
Abstract
The granulocyte–macrophage colony-stimulating factor (GM-CSF) is a cytokine with many important applications and, due to its immunostimulatory properties, could also be used as a vaccine adjuvant. A simple strategy to produce recombinant mouse GM-CSF (mGM-CSF) in transgenic Nicotiana tabacum plants was used in this study. The mGM-CSF cDNA followed by the sequence encoding endoplasmic reticulum retention signal (KDEL) was cloned into the ImpactVector under the control of the strong promoter from the gene encoding a small subunit of Rubisco. In transgenic plants the accumulation level of recombinant mGM-CSF varied in the individual transformants from 8 to 19 µg/g of fresh leaf tissue, which makes up to 0.22% of total soluble protein. In most analyzed plants, the apparent molecular weight of the recombinant protein was larger than predicted due to its N-glycosylation, presumably in 2 sites. The recombinant plant-produced murine GM-CSF retained its biological activity as confirmed in vitro in proliferation assay using a mouse cell line, which is growth-dependent on GM-CSF.
Introduction
G
Genetically transformed plants and plant cells became an attractive source of recombinant proteins such as mammalian antibodies, blood substitutes, cytokines, or vaccines (Twyman and others 2003; Maliga and Graham 2004; Schillberg and others 2005; Saint-Jore-Dupas and others 2007; Streatfield 2007). Several research groups reported production of hGM-CSF not only in transformed plant cells suspension cultures of tobacco and rice (Matsumoto and others 1995; James and others 2000; Hong and others 2002; Lee and others 2002; Shin and others 2003; Kim and others 2008a, 2008b, 2008c), but also in transgenic plants such as tobacco (Sardana and others 2002), sugarcane (Wang and others 2005), and rice (Sardana and others 2007; Ning and others 2008).
The hGM-CSF is not fully active in mice cells and the murine GM-CSF is not fully active in human cells (COPE Cytokines & Cells Online Pathfinder Encyclopaedia, [http://www.copewithcytokines.de]). Although Ning and others (2008) tested the biological activity of recombinant plant-produced hGM-CSF in mice, the usefulness and reliability of the cross-species tests might be in the case of this protein controversial since the mouse and human GM-CSF share about 54% of identity and are species-specific. We are not aware of any reports on production of murine GM-CSF in plants. The main goal of this work was efficient production of biologically active mouse GM-CSF (mGM-CSF), which could be potentially applied in variety of preclinical tests conducted in murine models. The binary plasmid based on ImpactVector and containing the plant expression cassette with mGM-CSF cDNA controlled by the promoter and the transcriptional terminator of rbcS1 gene from Chrysanthemum morifolium was used for the transformation of Nicotiana tabacum. The low level of alkaloids in the used tobacco cultivar, LA Burley 21, will allow to use such plant material in oral or nasal administrations without any purification steps to test the immunostimulatory properties of the recombinant plant-produced mGM-CSF.
Materials and Methods
Construction of plasmids for plant transformation
The mGM-CSF cDNA was amplified from a recombinant plasmid kindly provided by Dr. Falko Steinbach, Veterinary Laboratories Agency, UK using PCR technique with 2 specific primers: NcoGMCSF (5′-TCCCATGGCACCCACCCGCTC-3′) and SacGMCSF (5′-GGAGCTCTTAAAGCTCATCTTTTTGGAC-3′). The samples were subjected to the following program: 4 min at 94°C; 30 cycles (30 s at 92°C, 30 s at 60°C, 30 s at 72°C); 3 min at 74°C. The generated 398-bp PCR product contained mGM-CSF cDNA without sequence encoding its own N-terminal signal sequence. The amplified DNA fragment was identical in the region covering the coding part of mature mGM-CSF to the sequences available in databases (eg, GenBank Acc. No. BC116880). Following NcoI and SacI digestion, the PCR product was cloned into the ImpactVector 1.3 (Plant Research International, Wageningen; [http://www.pri.wur.nl/UK/products/ImpactVector/]). The recombinant proteins contained N-terminal signal peptide from Anemone equistatin and C-terminal KDEL sequence responsible for its retention in endoplasmic reticulum (ER), and the transgene was controlled by the promoter and transcriptional terminator of rbcS1 gene from C. morifolium. Subsequently, the AscI–PacI excised expression cassette was re-cloned into the binary vector, pBINPLUS (Plant Research International, Wageningen). The resulting recombinant plasmid, pBIN/mGMCSF/KDEL (Fig. 1A), was used in a plant transformation procedure.
