
Abstract
The interferon (IFN)-α response of pigs to the stressing event of early weaning was investigated in a field trial. All the animals under study remained healthy and tested negative for common viral infections. However, a low-titered IFN-α response was detected in many sera by a bioassay on Madin-Darby bovine kidney (MDBK) cells on day +6 after weaning. Porcine IFN-α was unambiguously identified by a neutralization assay on a pool of IFN-α-positive sera. By gel filtration chromatography, the antiviral activity of sera on MDBK cells could be traced back to 3 components of apparent molecular mass 27/18/<14 kDa. Additional components of apparent molecular mass 58 and 41 kDa were revealed by ELISA in Nonidet P-40 lysates of peripheral blood mononuclear cells (PBMC). Also, many pigs tested positive in flow cytometry assays on PBMC for intracellular IFN-α. The expression of porcine IFN-α genes was investigated by reverse transcriptase (RT) real-time polymerase chain reaction at days −1, +6, and +12 with regard to weaning in PBMC of 9 piglets. On days −1 and +12, IFN A5, A6, A12, as well as (in fewer pigs) A1, A7, A11, and A2 genes were shown to be expressed. On the contrary, none of the above genes was expressed on day +6, when plenty of pig sera were IFN-α-positive. Our results indicate that weaning causes the release of IFN-α and the transient shut-off of the corresponding gene transcriptions in PBMC. Interestingly, only IFN A9 gene transcription was shown in vitro to be virus induction-dependent.
Introduction
O
To confirm a major role of IFN-α in the pigs' adaptation strategy to the weaning stress. To determine the influence of the weaning age on the IFN-α response. To reveal the IFN-α genes and molecules involved in such a response and to discriminate between constitutive, weaning-associated and virus-induced IFN-α responses.
Materials and Methods
Animals
The study was carried out on 39 Landrace × Large White, randomly selected piglets of 4 litters located in the same pen of a farrow-to-finish herd in Brescia Province, Italy. The herd was classified as “healthy” due to the previous clinical findings, the very low levels of environmental stressors evidenced in repeated inspections, and the excellent farm production figures at the beginning of the trial. On the same day, group 1 swine (2 litters, 19 piglets) were weaned at 28 days of age, whereas group 2 swine (2 litters, 20 piglets) were weaned at 22 days of age. All piglets were housed in the same pen and given the same starter feed containing amoxicillin (400 ppm), colistin (120 ppm), and zinc oxide (2,000 ppm) over the first 10 days after weaning. Pigs were clinically inspected, and blood samples in vacuum tubes with sodium heparin and without anticoagulant were collected at days −1, +6, +12 with regard to weaning.
We also included in our study 9 Landrace × Large White pigs, reared in the Experiment Station located at IZSLER, Brescia, Italy. Clinical inspections and blood samplings were carried out by an appointed veterinary officer under the control of the Animal Health and Welfare Division of the Italian National Veterinary Services.
Blood samples
Blood samples without anticoagulant were kept at room temperature for 2 h, refrigerated for 30 min, and then centrifuged at 4°C for 15 min at 2,000 rpm; serum was separated and stored in aliquots at −80°C for cytokine and polymerase chain reaction (PCR) assays. Heparinized blood samples were diluted 1:2 with RPMI 1640 medium and centrifuged on Histopaque 1.077 (Sigma-Aldrich) at 1,100 g for 25 min, 20°C. PBMC were separated, washed twice with RPMI 1640 medium, and counted. Cells were immediately frozen in aliquots at 4 × 106 viable cells/mL in RPMI 1640 medium +40% fetal calf serum (FCS) +10% dimethyl sulfoxide (DMSO). Later on, they were thawed and employed in the analyses on IFN-α genes and intracellular protein reported hereunder. In other experiments, PBMC were also cultured for in vitro induction of IFNs (see relevant section hereunder).
