
Abstract
Several mechanisms have been postulated for the anticancer effects of tocotrienols. In this study, for the first time, the anticancer effect of tocotrienols is linked to increased expression of interleukin-24 (IL-24) mRNA, a cytokine reported to have antitumor effects in many cancer models. Tocotrienol isomers (α-T3, γ-T3, and δ-T3) and tocotrienol-rich fraction (TRF) inhibited the growth of the 4T1 murine mammary cancer cells (P < 0.05), with IC50 values 8.99, 4.79, 3.73, and 8.63 μg/mL, respectively. Tumor incidence and tumor load in TRF-supplemented BALB/c mice was decreased by 57.1% and 93.6% (P < 0.05), respectively. The induction of the IL-24 mRNA in the 4T1 cells by vitamin E decreased in the following order: δ-T3 > γ-T3 > TRF > α-T3 > α-T, which was similar to their antiproliferative effects. The IL-24 mRNA levels in tumor tissues of BALB/c mice supplemented with TRF increased 2-fold when compared with control mice. Increased levels of IL-24 have been associated with inhibition of tumor growth and angiogenesis. Treatment of 4T1 cells with TRF and δ-T3 significantly decreased IL-8 and vascular endothelial growth factor mRNA levels. Hence, we report that tocotrienols have potent antiangiogenic and antitumor effects that is associated with increased levels of IL-24 mRNA.
Introduction
T
Tocotrienols have also been shown to be involved in immunoregulation, antibody production (Nesaretnam and others 2004), and inducing resistance to implanted tumors (Nesaretnam and others 2004; Wada and others 2005; Wong and others 2009). Tocotrienol treatments have shown increased activity of natural killer cells in doses lower than that required for similar response by tocopherols (Ashfaq and others 2000). Other studies have demonstrated the involvement of tocotrienols in immunoregulation via the inhibition of CD74/Li, a variant chain associated with major histocompatibility complex class II (Nesaretnam and others 2004). Tocotrienols have also been shown to inhibit the action of CD59, a complement regulatory protein present on the cell surface, which can prevent the formation of the membrane attack complex on tumor cells (Nesaretnam and others 2004).
In the present study, we investigated the antiproliferative effects of palm tocotrienols and its individual fractions on the proliferation of 4T1 murine mammary cancer cells using in vitro assay systems as well as in a murine model. Immune regulatory effects were explored to further assess the role of tocotrienols in the modulation of immune genes, specifically the interleukin-24 (IL-24), which has been reported to be a unique cytokine with potent antitumor effects and often referred to as the magic bullet for cancer (Lebedeva and others 2003; Ramesh and others 2004; Chada and others 2005).
The IL-24, isolated during a subtractive hybridization carried out to screen for genes differentially upregulated following growth arrest and terminal differentiation of a human melanoma cell line, was known initially as melanoma differentiation-associated gene-7 (mda-7) (Chada and others 2004). The mda-7 cDNA encodes a novel 24-kDa protein that has homology to the IL-10 protein and the IL-10 family members such as IL-19, IL-20, IL-22, and IL-26 (Kotenko 2002; Chada and others 2004). Based upon its position within the IL-10 family cluster at 1q32.2 and cytokine-like properties, the mda-7 was designated by the Human Genome Organization as IL-24 (Caudell and others 2002). The IL-24 has been found to be associated with various tumor-suppressive mechanisms (Jiang and others 1996).
Materials and Methods
Reagents
Growth medium, fetal bovine serum, and antibiotics were from Gibco BRL. Zap-oglobin II lytic reagent and Isoton III were purchased from Beckman Coulter. Tocotrienol-rich fraction (TRF) was obtained from the Malaysian Palm Oil Board. Alpha-tocopherol (α-T) (95%) was obtained from Sigma Chemical Company. Individual tocotrienol fractions (α, γ, and δ) (97%) were obtained from Carotech Sdn Bhd.
Cell culture and treatment conditions
4T1 cells were purchased from American Type Culture Collection. Stock cells were grown in RPMI 1640 medium supplemented with 10% fetal calf serum (FCS) and 1% antibiotic penicillin–streptomycin in a humidified atmosphere of 5% CO2 in air at 37°C. All phytochemicals (TRF, α-T3, γ-T3, δ-T3, and α-T) that were used in treatments were dissolved in ethanol and were diluted accordingly in phenol red-free RPMI medium 1640 supplemented with 5% dextran–charcoal-treated FCS (DCFCS). Control cultures contained the same volume of ethanol. The culture medium was changed every 3–4 days.
