with reduced efficiency of the anti-cryptococcal response (Singh and others 2006), and without further intervention, a poor clinical response. The secreted cytokines help orchestrate the innate and adaptive immune responses by attaching to their respective receptors, and triggering a signaling cascade that culminates in the activation and suppression of the immune function necessary to eliminate cryptococcal infection.
Cellular signal transduction is a major focus in international life science research in the recent decades. G-protein-coupled receptor (GPCR), an important hinge to transfer cellular signal, is one of the most critical drug targets (Siehler 2008). Activation of the GPCR leads to post-transcriptional modifications of various signaling proteins, such as CREB, AP-1, and NF-κB, and modulation of gene expression. The multifunctional molecule, β-arrestin 2 plays a pivotal role in desensitization and endocytosis of different types of GPCR, and binds various signaling molecules such as Src-family kinases, certain components of mitogen-activated protein kinase (MAPK) cascade, and E3 ligase Mdm2 (Lynch and others 2007; Ma and Pei 2007; Barki-Harrington and Rockman 2008). Interactions with β-arrestins usually modulate phosphorylation, ubiquitination, and/or subcellular distribution of their binding partners. β-Arrestin 2 was shown to block TNF-induced phosphorylation and degradation of IκB, interact with TRAF6, and negatively regulate Toll-like interleukin-1 receptor (TLR-IL-1R) signaling pathways (Wang and others 2006; Kawamata and others 2007). Recent data indicate that β-arrestin 2 plays a pivotal role in modulating NK activity and β-arrestin 2-deficient mice exhibited a greater efficacy against cytomegalovirus infection (Yu and others 2008). Paeoniflorin reduced inflammation in an adjuvant arthritis murine model as well as expression of β-arrestin 2 in mesenteric lymph nodes and may have induced TH1 tolerance (Wu and others 2007). However, the role of β-arrestin 2 in the immunoregulation of the pathogenesis of cryptococcal meningitis is unknown.
This study determined the expression level of β-arrestin 2 mRNA and protein in peripheral blood mononuclear cells (PBMCs) of patients with cryptococcal meningitis and further explored regulatory mechanisms of β-arrestin 2 in terms of cytotoxicity of PBMCs specific to Cryptococcus.
Materials and Methods
Patients
This study received informed consent from all patients and was approved by the Ethics Committee of Shanghai Changzheng Hospital. Thirty patients (18 males and 12 females) with cryptococcal meningitis were included in this study, mean age was 37 years old (range from 23 to 67 years). Cerebrospinal fluid (CSF) was obtained from patients by lumbar puncture. CSF was serially diluted and spotted onto Sabouraud dextrose agar plate. C. neoformans was identified from CSF and/or CSF culture. There were 30 controls (15 males and 15 females), mean age 38 years old (range from 21 to 52 years). All samples were detected as HIV-negative.
Antibodies, reagents, and plasmids
RNA was extracted with RNeasy Mini Kits (Qiagen, Croningen, The Netherlands). Levels of IL-4, IL-10, TNF-α, and IFN-γ were determined with enzyme-linked immunosorbent assay (ELISA) Kits (BioSource International Inc., Camarillo, CA). All chemicals, unless otherwise specified, were purchased from Sigma-Aldrich (St. Louis, MO). Plasmid cDNA encoding βgal, βarr2, pBS/U6/βarr2 siRNA, and pBS/U6/nonspecific siRNA plasmids were prepared as previously described (Wang and others 2006; Kawamata and others 2007; Lynch and others 2007; Barki-Harrington and Rockman 2008).
Purification of PBMCs and extraction of total RNA
Human whole blood (3 mL) was collected in sodium citrate anticoagulation tubes and diluted with equal volume of phosphate-buffered saline. The mixture was slowly transferred onto Lymphocytes Separation Medium Lymphoprep™ (AXIS-SHIELD, Norway) and centrifuged at 2,000 rpm for 20 min and the layer of mononuclear cells was collected. Total RNAs of PBMCs was extracted using Trizol reagent according to the manufacturer’s instructions (Invitrogen, Carlsbad, CA). After chloroform was added, mixed vigorously, and centrifuged at 10,000g for 20 min at 4°C, total RNA was precipitated from the aqueous phase with an equal volume of isopropanol and pelleted by centrifugation at 13,000g for 10 min at room temperature. Pellets containing RNA were washed once with 75% ethanol and resuspended in diethylpyrocarbonate (DEPC) treated water. The concentrations and purities of the RNA preparations were determined by absorbance at 260 and 280 nm. Total RNA were stored at −70°C for subsequent reverse transcription PCR.