Serological and molecular tests for viral infections of swine
The most common viral infections of pigs [sustained by Pseudorabies virus (PRV), Porcine Reproductive and Respiratory Syndrome virus (PRRSV), Influenza viruses, Porcine Circovirus 2 (PCV2)] were investigated by serological and molecular assays. These were employed on group 2 serum samples collected on days −1/ +12 (Ab assays) and +6 (PCR assays) with regard to weaning. Antibody to PRV (Suid Herpesvirus 1) was investigated by the HerdChek PRV gpI ELISA test kit (IDEXX Laboratories Inc.) according to the manufacturer's directions. The antibody (Ab) response of pigs to PRRSV (swine Arterivirus) was evaluated by another commercial kit (Herdchek Porcine Reproductive and Respiratory Syndrome Antibody test Kit; IDEXX Laboratories Inc.) according to the manufacturer's directions. Antibody to PCV2 was measured by a competition ELISA with a couple of monoclonal antibodies (mAbs) directed to PCV2 nucleoprotein (ORF2), as reported in a previous study (Sala and others 2000). Antibody to Influenza viruses H1N1, H1N2, and H3N2 was investigated by as many hemagglutination inhibition tests as previously described (Swenson and others 2008). Nucleic acids were extracted from serum samples by QIAamp® DNA mini kit (Qiagen) and High Pure Viral RNA kit (Roche), respectively, and used in tests for DNA and RNA viruses of swine. TaqMan® probe Real-Time PCR was performed for detection of PRRSV (Revilla-Fernandez and others 2005) and PCV2 (Chung and others 2005). Real-time PCR reactions were carried out in a CFX96™ Real-Time System (Bio-Rad). The fluorescence threshold limit was automatically set. Also, a PCR assay was performed for detection of PRV as previously described (Katz and Pedersen 1992). PCR amplification was performed by a Geneamp 2400 PCR system (Applied Biosystems) and detected by electrophoresis on a 2% agarose gel stained with ethidium bromide (50 μL/100 mL) in 1 × TAE buffer; DNA bands were visualized by a UV transilluminator (Chemidoc™; Bio-Rad).
Analyses of cytokines
Swine IFN-α was measured in serum samples by a cpe-inhibition assay on MDBK cells with Vesicular Stomatitis Virus (VSV) (Meager 1987b); the test was calibrated with a preparation of porcine recombinant IFN-α1 (PBL Biomedical Laboratories, cat. 17100-1). The units of this preparation are determined with regard to the international reference standard for human leukocyte IFN (Ga-902-530) provided by National Institutes of Health. Briefly, serial 4-fold dilutions of samples in tissue culture medium were reacted in duplicate in 96-well microtiter plates with 45,000 MDBK cells/well overnight at 37°C. Next day, cell monolayers were washed and infected with VSV. About 24 h later, cells were stained with neutral red (0.01% final), and IFN titers were determined after reading OD550 of sample, VSV-infected and uninfected control wells in an ELISA reader. The MDBK cells in current use in our laboratory show a much higher sensitivity to swine IFN-α as compared with swine IFN-γ, which is not a confounding element in the presence of low-titered antiviral activities in biological fluids of healthy pigs. This assay for porcine IFN-α in serum had been also validated in a previous study (Artursson and others 1989). Identification of the cytokine in pig sera was performed by a neutralization assay on MDBK cells of IFN α-positive sera (Meager 1987a) using mAb G16 to porcine IFN-α1 (Serotec, cat. MCA1935Z). Intracellular IFN-α in swine PBMC (3 × 107/mL) after lysis in 1.0% Nonidet P-40 (NP-40)/NaCl 150 mM/Tris 50 mM pH 8.0 + protease inhibitors (Protease Inhibitor Cocktail set III, cat. 539134; Calbiochem) was investigated by an ELISA assay with catcher mAb K9 and tracer, biotin-labeled mAb F17; these monoclonals have been used for a long time in high-sensitivity detection assays of porcine IFN-α (Diaz de Arce and others 1992). Cytokine concentrations were determined from a standard curve created with the above preparation of porcine recombinant IFN-α1 (PBL Biomedical Laboratories, cat. 17100-1). The aforementioned cell culture and ELISA assays for IFN-α were also employed on supernatants of cultured swine PBMC under different experimental conditions. IFN-γ was measured by sandwich ELISA. Briefly, Maxi Sorp NUNC ELISA plates were adsorbed with anti-swine IFN-γ capture mAb P2F6 (Thermo Scientific) at 5 μg/mL in 0.1 M carbonate/bicarbonate buffer pH 9.6 and incubated overnight at 4°C. After blocking with 4% bovine serum albumin in phosphate-buffered saline (PBS) (assay buffer), plates were washed thrice with 0.2% Tween 20 in PBS (PBS-Tween). Then, 50 μL/well of undiluted test samples, recombinant porcine IFN-γ at 500 pg/mL (Thermo Scientific, positive control, cat. 1857784), and assay buffer (blank control) were added in duplicate and incubated at room temperature for 1 h. Then, 50 μL/well of biotinylated, anti-swine IFN-γ mAb MP701B (Thermo Scientific) at 2 μg/mL in assay buffer was added, and plates were further incubated for 1 h at room temperature. After 3 washings with PBS-Tween, swine IFN-γ was revealed by horseradish peroxidase (HRP)-conjugated streptavidin and ortho-phenilenediamine + H2O2 as substrate of the color reaction. This was blocked after 15 min by adding 50 μL/well of 2N H2SO4. Plates were spectrophotometrically read at 492 nm. Cytokine concentrations were determined from a standard curve created with the above porcine recombinant IFN-γ.