Cell growth assay
Cells were trypsinized from the stock plates by treatment with trypsin/versene and added to an equal volume of phenol red-free RPMI medium 1640 supplemented with 5% DCFCS. The FCS was treated with dextran–charcoal to remove many of the growth factors, micronutrients, and macronutrients that may interfere with the growth assay. The 4T1 cells were resuspended to a cell count of 0.1 × 105 cells/mL with the use of a hemocytometer and plated in monolayer in 0.5 mL aliquots into 24-well plastic culture dishes. After 24 h, cells were treated with appropriate concentrations of the various phytochemicals. Cell count was performed after 12 days of culture. To perform cell count, the cells were washed twice with 0.9% NaCl to remove the nonadherent dead cells. Then, the cells were lysed in 0.5 mL of 2.5 mM HEPES buffer/1.5 M MgCl2. Following this, 2 drops of Zap-oglobin II lytic reagent was added and the mixture was incubated for 5–15 min. The nuclei released were suspended in Isoton III and counted on a Coulter counter with particle size set at >5 μm. All cell counts were carried out in triplicates on triplicate well contents.
Tumor inoculation and drug administration
Female BALB/c mice aged 4 weeks were purchased from the Institute of Medical Research. The mice were housed in an animal holding facility (AHF) at the Malaysian Palm Oil Board. Food and water were given ad libitum. The mice were allowed to acclimatize in the AHF for 1 week. The mice were randomly divided into an experimental group and a control group (10 animals in each group). The control was orally gavaged with the vehicle (soy oil). The mice in the experimental group were given 1 mg of TRF (dissolved in soy oil) by oral gavage (3 times a week) at 2 weeks before the start of tumor induction. These mice continued to receive the same supplementation for the whole duration of the experiment. A cell suspension containing 1 × 104 4T1 cells/mL in RPMI was prepared. The tumor cell suspension (100 μL) was inoculated into the right mammary fat pad of each mouse. The inoculation site was palpated every 2 days. Tumors were measured by calipers in 2 perpendicular diameters and their volume was estimated using the following formula: V = 0.52 × L2 × W (where V is volume, L is length, and W is width), which was previously described (Wong and others 2009). The mice in the control and experimental groups were weighed. The TRF treatment did not produce any significant change in the growth rate or final body weight in comparison with the group of control animals (data not shown). The experiment was terminated at 45 days after inoculation of tumor because the tumor load in the control mice became too big. All the animals were sacrificed. The tumors were excised from the animals and snap-frozen in liquid nitrogen before RNA extraction. The present study was approved by the Research and Ethics Committee of the International Medical University, and animal care and handling were strictly according to the guidelines provided by this committee.
RNA isolation
Total RNA was extracted from 4T1 mouse mammary cancer cells treated with different concentrations (2–20 μg/mL) of TRF, α-T, δ-T3, α-T3, and γ-T3 for 72 h and untreated cells using Trizol (Invitrogen) according to the manufacturer's protocol. The RNA clean-up and DNAse digestion were performed using the RNeasy® Mini kit (Qiagen). The RNA was quantified and assessed for purity by measurement of optical density at 260 and 280 nm using a UV fiberoptic spectrophotometer (IMPLEN NanoPhotometer) and was qualitatively assessed by measurement of relative 28S and 18S ribosomal band intensities, using the BioRad Experion Automated Electrophoresis System. Aliquots were kept frozen at −80°C until further processing.
Real-time reverse transcriptase–polymerase chain reaction
The relative expression of IL-24 was analyzed by quantitative reverse transcriptase–polymerase chain reaction (RT-PCR) with β-actin as the internal control. The following pairs of reverse and forward primer sets were used: IL-24, 5′ TGCTGAGAGCTGTTACCTTG 3′ and 5′ ACTGGGTTGCAGTTGTG ACATGATGA 3′ (GenBank accession No. NM_053095.2); and β-actin, 5′ AGAAGGATCCTATGTGGGCG 3′ and 5′ CATGTCGTCCCAGTTGGTGAC 3′ (GenBank accession No. EF_095208.1). The RT-PCR mixture contained 15 μL of 2 × reaction buffer, 5 μL each of RNA, 0.125 μL each of forward and reverse primers (100 μM), and 0.5 μL of RT-Platinum Taq in a final volume of 25 μL. The reaction was done at 50°C for 30 min, 95°C for 5 min, 95°C for 40 cycles of 15 s each, 60°C for 30 s, and 60°C for 30 s, followed by a melt curve step. RT-PCRs were performed in triplicates. We used the 2−ΔΔCt method with the tested primers to calculate relative expression levels of the transcripts.