Reverse transcription
Reverse transcription of 2 µg total RNA as template (2 µg) was performed in reaction mixtures containing l× Reverse Transcription Buffer, 0.5 mmol/L dNTPs, l µmol/L Oligo-dT primer, 10 U RNase inhibitor, and 4 U Superscript™ α reverse transcriptase (TaqMan Reverse Transcription Kits (Applied Biosystems, Foster City, CA). The reaction mixtures were incubated at 25°C for 10 min, at 42°C for 50 min, and then at 85°C for 5 min. The reaction mixtures were chilled on ice for 5 min and incubated with 1µL E. coli RNase at 37°C for 20 min to remove possible remaining RNA according to the manufacturer’s instructions. Produced cDNA were stored at −20°C for subsequent quantitative real-time PCR.
Primers and probes for quantitative real-time PCR
Primers and probes were designed using TaqMan-MGB technology and Primer Express software according to human β-arrestin 2 and GAPDH mRNA sequence available from GenBank. 5′-Terminal of probes was labeled with Fluorescent light-emitting group FAM (6-carboxy-fluorescein). Non-Fluorescent Quencher MGB (Minor Groove Binder) was used to label the 3′-terminal of probes instead of general Fluorescent Quencher TAMRA (6-carboxy-tetramethylrhodamine), to eliminate background fluorescence and enhance the resolution and specificity of signal. All primers and probes used were synthesized by Applied Biosystems. β-Arrestin 2: forward primer: 5′-CCCTGG ATGGGAAACTCAAG-3′, reverse primer: 5′-AGGATT CCCAGCACCTCCTT-3′; probe: FAM-CAGCACCATCGTG AAG-MGB. GAPDH: forward primer: 5′-CCATCAATGACC CCTTCATTG-3′, reverse primer: 5′-CATGGGTGGAA TCATATTGGAAC-3′, probe: FAM-CCTCAACTACATGGT TTAC-MGB.
Quantitative real-time PCR
Real-time PCR was carried out in a 20-µL reaction mixture containing 50 ng template cDNA, 10 µL PCR Master mix, 1 µL 20× mix dNTP, 8 pmol primers, 4 pmol probes, and RNAase-free water. The PCR began with a hot start at 50°C for 2 min and 95°C for 10 min, and proceeded with 40 cycles of denaturation at 95°C for 15 s, annealing and extension at 60°C for 1 min with a final extension at 60°C for 10 min. Relative level of β-arrestin 2 mRNA in patients and control groups was obtained by comparison of cycle threshold (Ct) difference between β-arrestin 2 and control gene GAPDH. Relative quantitation of interested gene was calculated according to the following formula. Relative quantitation of interested gene = 2−(ΔCt) and ΔΔCt = (ΔCt, Q) − (ΔCt, C), where (ΔCt, Q) and (ΔCt, C) are Ct differences between interested gene and control gene both in patients and normal groups, respectively.
Determination of IL-4, IL-10, TNF-α, IFN-γ, immunoglobulin isotypes, and complement components in serum by ELISA and array protein assay
Serum was obtained from peripheral blood and stored at −80°C. Different cytokines were detected blindly using ELISA Kits (BioSource) and confirmed by blocking at 4°C overnight with corresponding monoclonal antibodies. Calibration curve was drawn from titrated human recombinant 4-1BBL-Fc and IL-18 results. After blocking with BSA for 2 h, samples were diluted to determine concentration, placed in wells, and incubated at 37°C for 2 h. After washing 3 times, HRP-streptavidin was added, and plates were incubated at 37°C for 1 h. After washing, TMB peroxidase was added and cytokine concentrations (× dilution factors) were detected at 450 nm wavelength. Levels of IgG, IgA, IgM, C3, and C4 in PBMCs from patients and normal people were determined by DADE BEHRING BN2 array protein system.