Gel-filtration chromatography
A 2.5 × 110-cm, Sephadex G-75 column (GE Healthcare, cat. 17-0051-01) in PBS pH 7.2 was packed according to the manufacturer's directions by an ECONO-SYSTEM low-pressure chromatography apparatus (Bio-Rad Laboratories Inc.). Next, it was calibrated with the following molecular weight standards: bovine serum albumin (66 kDa), Ovoalbumin (45 kDa), IgG light and heavy chains (25 and 55 kDa), Lysozyme (14 kDa), and Neutral Red (0.4 kDa, as total volume marker). Serum samples and postnuclear fractions (10,000 rpm, 20 min) of NP-40 cell lysates from swine PBMC were applied to the calibrated column, which was run at 12 mL h−1 at room temperature. Each 0.5-mL fraction was refrigerated and supplemented with 50 μL of FCS + 15 μL of an antibiotic mixture (penicillin, streptomycin, and amphotericyn B, at concentrations of 500 IU/mL, 250 IU/mL, and 0.5 mg/mL, respectively). Next day, fractions were centrifuged (10,000 rpm, 20 min, 5°C) and supernatants were checked for IFN-α by the bioassay on MDBK cells or sandwich ELISA for serum samples and NP-40 lysates of PBMC, respectively.
Flow cytometry
Staining of cells for IFN-α was carried out according to an established procedure (Walravens and others 2002), with minor modifications. Briefly, frozen PBMC were rapidly thawed at 38°C and fixed in 4% paraformaldehyde at 4°C for 10 min. Cells were washed with ice-cold PBS and permeabilized with PBS-10% FCS-0.1% saponin (PBS-FCS-S). Then, they were incubated with anti-porcine IFN-α1 mAb G16 (1 μg/mL in PBS-FCS-S) or buffer only (control) for 30 min at room temperature, washed, and again incubated in PBS-FCS-S containing goat, F(ab′)2, anti-mouse IgG1-Phycoerythrin (Southern Biotechnology Associates, cat. 1072-09) for 30 min at room temperature. After washing once in PBS-FCS-S and twice in PBS, mAb-treated and control cells were resuspended in PBS-4% paraformaldehyde at 4°C. Samples were analyzed on a Bryte HS flow cytometer (BIO-RAD Microscience) using Win-Bryte software. The typical forward and side scatter, lymphocyte/monocyte gate was set to exclude dead cells from the analysis. The percentage of positive cells beyond the threshold FL2 fluorescence channel was assessed in each sample on 25,000 events and compared between mAb-treated and control cells. Under our usual working conditions (1%–2% of unspecific staining and comparison between 25,000 mAb-treated and as many control cells), a net 0.3% prevalence of IFN-α-positive cells is statistically significant (Fischer exact test, P < 0.05).
IFN-α gene expression and primer design
Total RNA was extracted from 8 × 106 swine PBMC using RNeasy® mini kit (Invitrogen) following the manufacturer's directions. The concentration and integrity of the extracted RNA were evaluated by a Qubit™ Fluorometer (Invitrogen) using the Qunt-iT RNA Assay Kit (Invitrogen) according to the manufacturer's protocol. The expression of 11 porcine IFN-α genes was then determined by reverse transcriptase (RT) real-time PCR, using the primer sets described by Cheng and coworkers (2006) (Table 1). Each gene sequence was obtained from the NCBI data base (
F, forward primer; R, reverse primer.