QuantiGene assay
QuantiGene assay as described by company protocols (Panomics) was used for direct quantification of IL-24, IL-8, and vascular endothelial growth factor (VEGF) levels in RNA from 4T1 cells treated with TRF, δ-T3, and α-T and compared against untreated control cells. The assay employs a sandwich nucleic acid hybridization method that uses branch DNA molecules to amplify signal from captured target RNA. The assay was carried out as described in QuantiGene 2.0 Reagent System User Manual (Panomics). About 100 ng RNA/well was loaded into 96-well plate for detection of target probe and 1 ng RNA/well was loaded for detection of β-actin. Working probe set was prepared for each target and capturing of target RNA was carried out at 55°C overnight. Following the overnight hybridization, signal amplification and detection steps were carried out using target-specific preamplifiers, amplifiers, and label probes. Signal detection was carried out by addition of chemilumigenic substrate that generates a luminescent signal that is proportionate to the amount of target mRNA present in the sample. Luminescent signal was detected using the GloRunner Microplate Luminometer at a wavelength of 550 nm.
Statistical analysis
Data are presented as the mean ± standard deviation, where n is the number of mice used in the study. In all animal experiments, samples were assessed individually (ie, not pooled). Either one-way analysis of variance (followed by Tukey's pair-wise comparisons) or Student's t-test was used to determine the significance between control and treated samples, with P < 0.05 set as the level of acceptable statistically significant difference unless stated otherwise.
Results
TRF and individual fractions of tocotrienols inhibit the growth of 4T1 mouse mammary cancer cells in vitro
Treatment with TRF and individual tocotrienols (α, γ, and δ) for 12 days inhibited the growth of 4T1 mouse mammary cancer cells (Fig. 1). The inhibitory effect on cell growth followed the following order: α-T3 < TRF < γ-T3 < δ-T3. Treatment with δ-T3 for 12 days significantly inhibited 4T1 cell growth from as low as 2 μg/mL (P < 0.001). This means that δ-T3 could exert a very potent inhibitory effect even at low concentrations. Complete growth suppression was achieved at 6 μg/mL of δ-T3 and γ-T3, with cell numbers falling below the plating density. The γ-T3 also significantly inhibited cell growth from 2 μg/mL (P < 0.001), with cell numbers falling below the plating density at 10 μg/mL, whereas TRF showed significant inhibitory effects from 4 μg/mL (P = 0.006). In contrast to these results with tocotrienols, treatment of 4T1 cells with a similar concentration (2–20 μg/mL) of α-T did not show any inhibitory effect. The TRF has a standardized composition of 32% α-T, 25% α-T3, 29% γ-T3, and 14% δ-T3. The δ-T3 and γ-T3 showed the highest inhibitory effects in the cell growth assays. As described in Fig. 1, the inhibition concentrations at 50% (IC50) values of α-T3, TRF, γ-T3, and δ-T3 were 8.99, 8.63, 4.79, and 3.73 μg/mL, respectively, whereas the IC50 value for α-T was beyond the test range, ie, >20 μg/mL.