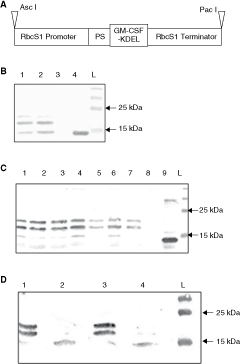
Production of recombinant mouse granulocyte–macrophage colony-stimulating factor (mGM-CSF) in plants. (
Transient plant transformation
Fifty milliliters of LB medium containing kanamycin (50 µg/mL), 10 mM 2-[N-morpholino] ethanesulfonic acid (MES) pH 6.3 and 20 µM acetosyringen was inoculated (1:10) with overnight culture of Agrobacterium tumefaciens strain LBA 4404 and grown overnight at 28°C. Then, bacterial pellet collected after centrifugation at 3,000g, 5 min was resuspended in the buffer containing 10 mM MES pH 6.3 and 10 mM MgCl2 to a final volume giving the optical density OD600 = 0.5. The bacterial suspension was incubated at room temperature for 3 h. Next, using 1 mL syringe without a needle the bacterial suspension was injected into the underside of leaves of the Nicotiana benthamiana plants. Three days later, the infiltrated leaves were collected and subjected to Western blot analysis to confirm protein production.
Plant transformation and plant growth conditions
The recombinant plasmid pBIN/mGMCSF/KDEL was introduced into electrocompetent A. tumefaciens strain LBA4404 by electroporation and subsequently used for transformation of the seedlings of a low alkaloid line of tobacco, LA Burley 21 (Legg and others 1970). The transformation procedure was performed according to the described procedure (Rossi and others 1993). Plants were grown on MS medium (Murashige and Skoog 1962) supplemented with MS vitamins and kanamycin (150 µg/mL). Plants cultivated in vitro were kept in a sterile containers in a culture room in the following conditions: temperature 24°C, day/night cycles—16 h/8 h. Transformants selected for seed production were transferred to soil, self-pollinated, and maintained in the greenhouse.
Genomic PCR and Northern blot analysis
Genomic DNA for PCR analysis was prepared by grinding a small leaf piece (a disc of 0.5 cm in diameter) from one plant with 0.4 mL of extraction buffer (200 mM Tris–HCl, pH 7.5, 250 mM NaCl, 25 mM EDTA, 0.5% SDS) and incubation at RT for 1 h. The extract was clarified by 5-min centrifugation (12,000g), then, DNA was precipitated with equal volume of isopropanol. Pellet was dissolved in 0.1 mL of TE buffer (pH 8.0) and 1 µL was used in PCR analysis.
RNA was isolated using RNasy Plant Mini Kit (Qiagen, Hilden, Germany). RNA samples, 20 µg/well, were separated on a 1.2% agarose with 2% formaldehyde according to the standard procedure (Sambrook and others 1989). Northern blots were hybridized with DIG-labeled RNA probe complementary to mGM-CSF transcript using DIG Northern Starter Kit (Roche, Mannheim, Germany). The probe was generated according to manufacturer’s protocol (Roche) in an in vitro transcription reaction containing 1 µg of linearized recombinant plasmid carrying full-length mGM-CSF cDNA. Recombinant plasmid used as DNA template for in vitro transcription was linearized at restriction enzyme site located downstream of the mGM-CSF cDNA. After the restriction digestion, the DNA was purified by phenol/chloroform extraction.
Protein extraction
Leaves frozen in liquid nitrogen were ground in the extraction buffer (50 mM Tris–HCl, pH 8.0, 1 mM EDTA, pH 8.0, 0.05% β-mercaptoethanol) and 1× protease inhibitor cocktail (Complete™, Roche, Mannheim, Germany). Usually, for each 100 mg of the fresh leaf tissue 100 µL of the above buffer was used. The extract was centrifuge at 13,000g for 15 min in 4°C. Total soluble protein concentration was determined by Bradford protein assay (BioRad, Chicago, IL, USA). The samples were used in ELISA, PAGE, and Western blot analyses. For mGM-CSF bioassay, the lyophilized leaves were ground in PBS buffer (150 mM NaCl, 8.1 mM Na2HPO4, 1.5 mM KH2PO4, 2.7 mM KCl) with 0.1% Tween-20. Usually for each 10 mg of the plant material 100 µL of the above buffer was used.