In vitro induction of IFN α and γ from swine PBMC
Venous blood of four, 70- to 80-day-old Landrace × Large White pigs (IZSLER Experiment Station) were collected in heparinized tubes; blood was diluted 1:2 with RPMI 1640 medium and centrifuged on Histopaque 1.077 (Sigma-Aldrich) as indicated above. PBMC were resuspended at 4 × 106 cells/mL (final) in RPMI 1640 medium + 10% FCS; 2 mL/well were seeded in 12-well Costar tissue culture plates. PBMC were either kept as unstimulated controls or treated for 2 h with recombinant porcine IFN-α1 (priming, 100 U mL−1 final) and/or with an infectious Paramixovirus [Newcastle Disease virus (NDV), Lasota vaccine strain] at 30 hemagglutinating units mL−1 for IFN induction. PBMC were incubated for 18 h at 37°C. Supernatants were tested for IFNs α and γ by sandwich ELISAs and the aforementioned MDBK bioassay; RNA was extracted from PBMC to evaluate IFN-α gene expression. In PBMC cultures of 3 other pigs of the same age and breed (IZSLER Experiment Station), IFN induction with NDV without priming under the above conditions was flanked in other wells by a stimulation with recombinant porcine interleukin-1 (IL-1)β (AbD Serotec Atlantic, cat. 19020) at 0.5, 2, and 8 ng/mL, respectively, in the presence/absence of recombinant porcine TNF-α at 0.4 ng/mL (Prospec, cat. Cyt 405). PBMC cultures were set up, and tests for IFNs α and γ were performed as described above.
Statistical analyses
The prevalence of IFN α-positive samples in the MDBK bioassay (serum), flow cytometry (PBMC), and in vivo gene expression (PBMC) was checked by Fischer's exact test. The significance threshold was set at P < 0.05. The expression level of a given target IFN-α gene in vitro was determined by the 2−ΔΔCt method (Bustin 2000; Pfaffl 2001). In particular, the levels of gene expression in vitro were computed as n-fold of the mean expression level in unstimulated PBMC cultures and checked by analysis of variance (Prism 2.01, GraphPad Software). The significance threshold was set at P < 0.05.
Results
Field trial: clinical findings
All pigs remained healthy during the whole period of observation. In particular, there was no detectable clinical difference under satisfactory hygiene and welfare conditions between the 2 groups of pigs under study.
Viral infections at weaning
All animals were PRV Ab-negative at both samplings (days −1 and +12 with respect to weaning). Nine out of 16 pigs tested positive for PRRSV-specific, maternally derived Ab on day −1; none of them was Ab-positive on day +12. Eight out of 10 pigs had detectable levels at weaning of maternally derived Ab to PCV2, which showed the same titers on day +12. As for Influenza viruses, the hemagglutination inhibition Ab titers were either identical between day −1 and +12 or nonsignificantly decreased (2-fold). Further, all sera of group 2 pigs tested negative in PCR assays for PRV, PRRSV and PCV2 on day +6. Results indicate that pigs were not infected by the usual viral agents of swine in the first 2 weeks after weaning.
IFN-α response at weaning
Despite the good health and farm hygiene conditions and the lack of detectable viral infections, a low-titered IFN-α response (1–27 IU/mL) was detected in many sera at day +6 after weaning in both groups (Table 2), the prevalence being significantly higher among group 2 piglets weaned at 22 days of age (P < 0.01). Most important, IFN-α was unambiguously identified by a neutralization assay on a pool of IFN-α-positive sera. By gel filtration chromatography on a Sephadex G-75 column, the antiviral activity of sera could be traced back to 3 components of apparent molecular mass 27/18/ < 14 kDa. NP-40 lysates of PBMC revealed the same molecular mass values as well as additional components of 58 and 41 kDa. Further, PBMC of 4 piglets in group 1 and 10 piglets in group 2 (P < 0.1) were also positive in a flow cytometry assay for intracellular porcine IFN-α at day +6 after weaning (Fig. 1). The prevalence of IFN-α-positive PBMC in flow cytometry showed no evident correlation with the titer of the corresponding serum IFN-α response (data not shown). On the whole, the results indicated that pigs mount an IFN-α response at weaning even under conditions of very low or absent infectious pressure.