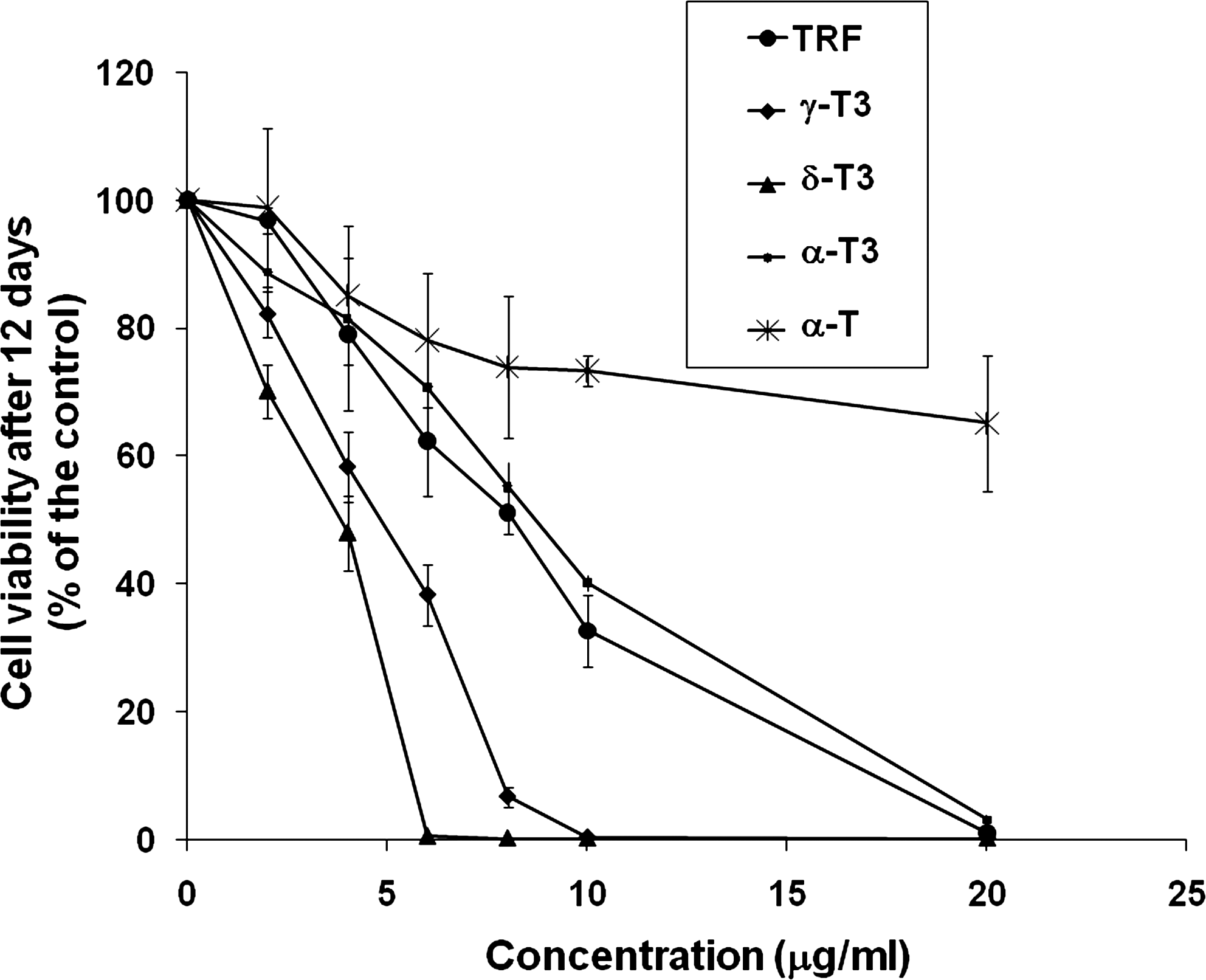
Effects of TRF, individual tocotrienols (α-T3, γ-T3, and δ-T3), and α-tocopherol (α-T) on 4T1 murine mammary cancer cell growth. Cells were treated for 12 days, and viable cells were determined at the end of treatment using Coulter counter. Error bars represent the standard deviation of triplicate wells. IC50 values for each treatment were calculated from dose–response curves using DPlot. IC50 values obtained were as follows: TRF (8.63 ± 0.53), α-T3 (8.99 ± 0.34), γ-T3 (4.79 ± 0.53), δ-T3 (3.73 ± 0.39), and α-T (>20 ± 0). The values are means ± standard deviation of cell numbers obtained from triplicate wells. TRF, tocotrienol-rich fraction.
TRF supplementation suppresses mammary tumor growth in mice
The in vivo effect of TRF on the mammary tumor growth in BALB/c mice was investigated by examination of tumor formation for a period of 45 days in TRF-fed and vehicle control mice. Tumor incidence was significantly reduced by 57.1% with supplementation of 1 mg TRF compared with vehicle control (P < 0.01; Fig. 2A). Supplementation with TRF was also shown to significantly reduce the tumor load in the mice by 93.6% when compared with the control (P < 0.05; Fig. 2B).

Cumulative tumor incidence
Tocotrienols upregulate the expression of IL-24 in 4T1 mouse mammary cancer cells
IL-24 overexpression has been linked with increased tumor protective effects via induction of apoptosis, angiogenesis, and other antitumor mechanisms (Wong and others 2009). We investigated the expression of IL-24 in 4T1 cells treated with a range of concentrations (2, 4, 6, and 8 μg/mL) of TRF, δ-T3 (because δ-T3 exhibited highest potency in inhibition of tumor cell growth), and α-T using the QuantiGene assay. As shown in Fig. 3, all of the tested concentrations of δ-T3 significantly (P < 0.05) increased the expression of the IL-24 transcript from concentration as low as 2–8 μg/mL in a dose-dependent manner. The TRF increased the expression of the IL-24 mRNA from concentrations higher than 4 μg/mL when compared with untreated control cells (Fig. 3). For all the tested compounds, the highest level of IL-24 was achieved at 6 μg/mL. In addition, for all the tested compounds, the expression of IL-24 began to decline in the same order when the cells were treated with 8 μg/mL or higher, ie, δ-T3 > TRF > α-T; however, the differences observed between 6 and 8 μg/mL treatment were not significant (P > 0.05). We further confirmed the results using real-time RT-PCR on the RNA extracted from 4T1 mouse mammary cancer cells treated with 8 μg/mL of TRF, α-T3, γ-T3, and δ-T3 to determine if the expression of IL-24 was corresponding to the inhibitory effects of the isomers. The concentration was chosen based on the IC50 values of these compounds. We compared the IL-24 mRNA expression levels with the cells treated with α-T and untreated vehicle control cells. Figure 4 shows the expression levels of IL-24 to be highest in δ-T3-treated cells followed by γ-T3, TRF, and α-T3 (P < 0.05), whereas the α-T-treated cells did not show significant increase (P > 0.05).