Western blot
Compared with control gene β-actin, level of β-arrestin 2 protein in PBMCs of patients with cryptococcal meningitis was detected by antibodies against β-arrestin 2 (Santa Cruz Biotechnology, Santa Cruz, CA) and Western blotting using enhanced chemiluminescence (ECL) as specified by the manufacturer (GE Healthcare).
Preparation of inactivated Cryptococcus
C. neoformans strain BLS27 (Chinese Medicine Preservation and Administration of Fungus Center) was stored in Sabouraud’s medium at −70°C. A single colony was picked and inoculated into YEPD medium and incubated at 37°C for 2 days with moderate shaking. Harvested cells were washed 3 times and diluted to a final concentration at 1 × 106 cells/mL with sterile 0.9% NaCl. Yeast cells were counted in a hemocytometer and autoclaved at 121 kPa for 10 min and then stored at −20°C in aliquots.
Preparation of cryptococcal-specific PBMC and transfection
To generate cryptococcal-specific PBMC, PBMC were repeatedly stimulated with autologous mature dendritic cells (mDCs) pulsed with inactivated Cryptococcus strain B3501 (1 × 106 cells/mL). In brief, mDCs were harvested, washed with serum-free medium, and pulsed overnight with 100 µg/mL of inactivated Cryptococcus. The cryptococcal-stimulated mDCs were harvested, washed, irradiated at 10 Gy, resuspended in RPMI 1640 medium supplemented with 10% FCS, and were used to prime autologous PBMC at a 1:20 stimulator-to-responder cell ratio. Cultures were restimulated every 7 days with irradiated cryptococcal-pulsed mDCs, and a total of 3 stimulations were done. IL-2 (100 U/mL) was added to the cultures 2 days after the second stimulation and the cells were fed twice a week with fresh medium containing 10% FCS and IL-2 (100 U/mL). After 1 week since the third stimulation, cryptococcal-specific PBMC were processed for transfection with plasmids of βarr2 silence or overexpression by Human PBMC Nucleofecor® Kits (Amaxa Inc., Gaithersburg, MD) following manufacturer’s protocol, for function and expression assays. For all transfection experiments, controls included CMV-β-gal (for expression), or nonspecific siRNA (NS siRNA) for siRNA. Silence or overexpression of β-arrestin 2 in the transfected PBMC was confirmed by Western blotting. The treated cells were continued to cytotoxicity experiments.
Cytotoxicity experiments to Cryptococcus on PBMCs
The 5 × 103 CFUs Cryptococcus and 10 × 103 PBMCs were co-incubated in 150 µL medium containing 10% mixed human serum at 37°C. Twenty-four hours later, cells were lysed with 0.1% Triton X-100 and serial dilutions were plated on sabouraud dextrose agar at 24 and 48 h post-culture. (Pretest results suggested that viability of fungus was not affected by Triton X-100.) The number of viable C. neoformans was determined by colony counts. The control groups contained Cryptococcus, medium and human mixed serum, and PBMC. Proportion of Cryptococcus was calculated by the following formula: (Experimental CFU/Inoculated CFU-1) × 100. The growth ratio of C. neoformans is defined as “0” if the start and the final incubated CFU were the same, double amount of Cryptococcus as 100 and invalid Cryptococcus as <0.
Statistical analysis
Continuous data were expressed as mean ± standard deviation (SD) or median (interquartile range) and tested with Student’s t-test or Mann–Whitney test if variables were with large standard deviation. Categorical data were expressed as numbers (%) and tested with chi-square test. The correlation was tested with Spearman’s correlation coefficients because variables had high standard deviation and were presented with scatter plot. The repeated measures data were evaluated by linear mixed model. The analyses were performed using the SPSS 15.0 software package (SPSS Institute Inc., Chicago, IL). All statistical assessments were 2-sided using a significance level of 0.05.
Results
Overexpression of β-arrestin 2 in PBMCs of patients with cryptococcal meningitis
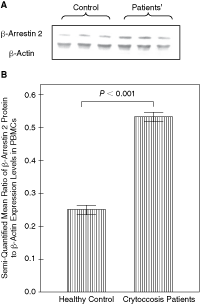
Expression of β-arrestin 2 in PBMCs of normal people and patients with cryptococcal meningitis was detected by Western blotting with ECL compared with control gene β-actin as shown in Figure 1. The relative production of β-arrestin 2 was significantly greater in patients with cryptococcal meningitis (0.5326) than normal controls (0.2487) (P < 0.01).