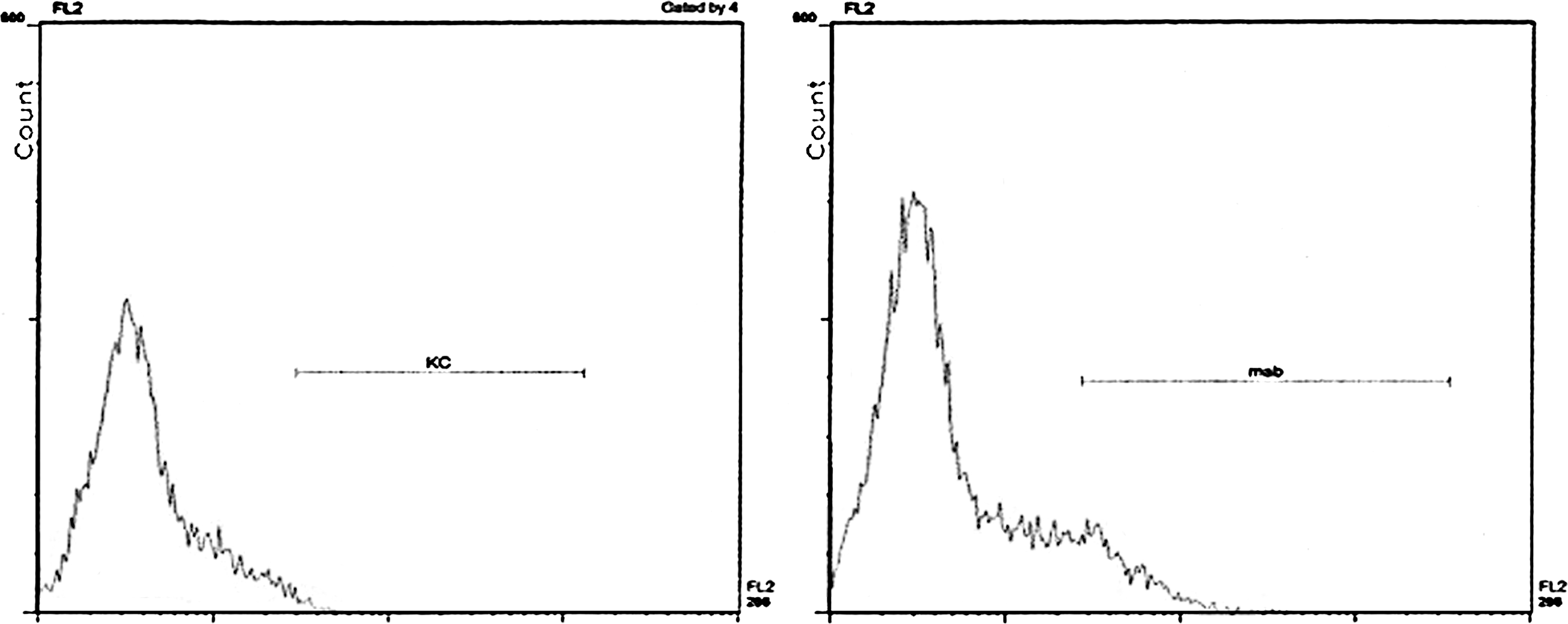
Flow cytometry assay for intracellular porcine IFN-α in swine PBMC at day +6 after weaning. Fixed and permeabilized swine PBMC were analyzed for intracellular IFN-α by flow cytometry. PBMC of 1 pig were stained with either mAb G16 + anti-mouse IgG1-Phycoerythrin (right) or buffer + anti-mouse IgG1-Phycoerythrin (control, left). IFN, interferon; PBMC, peripheral blood mononuclear cells; mAb, monoclonal Ab.
Results are expressed as number of IFN-α-positive sera out of the total (19 in group 1 and 20 in group 2) in a VSV cpe inhibition assay on MDBK cells, calibrated with Porcine recombinant IFN-α1.
Significantly different from the prevalence in group 2 on day +6 (P < 0.01).
IFN, interferon; VSV, vesicular stomatitis virus; MDBK, Madin-Darby bovine kidney.