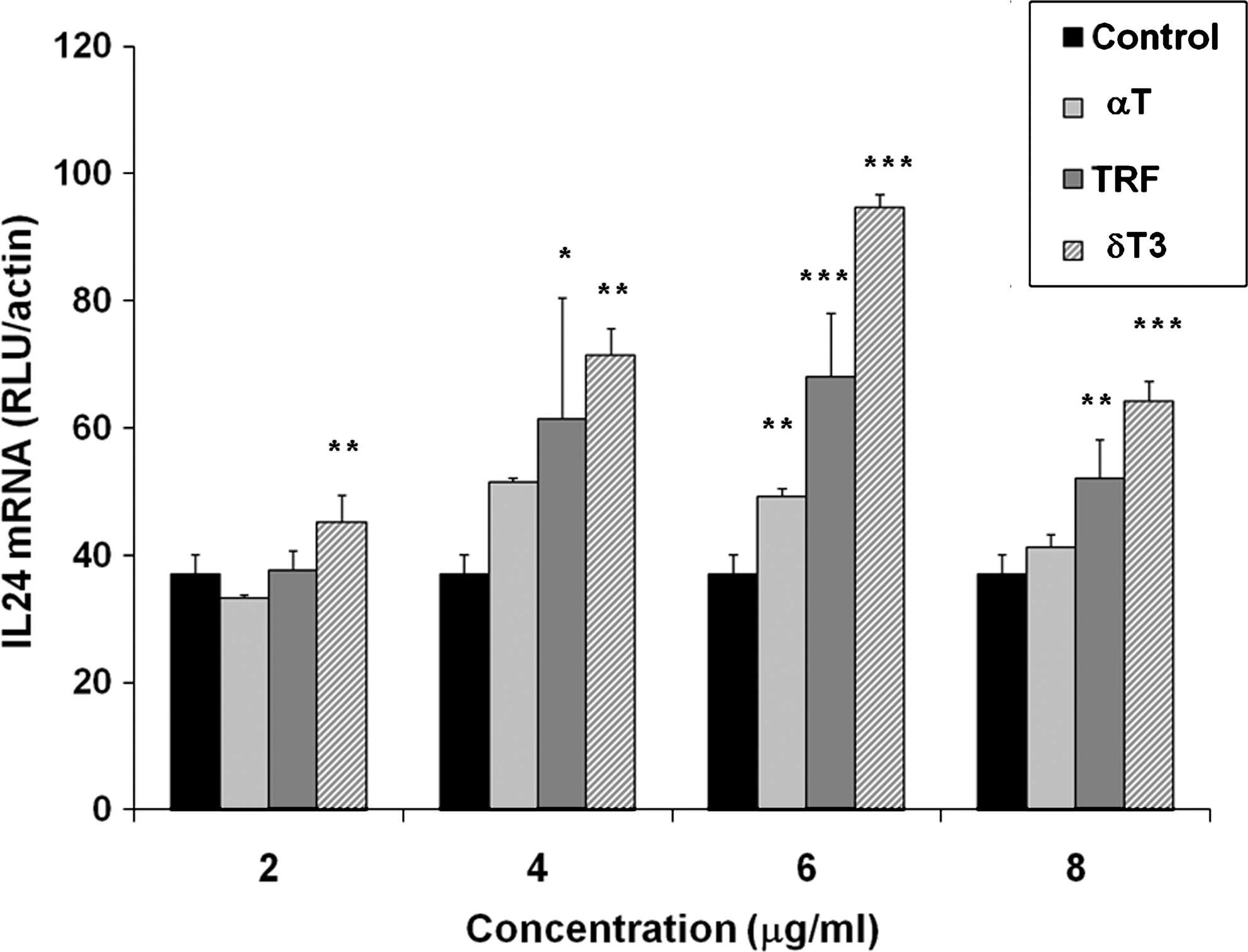
Expression of IL-24 mRNA in 4T1 murine mammary cancer cells treated with 2, 4, 6, and 8 μg/mL TRF, δ-T3, and α-T and untreated control cells. *P < 0.05, **P < 0.01, and ***P < 0.001 represent significant differences between IL-24 mRNA levels in treated cells and untreated control cells. IL, interleukin; TRF, tocotrienol-rich fraction.