(
Table 1 presented demographic characteristics and laboratory data of the 30 case group (n = 30), control group (n = 30), and the 60 total subjects, including 33 (55.0%) males and 27 (45.0%) females. The PBMCs level had a median (interquartile range) of 0.39 (0.25–0.53). The β-arrestin 2 mRNA level exhibited a median (interquartile range) of 1.98 (1.00–3.26). While the IFN-γ had a median level (interquartile range) of 34.96 pg/mL (5.14–69.01 pg/mL), the mean IL-4 level in total subjects was 2.37 ± 0.21 (pg/mL). As shown in Table 1, the PBMCs level and β-arrestin 2 mRNA level were significantly higher in case group than control group, but the IFN-γ level was lower in case group than control group. The IL-4 level was also higher in case group than control group (2.43 ± 0.17 pg/mL vs. 2.31 ± 0.23 pg/mL; P = 0.026).
D
Continuous data were expressed as mean ± SD or median (interquartile range). Categorical data were expressed as frequencies and percentage and tested with chi square.
aStudent’s t-test.
bChi-square test.
cWilcoxon Rank Sums test.
*Statistical significance, P < 0.05.
Abbreviations: IFN, interferon; IL, interleukin; PBMC, peripheral blood mononuclear cell; TNF, tumor necrosis factor.
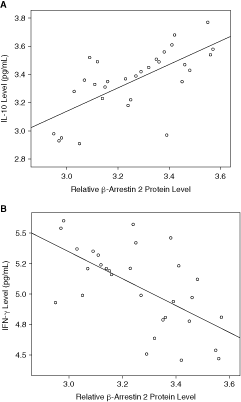
Figure 2A presented the scatter plot that showed the association between the relative protein expression levels of β-arrestin 2 and IL-10 level (pg/mL) in the case group. The correlation was a significant, medium positive correlation tested by Spearman’s correlation coefficients (n = 30; Spearman’s δ = 0.681; P < 0.001). Figure 2B presented the scatter plot that showed the association between the protein relative expression levels of β-arrestin 2 and IFN-γ level (pg/mL) in the case group. There was a significant, medium negative correlation tested by Spearman’s correlation coefficients (n = 30; Spearman’s δ = −0.597; P = 0.001).
(
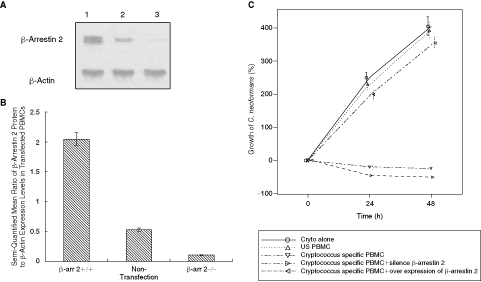
In order to further explore the correlation between β-arrestin 2 levels and reduced IFN-γ production, a transfection of β-arrestin 2 gene was performed on cryptococcal-specific PBMC by electroporation. In brief, cryptococcal-specific PBMC were prepared as described in methods. The extent of β-arrestin 2 overexpression was confirmed by Western blot 48 h after transfection (Fig. 3A). β-Arrestin 2-transfected PBMC exhibited an overexpression of β-arrestin 2 (mean ratio of β-arrestin 2:β-actin: 2.04) compared to non-transfected control PBMCs (0.53; P < 0.01) (Fig. 3B). The βarr2-transfected PBMC only modestly suppressed the growth of Cryptococcus and thus, the βarr2-transfected PBMC exhibited significantly reduced anti-cryptococcal cytotoxic activity (Fig. 3C; P < 0.01).
(
Conversely, transfection of siRNA β-arrestin oligonucleotides into anti-cryptococcal-specific PBMCs reduced the expression of β-arrestin (0.1248) in comparison to control siRNA-transfected PBMCs (0.5012; Fig. 3A). After transfection with β-arrestin 2 siRNA or control oligonucleotides, PBMC cells were investigated for anti-cryptococcal activity. Comparisons between each cryptococcal-specific PBMC and growth of C. neoformans (%) were evaluated by a linear mixed model considering each group repeated measures per 24 h. Silencing of β-arrestin 2 expression led to a nonsignificant increase in anti-cryptococcal activity as shown in Figure 3C.