In vivo IFN-α gene expression at weaning
The above results prompted us to investigate IFN-α gene expression at days −1, +6, and +12 with regard to weaning. On day −1, IFN A5/6 and A12 genes were shown to be expressed in 7 and 6 piglets, respectively; IFN A1, A2, A7/11 genes were expressed on the same day in fewer pigs. On the contrary, no IFN-α gene was expressed in PBMC on day +6, when plenty of pig sera were IFN-α-positive. Expression of the above genes was shown to resume on day +12: IFN A2 and A5/6 were detected in 6 out of 9 animals, whereas IFN A1, A7/11 and A12 were expressed in fewer pigs. IFN A3, A4, A8, and A9 genes were never expressed (Table 3). The time course of IFN A5/6 expression in Fig. 2 is representative of all the genes under study. For each IFN-α gene, there was no significant difference between the prevalence of constitutive expression among pigs on days −1 and +12; other differences (day −1 versus +6; day +6 versus +12) were obviously significant (P < 0.05). The complete lack of gene expression on day +6 (all real-time PCR tests negative at cycle 40) made any further quantitative analysis redundant and unnecessary. An almost complete lack of IFN-α gene expression was also observed in 2 age and breed-matched unweaned piglets of a small, low-stress group suckling their sow's milk (Table 3). Please notice that the specificity of the primers adopted for A3 gene amplification was less than optimal under our laboratory conditions; as a result, a caveat should be expressed about the expression of A3, which was anyhow much lower compared with other IFN-α genes.
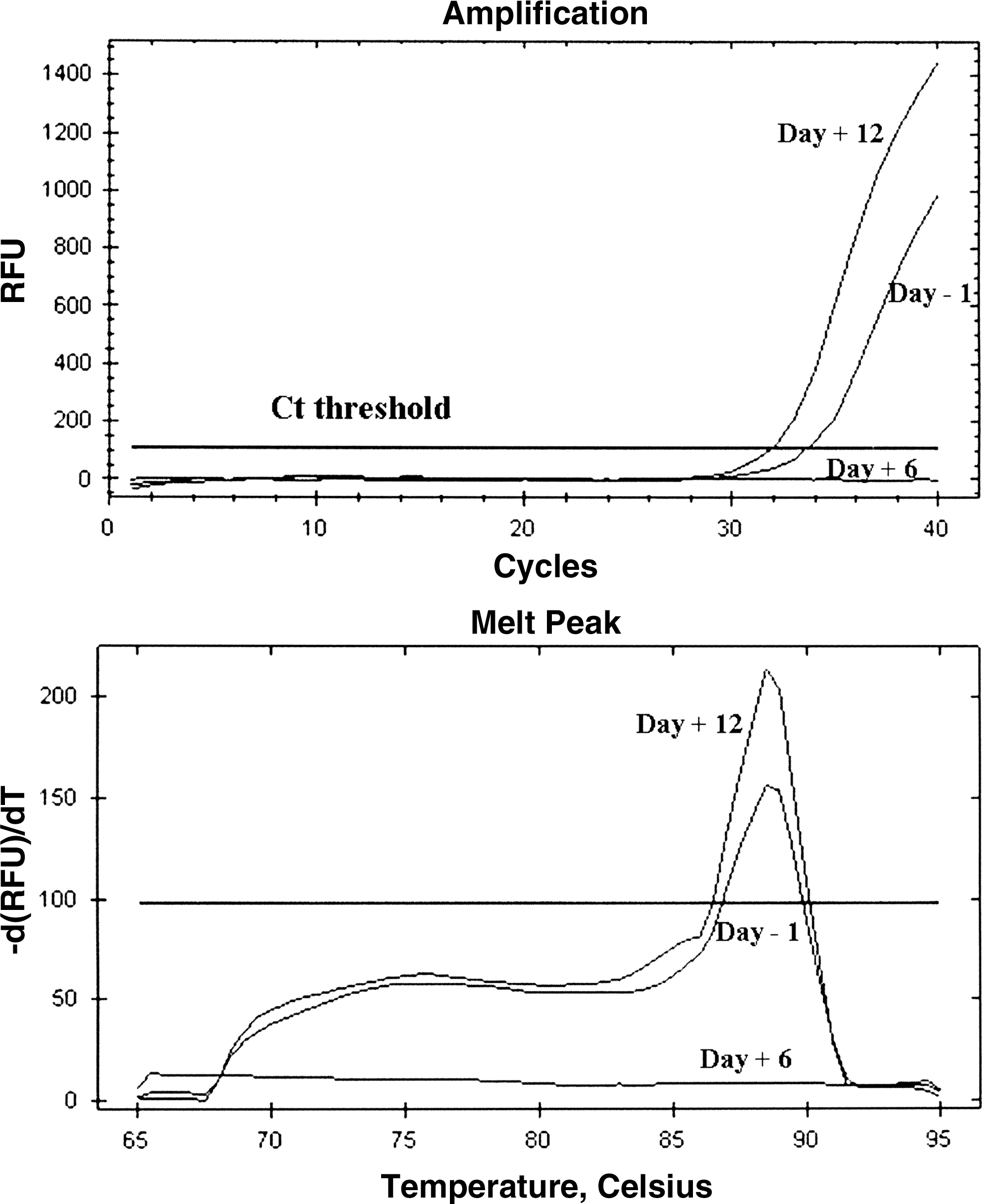
RT real-time PCR for porcine IFN-A5/6 genes at days −1, +6, and +12 with regard to weaning. RT real-time PCR for IFN-A5/6 genes was carried out on PBMC of the same pig of group 2 at days −1/ +6/ +12 with regard to weaning (upper graph). The specificity of the reactions was checked on the basis of the relevant melting curves (lower graph). PCR, polymerase chain reaction; RT, reverse transcriptase.