IL-24 expression levels after treatment with 8 μg/mL α-T, TRF, α-T3, γ-T3, and δ-T3. Samples were prepared in triplicates and mean values of target calculated at the end of the polymerase chain reaction using the relative standard curve method to determine relative quantification; *a statistically significant value with P < 0.05. IL, interleukin; TRF, tocotrienol-rich fraction.
TRF increases the expression of IL-24 mRNA in tumor tissues
The expression of the IL-24 mRNA in tumor tissues of TRF-supplemented BALB/c mice was determined using QuantiGene assay. Tumors from vehicle-supplemented mice were used as controls. The tumor excised from the TRF-supplemented mice had significantly (P < 0.05) higher expression of the IL-24 mRNA compared with tumors from control animals (Fig. 5). The expression of IL-24 mRNA in tumor from experimental mice was 2-fold higher when compared with the control (Fig. 5).
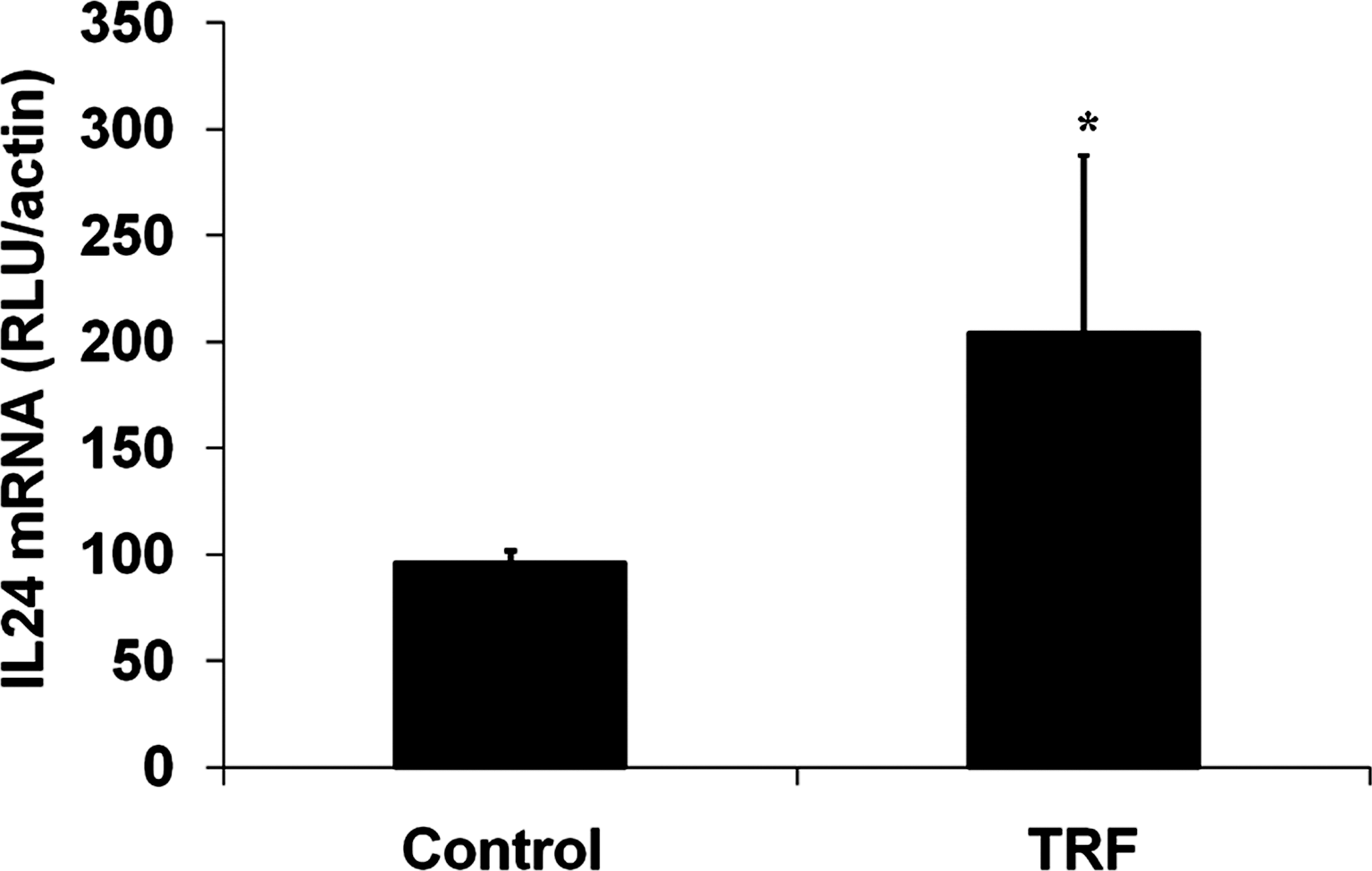
Expression of IL-24 mRNA in tumor tissues of TRF-supplemented mice compared with vehicle control mice. *P < 0.05 represents significant difference between IL-24 mRNA level in TRF-supplemented mice and control mice. IL, interleukin; TRF, tocotrienol-rich fraction.