Discussion
Currently, the incidence of cryptococcal meningitis is increasing in China. Curative treatment remains a great challenge for cryptococcal meningitis due to its currently long therapeutic course, concomitant side effects, and the high variability of Cryptococcus (Guerrero and Fries 2008). Therefore, our current studies focused on elucidation of targets that would improve the efficacy of antifungal therapy and enhance protective immune response against Cryptococcus sp.
The arrestin proteins family includes the ubiquitous β-arrestin 1 and 2, and 2 classes of arrestins expressed exclusively in retina (rod and cone arrestins) of mammals (Ma and Pei 2007). β-arrestins 1 and 2 play key roles in desensitization and endocytosis of GPCRs as well as bridging with the intracellular signal transduction molecules (Ma and Pei 2007). A previous study demonstrated that the binding of β-arrestin 2 to IκB suppressed its phosphorylation, reduced its degradation by ubiquitin. Consequently, NFκB–IκB complexes could not be dissociated, NFκB could not be activated, and TNF-α signal transduction were regulated (Kawamata and others 2007). Endotoxin-induced proinflammatory cytokines were up-regulated in β-arrestin 2-deficient mice and these mice were more susceptible to endotoxic shock (Wang and others 2006). Thus, β-arrestins may thus serve as potential therapeutic targets to control inflammatory diseases.
In this study, overexpression of β-arrestin 2 in PBMCs of patients with cryptococcal meningitis possibly contributed to a decline of cytotoxicity against Cryptococcus and increased disease severity, consistent with the decline of β-arrestin 2 and improved efficacy against cytomegalovirus (Yu and others 2008). Results from in vitro experiments suggested that cytotoxicity of cryptococcal-specific PBMC decreased with overexpression of β-arrestin 2. These results suggested that boosting a TH1 immune response may lead to an efficient treatment that eliminates cryptococcal meningitis, as observed (Siddiqui and others 2005). There were no significant changes in cytotoxicity of PBMC specific to Cryptococcus after gene silencing of β-arrestin 2. This result suggested that signaling pathways besides β-arrestin 2 played major roles in the cytotoxicity responses.
Pietrella et al. demonstrated that the mannoprotein from C. neoformans stimulated early IL-12 secretion, which induced IFN-γ secretion by NK cells and led to an effective TH1 response to Cryptococcus (2004). However, a reduction or delay in IL-12 secretion modified the TH1 anti-cryptococcal-specific immune response, weakened resistance against pulmonary cryptococcosis, and disseminated cryptococcosis possibly ascribed to reduction of IFN-γ (Pietrella and others 2004). Additionally, elimination of Cryptococcus was weakened by reduced MHC-α production on APCs and decrease of inflammatory cytokines IL-1β and TNF-α from monocytes. In this study, patients with cryptococcal meningitis had significantly reduced IFN-γ expression in serum in comparison to a control group, similar to renal transplant patients with cryptococcal meningitis (Singh and others 2006). Furthermore, they overexpressed IL-10 in serum compared with the control group, analogous to renal transplant patients with cryptococcal meningitis (Singh and others 2006). Production of β-arrestin 2 in PBMCs correlated positively with level of IL-10, but negatively correlated with level of IFN-γ in serum. This result suggested that overexpression of β-arrestin 2 possibly led to immune dysregulation and a skewing of the ratio from TH1 to TH2, which has correlated with a lower immune response against fungus and outbreaks of Cryptococcus sp. (Singh and others 2006). The effect of β-arrestin 2 modulation on the TH2-associated IL-13 expression is also of interest. Therefore, further studies on the initial immune response to Cryptococcus, its maintenance, and immunoregulatory mechanisms such as β-arrestin 2 are warranted.
Footnotes
Acknowledgments
This research was supported by grants from the Ministry of Science and Technology (#2006AA02Z496), the National Science Foundation of China (#30671840, #30772017, #30770997), and the Shanghai Municipal Commission for Science and Technology (08QH14001, 07XD14013, 09JC1405400).
Author Disclosure Statement
No competing financial interests exist.