RT real-time PCR for IFN-α genes was carried out on uncultured PBMC of 9 pigs of group 2 at days −1, +6, and +12 with regard to weaning. All samples on day +6 tested negative. The same procedure was applied to PBMC of 2 age and breed-matched, unweaned control piglets in our Experiment Station.
+, fluorescence signal detected within real-time PCR cycle 40.
−, no signal detected within real-time PCR cycle 40.
PBMC, peripheral blood mononuclear cells; PCR, polymerase chain reaction; RT, reverse transcriptase.
IFN proteins and gene expression in vitro
The different involvement of IFN-α genes in constitutive expression and their modulation by the weaning stress raised 2 fundamental issues: does this pattern of response also occur in case of infectious (viral) stressors? Are inflammatory cytokines released at weaning implied in the modulation of type I IFN system? This is the reason that we investigated IFN genes and proteins in a relevant model of in vitro cultured swine PBMC from healthy, 70- to 80-day-old animals, far away from the weaning stress. In the experiments with a viral agent (NDV, Table 4), IFN-A1, A2, A4, A5/6, and A7/11 genes were shown to be involved in constitutive expression in uncultured PBMC (time 0 samples). Further IFN-α genes were then expressed in culture. Interestingly, expression was often shown in unstimulated PBMC cultures as well. As a result, only the expression of IFN A9 gene was shown to be significantly higher (P < 0.01) after exposure to NDV (with/without priming) (Fig. 3); no significant difference was shown instead for all the other genes under study between virus-stimulated and control cultures. As for the release of IFN-α, this was absolutely dependent on NDV stimulation with or without priming. Pig 1 was a low responder as opposed to the other ones (pigs 2,3,4), which also released IFN-γ after NDV stimulation. On the whole, the correlation between the ELISA and the cell culture assays for IFN-α was poor for all the samples under study.