Western blot and ELISA
Protein samples (about 25 µg/well) were separated on 15% SDS-PAGE and transferred to a nitrocellulose membrane (Hybond-C Extra, Amersham Biosciences, Uppsala, Sweden). The mGM-CSF protein was detected using polyclonal rabbit antibodies against mouse GM-CSF (LabEmpire/BioShop Canada Inc, Burlington, Canada) and Western Breeze chromogenic Western blot immunodetection kit (Invitrogen, Basel, Switzerland).
A sandwich ELISA was performed using Mouse GM-CSF Quantikine Colorimetric Sandwich ELISA (R&DSystems, Abington, United Kingdom). The 1:2 serial dilutions of plant protein extracts were from 10−5 to 1.28 × 10−7. The commercially available (Biodesign, Saco, ME, USA) recombinant mGM-CSF protein produced in E. coli cells was used as a control. Protein concentration was determined using a standard curve for absorbance at 450 nm.
De-glycosylation
Enzymatic de-glycosylation of recombinant mGM-CSF extracted from transgenic plants was carried out using Endo H and PNGase F (New England BioLabs, Hitchin, United Kingdom) according to manufacturer’s instruction; however, the reaction time was longer (3 h). About 25 µg of the crude protein extract from transgenic plant producing mGM-CSF was used in the reaction. The samples were analyzed by Western blot.
mGM-CSF bioassay
The GM-CSF–dependent mouse lymphoblast cell line, FDC-P1 (American Type Culture Collection, Manassas, VA, USA), was used to determine the biological activity of plant-produced mGM-CSF. FDC-P1 cells were maintained according to the provider’s instruction as a suspension culture in Dulbeco’s modified Eagle’s medium (DMEM) (Mediatech, Manassas, VA, USA) supplemented with 10% fetal bovine serum and 0.5 ng/mL of recombinant mGM-CSF (Biodesign, Saco, ME, USA). Serial 1:2 dilutions (from 10−5 to 1.28 × 10−7) of the protein extracts (sterilized through 0.22 µm filter) from either the transgenic or non-transgenic plants were mixed with FDC-P1 cells (5 × 104 cells/well) in a total volume of 200 µL. As a reference standard, serial 1:2 dilutions of recombinant mGM-CSF (Biodesign, Saco, ME, USA) in a range of 4 to 0.03 ng/mL or DMEM medium alone were cultured with the cells. Each sample was assayed in triplicate. After 48 h of incubation at 37°C in 5% CO2, the viability of the cells was quantified using MTT (Research Products International, Mount Prospect, IL, USA) method (Hansen and others 1989).
Results
Tobacco transformation and characterization of transgenic plants
The essential elements of the plant expression cassette used for the production of mouse GM-CSF (mGM-CSF) in tobacco are shown in Figure 1A. Before proceeding to the stable tobacco transformation, the functionality of the expression cassette was confirmed in N. benthamiana leaves infiltrated with A. tumefaciens harboring recombinant plasmid (Fig. 1B). Subsequently, the mGM-CSF cassette was introduced into N. tabacum cv. LA Burley 21, which contains only 0.2%–0.5% of alkaloids on a dry weight basis (Legg and others 1970). This allows to use such plant material for animal feeding (or other mucosal administration) without a strong risk of getting undesired effects of alkaloid consumption. Forty regenerated kanamycin-resistant “plantlets” were chosen for further analysis after Agrobacterium-mediated transformation. The initial screening by PCR confirmed the presence of the full-length copy of mGM-CSF cDNA in genomic DNA of all plants examined. Additionally, the plants were also screened for the presence of the entire expression cassette using appropriate primers corresponding to the RbcS1 promoter and terminator regions, and the subsequent sequencing of the PCR products. This analysis confirmed the presence of the correct nucleotide sequence of the transgene in plant genomic DNA of all tested transformants. Chosen plants were transferred into soil and were subjected to further analysis. Total RNA isolated from the leaves of the selected plants was fractionated in agarose gels in denaturing conditions and transferred onto a nylon membrane. Hybridization with DIG-labeled (−) RNA of mGM-CSF transcript showed the presence of (+) RNA of mGM-CSF in all analyzed transgenic plants (data not shown). The transgene was successfully transcribed in all analyzed plants; however, the expression level varied in individual transformants.