Tocotrienols increase the expression of IL-24 mRNA and decrease the expression of IL-8 and VEGF in 4T1 murine mammary cancer cells
As IL-24 and tocotrienols have been reported to inhibit tumor angiogenesis, we determined the expression levels of some angiogenic markers such as IL-8 and VEGF in 4T1 murine mammary cancer cells treated with 8 μg/mL TRF, δ-T3, and α-T and untreated control cells using the QuantiGene assay. Figure 6 shows that the levels of both IL-8 (P < 0.05) and VEGF (P < 0.01) were significantly reduced in both the TRF- and δ-T3-treated 4T1 cells when compared with α-T-treated and control cells. In contrast, the expression of IL-24 significantly increased in TRF (P < 0.01) and δ-T3 (P < 0.001)-treated cells when compared with α-T-treated and control cells (Fig. 6). The IL-24 expression was highest in δ-T3-treated cells.

Expression of IL-24, IL-8, and VEGF mRNA in 4T1 murine mammary cancer cells treated with 8 μg/mL TRF, δ-T3, and α-T and untreated control cells. *P < 0.05, **P < 0.01, and ***P < 0.001 represent significant differences between mRNA levels in treated cells and untreated control cells. IL, interleukin; TRF, tocotrienol-rich fraction. VEGF, vascular endothelial growth factor.
Discussion
Tocotrienols have previously been reported for their potent anticancer properties in various cancer models both in vivo and in vitro (Sylvester and Theriault 2003; Nesaretnam 2008). In the present study, we further investigated the potent effects of individual fractions of tocotrienols, especially δ-T3, and compared its effect to TRF and α-T using an in vitro and in vivo test system. δ-Tocotrienol showed the highest potency in inhibiting 4T1 murine mammary cancer cell growth and this is followed by γ-T3, TRF, and α-T3, whereas α-T did not exhibit any inhibitory effect at the same concentrations. These observations are consistent with previous observations made on PC-3 prostate cancer cells where δ-T3, γ-T3, and TRF were found to be potent inhibitors of PC-3 cell proliferation in vitro, whereas α-T3 and α-T had no significant effect (Nesaretnam and others 2008). Similar studies have shown that tocotrienols, but not α-T, suppressed the growth of human breast and colorectal cancer cells (Nesaretnam and others 1995, 1998, 2000; Eitsuka and others 2006). In another study, it was shown that the anticarcinogenic property of tocotrienols could work in synergy with tamoxifen as the combination of these 2 treatments was found to be more potent than tamoxifen alone (Guthrie and others 1997).
The antitumor activity of tocotrienols may not be associated with just their antioxidant activity only (Nesaretnam 2008). Recent findings have shown that the antitumor action of tocotrienols may be due to several mechanisms such as antiproliferation (Wada and others 2005; Srivastava and Gupta 2006), antiangiogenesis (Nakagawa and others 2007; Wong and others 2009), anti-inflammatory (Wu and others 2008; Yam and others 2009), initiation of apoptosis (Shah and others 2003; Ahn and others 2007), and enhancing immunological functions (Gu and others 1999; Hiura and others 2009).
The higher inhibitory effects displayed by tocotrienols, especially δ-T3, over α-T could be attributed to the unsaturated phytyl side chain of the tocotrienols. The side chain is primarily involved in the production of isoprenoid intermediates that have been proven to exhibit anticancer properties by suppressing the mevalonate pathway (Elson 1995) and initiate cellular apoptosis by reducing ras and lamin B processing (Elson and others 1999). It has also been suggested that the unsaturated side chain, resulting in less planar molecular conformation, allowed tocotrienols to permeate the membrane lipid bilayer easily into target cells (McIntyre and others 2000).
In the present study, we also found that mice supplemented with TRF significantly reduced tumor incidence and tumor volume when compared with vehicle control mice. This observation further confirms that the antitumor effects of tocotrienols are achievable in an in vivo system as previously reported (Nesaretnam and others 2004; Wong and others 2009). In another study that compared the effects of tocotrienols with α-T, intraperitoneal administration of α-T3 and γ-T3, but not α-T, showed a life-prolonging effect in mice with transplanted tumors (Lebedeva and others 2005).