Detection of N-glycosylated recombinant mGM-CSF
The predicted molecular weight of mGM-CSF after removal of the N-terminal leader peptide is below 15 kDa. The protein of the expected size was observed in the case of recombinant mGM-CSF produced in E. coli cells (Fig. 1B and 1C). However, the anti-mGM-CSF antibodies recognized 3 specific bands, one corresponding to 15-kDa protein and 2 others in a range between 17 and 22 kDa in both plant systems, in N. benthamiana leaves during transient expression (Fig. 1B) and in stably transformed N. tabacum (Fig. 1C). This observation, taking into consideration the fact of confirming of the correct nucleotide sequence of the transgene (see above), suggested that recombinant mGM-CSF might be post-translationally modified in plant cells. In fact, in silico analysis indicated that mGM-CSF contains 2 possible sites for N-glycosylation in the positions 66 and 75 of the predicted mature recombinant protein (without the N-terminal signal peptide). To verify this possibility experimentally, the crude protein extract from the transgenic tobacco was digested with 2 endoglycosidases, EndoH and PNGase F. The Western blot analysis following the reactions clearly showed that treatment with any of 2 used enzymes resulted in decreased molecular weight of the recombinant protein, which indicated that mGM-CSF is N-glycosylated in plants (Fig. 1D). The differences in relative bands intensities between the systems were observed and they might result from inefficiency of mGM-CSF glycosylation during transient expression test.
Production level and biological activity of recombinant mGM-CSF
The level of recombinant mGM-CSF was estimated by ELISA assay in several potted (grown in soil) T0 plants. As shown in Figure 2A, the detected level of mGM-CSF production varied from 8 to 19 µg/g of fresh weight of their leaf tissue.
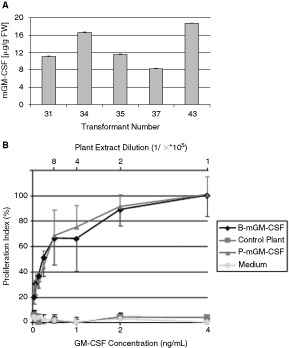
Determining the accumulation level (
The aim of subsequent experiments was verification of the biological activity of the recombinant mGM-CSF. As it is shown in Figure 2B, plant-derived mGM-CSF stimulated proliferation of the mouse lymphoblast cell line, FDC-P1, whose growth is dependent on GM-CSF. Neither extract from non-transgenic plants nor the medium that was not supplemented with mGM-CSF supported the proliferation of these cells. In contrast, the protein extract from transgenic plants as well as E. coli-derived mGM-CSF were able to maintain the growth of cell culture. The extract from mGM-CSF-producing transgenic plant-diluted 105-fold stimulates cells growth equally efficiently as E. coli-produced mGM-CSF supplemented at concentration of 4 µg/L of the cell culture medium.
Stability of recombinant protein upon storage of the lyophilized leaves
To estimate the best conditions for long-term storage of a large amount of plant leaf tissues containing recombinant mGM-CSF protein, we took into account both, a necessity to prevent protein degradation and a convenience of the storage. Samples of the fresh collected leaves were divided into 2 equal parts. One part was immediately frozen in the liquid nitrogen and kept at −70°C, while the second one was lyophilized and kept at room temperature. One month later, the freshly prepared protein extracts from the stored plant material were subjected to Western blot analysis. No difference was observed between samples derived from leaves kept at −70°C or at room temperature (Fig. 3). This result indicates the possibility of a convenient and simple storage of the lyophilized plant material containing recombinant mGM-CSF at room temperature.
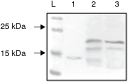
Analysis of the stability of plant-produced mouse granulocyte–macrophage colony-stimulating factor (mGM-CSF) upon lyophilization of the leaf tissues. Lane 1—Escherichia coli-derived purified mGM-CSF; lanes 2 and 3—protein extract from lyophilized leaves stored at 25°C and fresh leaves stored at −70°C, respectively; L—pre-stained protein molecular marker with indicated bands corresponding to 25 and 15 kDa.