Many studies have reported that IL-24 can facilitate tumor regression, making this cytokine as the new focus area for anticancer research (Su and others 1998; Cunningham and others 2005). The IL-24 has been shown to suppress growth in vitro and induce apoptosis in diverse human cancer cells without inducing harmful effects in normal cells (Chada and others 2004). These findings indicate that IL-24 may act as a novel tumor suppressor protein. To date, there are no studies that show that tocotrienols can modulate the expression of IL-24. One of the goals of the present study is to determine if the antitumor effects observed with tocotrienols can be linked to the IL-24 pathway. In the present study, we have shown that tocotrienols, ie, TRF, α-T3, γ-T3, and δ-T3, significantly increased the expression of IL-24 mRNA in murine mammary cancer cells in a manner similar to their antiproliferative effects. A similar effect was not observed with α-T. We believe that this is the first report that links tocotrienols to the IL-24 pathway.
We also observed a similar increase in the IL-24 transcript in tumor excised from TRF-supplemented mice where the expression of the IL-24 mRNA was increased 2-fold compared with control. This finding complements the observation of decreased tumor incidence and tumor load in TRF-supplemented mice. The IL-24 has previously been shown to inhibit growth of lung tumor and metastases in vivo (Su and others 1998; Saeki and others 2002; Fisher and others 2003; Chada and others 2004; Ramesh and others 2004). Advanced application of IL-24 in cancer immunotherapy has progressed to a phase 1 clinical trial, which has promising results on the use of this cytokine as an immunotherapeutic agent for cancers (Bar-Eli 1999).
As mentioned earlier, tocotrienols have been reported to inhibit tumor angiogenesis by inhibiting proliferation, migration, and vascularization and reducing expression of VEGF in human endothelial growth factor cells as well as in vivo using mouse models (Nakagawa and others 2007; Wong and others 2009). The present study reconfirms our previous observations (Wong and others 2009) that TRF and δ-T3 exhibit antiangiogenic effects by downregulating the expressions of VEGF in tumor cells. In the present study, we further report that there is reduced expression of VEGF and IL-8 in 4T1 cells cultured with TRF and δ-T3. IL-8 is a powerful chemotactic factor that can provide a growth advantage to tumor cells (Murphy and others 2005). A parallel increase in IL-8 and its receptors has been associated with proliferation and microvessel density in prostate cancer (Ferrara and others 1993). Further, there are higher levels of this cytokine in serum of patients with metastatic prostate cancer (Ferrara and others 1993). The expression of VEGF has been shown to increase tumor growth and angiogenesis (Su and others 2005; Zhang and others 1995).
Interestingly, the present study shows that in 4T1 cells treated with TRF and δ-T3, there is a significant decrease in the expression of transcripts for proangiogenic factors such as IL-8 and VEGF and a concurrent increase in levels of IL-24, which has antiangiogenic activity (Gupta and others 2006). We were not able to show a similar effect with α-T at the same concentrations. Hence, we propose that the antiangiogenic effects of tocotrienols may be linked to the increased levels of IL-24 as there are studies that link the tumor-suppressive mechanisms of IL-24 to antiangiogenic activity (Mhashilkar and others 2001; Gupta and others 2006). In addition, gene transfer studies into human xenografts using Ad-mda-7 or mda-7/IL-24 plasmids have resulted in inhibition of tumor growth via antiangiogenic activity (Gupta and others 2006).
Correlation between the antitumor activities of tocotrienols and IL-24 are not only observed in the antiproliferative and antiangiogenic mechanisms of these 2 compounds but also in their effects in inducing apoptosis in a range of cancer cell types. Adequate evidence has accumulated to demonstrate that overexpression of the mda-7 gene, using either plasmid vectors or a replication-defective adenovirus, results in growth suppression and induction of apoptosis in a broad range of cancer cells (Jiang and others 1996; Su and others 1998; Madireddi and others 2000; Mhashilkar and others 2001). It has been reported previously that TRF and δ-T3 induce apoptosis in prostate cancer cells via the Fas receptor and FasL signaling pathway (Nesaretnam and others 2008). We propose that the apoptosis-inducing effects observed in tocotrienols may possibly be due to increased levels of IL-24; however, more studies are required to confirm these observations.
Although extensive research is underway to explain the actual pathway underlying the multiple effects of tocotrienols, no exact mechanism has been identified as to how tocotrienols work. Summary of the findings from the present study shows that most of the antitumor effects exhibited by tocotrienols closely relate to the high expression of IL-24 mRNA, which plays a crucial role in tumor suppression. Thus, we conclude that a possible mechanism of the antitumor effects of tocotrienols is via increased expression of IL-24.
Footnotes
Acknowledgments
The authors thank the Malaysian Palm Oil Board and the International Medical University of Kuala Lumpur for funding this study.
Author Disclosure Statement
No competing financial interests exist.