Discussion
No previous attempts of a murine GM-CSF production in plants were reported; however, several groups worked on production of human GM-CSF in various plant-based systems, including suspension cultures, transgenic plants, and viral vectors. In stably transformed cell suspension cultures of tobacco or rice with the construct containing hGM-CSF cDNA under the control of the CaMV 35S promoter and a 5′-nontranslated region from the tobacco etch virus (TEV leader sequence), the production level was 250 and 150 µg/L of extracellular and intracellular hGM-CSF, respectively (James and others 2000). Performed comparative analysis showed that TEV leader sequence could increase protein production over 2-fold. Positive effects on recovery of secreted hGM-CSF were also observed after addition of stabilizing factors such as BSA or salt to the growth medium (James and others 2000). In other reported case, the expression of up to 783 µg/L was observed after addition of 5% (w/v) gelatin into tobacco cell suspension culture (Lee and others 2002). A much higher yield of recombinant protein was obtained in rice cell cultures, where the Ramy 3D promoter-driven expression of hGM-CSF was induced in sucrose-free conditions. In this system the cell cycle phase with the highest level of protease activity was avoided and the maximum production of hGM-CSF of 129 mg/L was observed (Shin and others 2003). Other researchers that focused on using rice cells suspension culture reported 16 mg/L of the cell culture volume as the maximum concentration of the extracellular hGM-CSF (Joo and others 2006). For the production of hGM-CSF in transgenic sugarcane 2 different promoters, Mubi-1 and Scubi9 (derived from the maize gene encoding polyubiquitin 1 and from the sugarcane gene encoding polyubiquitin 9, respectively), were examined. However, the accumulation of hGM-CSF reached only maximum 0.02% of TSP and no significant difference was observed between the used promoters (Wang and others 2005).
Herein, we report about 10-fold higher expression of mGM-CSF (about 0.3% of TSP) in the leaf tissues of tobacco. The higher level (of hGM-CSF accumulation) has been achieved only in the case of seed-targeted expression of transgenic rice (Sardana and others 2007; Ning and others 2008) and in the case of using plant virus-based system in N. benthamiana leaves (Zhou and others 2006). The previously published results of the comparative analysis indicated that RbcS1 promoter present in the ImpactVector is about 8-fold stronger than the commonly used 35S promoter from cauliflower mosaic virus (CaMV 35S) with the double enhancer (Outchkourov and others 2003). Additionally, the sequence encoding the KDEL signal was joined to the 3′-end of mGM-CSF cDNA. Due to the C-terminal KDEL sequence the secreted (containing the N-terminal leader sequence) protein is retained in the ER and does not go downstream into Golgi system. Such targeting could significantly improve the accumulation level of the recombinant protein because ER contains lower concentration and/or different proteases responsible for post-translational degradation. A high level of accumulation of mGM-CSF in plants was expected because of the above features. Indeed, the level of recombinant mGM-CSF protein achieved in our case is several-fold higher than the level reported previously for hGM-CSF produced in the leaves of transgenic plants (Sardana and others 2002, 2007; Wang and others 2005). Interestingly, a consistent difference in the N-glycosylation pattern of mGM-CSF was observed in transiently and stably transformed plants suggesting that one of the sites is less effectively recognized by the N-glycosylation machinery in the transient expression.
We have demonstrated that the plant-derived mGM-CSF protein was biologically active in proliferation assay based on the appropriate mouse cells. Plant extract diluted even 16 × 105-fold still was able to efficiently stimulate the growth of GM-CSF–dependent cells. This result seems to be a good prediction for the future usage of such plant material to develop veterinary application, for instance, as an adjuvant of experimental veterinary vaccines. Moreover, we have shown that the recombinant protein is N-glycosylated in plant cells and that lyophilization of the leaves might be a good approach to avoid its degradation upon storage of plant material at room temperature. The demonstrated stability of the recombinant mGM-CSF along with the low level of alkaloids in the tobacco cultivar used for the transformation in our experiment allows to apply plant material without need of its purification in future animal experiments to test the adjuvant properties of the plant-produced recombinant murine GM-CSF.
Finally, although usage of the ImpactVector system resulted in a relatively high accumulation level of the recombinant mGM-CSF in the leaves of transgenic tobacco, further experiments to improve the yield are necessary. These could include such approaches as, optimization of the codon usage or introducing the sequences that might provide better protection against degradation by plant proteases or facilitate storage.
Footnotes
Acknowledgments
We are grateful to Dr. Falko Steinbach (Veterinary Laboratories Agency, UK) for mGM-CSF cDNA and to Dr. Frantz Liszewska and Kacper Kazimierczuk for their input during initial cloning experiments. This work was supported by the Polish Ministry of Science and Higher Education (grants N302 061934 and 70/E-63/SN-0014/2007).
Author Disclosure Statement
No competing financial interests exist.