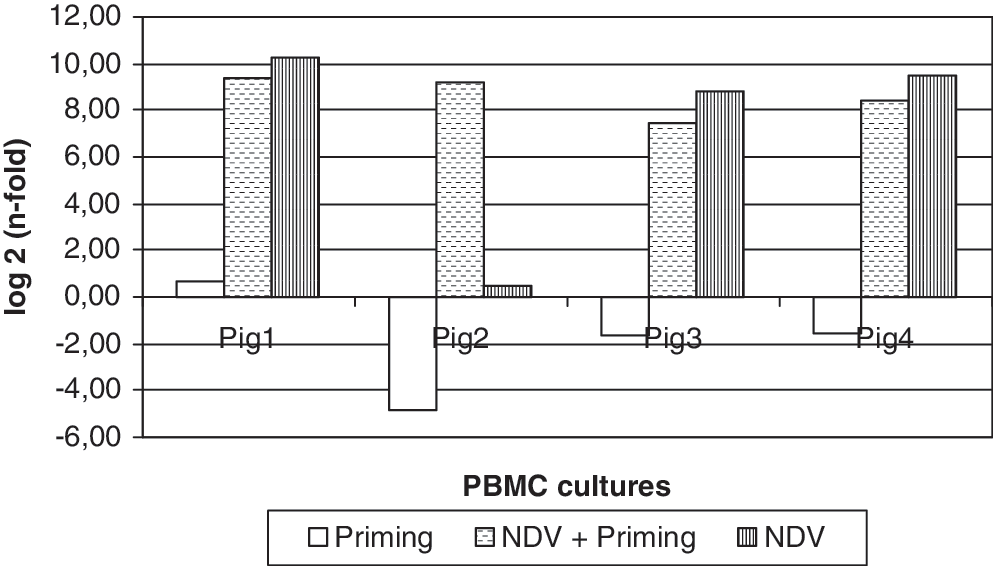
Expression of porcine IFN-A9 gene in vitro. PBMC of 4 pigs were either kept as unstimulated controls or treated for 2 h with recombinant porcine IFN-α1 (priming, 100 U mL−1 final) and/or with an infectious Paramixovirus (Newcastle Disease virus, NDV, Lasota vaccine strain) at 30 hemagglutinating units mL−1 for IFN induction. PBMC were incubated for 18 h at 37°C. RNA was extracted from PBMC, and IFN-A9 gene expression was measured by RT real-time PCR. Expression of IFN-A9 gene is computed as n-fold of the expression level in unstimulated control cultures. Expression is significantly different (P < 0.01) after NDV stimulation with and without priming.
PBMC of 4 pigs were either kept as unstimulated controls or treated for 2 h with recombinant porcine IFN-α1 (priming, 100 U mL−1 final) and/or with an infectious Paramixovirus [Newcastle Disease virus (NDV), Lasota vaccine strain] at 30 hemagglutinating units mL−1 for IFN induction. PBMC were incubated for 18 h at 37°C. Supernatants were tested for IFNs α and γ by sandwich ELISAs and a MDBK bioassay; RNA was extracted from PBMC to evaluate IFN-α gene expression by RT real-time PCR.
IFN proteins: results are reported in terms of IU/mL and pg/mL for IFN-α and IFN-γ, respectively.
IFN-α genes: expression within 40 real-time PCR amplification cycles is reported as + if confirmed by the relevant melting curve.
As for the inflammatory cytokine model in vitro, IL-1β with or without TNF-α gave rise to the expression of IFN-A9 gene and low-titered protein release in 1 pig, whereas a second one showed these latter features at time 0; the subsequent stimulation with NDV blocked in the latter pig the expression of all the IFN-α genes under study but not the secretion of IFN-α protein (Table 5). The aforementioned caveat about A3 gene amplification is also valid for our in vitro experiments.
PBMC cultures of 3 pigs were either kept as unstimulated controls or treated for 18 h at 37°C with either NDV at 30 hemagglutinating units mL−1 or recombinant porcine IL-1β at 0.5, 2, and 8 ng/mL, respectively, in the presence/absence of recombinant porcine TNF-α at 0.4 ng/mL. Supernatants were tested for IFNs α and γ by sandwich ELISAs and a MDBK bioassay; RNA was extracted from PBMC to evaluate IFN-α gene expression by RT real-time PCR. IFN proteins: results are reported in terms of IU/mL and pg/mL for IFN-α and IFN-γ, respectively. After neutral red staining and color extraction, OD values below the 50% end point in MDBK cells but significantly different from those of VSV-treated control cells correspond to IFN-α titres <1 IU/mL. IFN-α genes: expression within 40 real-time PCR amplification cycles is reported as + if confirmed by the relevant melting curve.
ND, not done.
TNF, tumor necrosis factor; IL, interleukin.
Discussion
The results obtained in this study complement those of previous investigations into the biological activities of IFN-α in the SPF pig model (Amadori and others 2009), and concur to a global view of type I IFN as homeostatic control system of both infectious and noninfectious stressors. Our experimental setting included conventional healthy pigs exposed to low environmental microbial pressure, to detect and characterize their constitutive type I IFN response. The subsequent stress of early versus late weaning (22 and 28 days of age, respectively) was the scenario intended for evaluating an in vivo homeostatic control action of IFN-α; in vitro constitutive and virus-induced IFN-α responses were determined as well. Finally, IL-1β was investigated in vitro due to its role in IFN induction (Soos and Szente 2003) and in weaning stress (McCracken and others 1995; Pié and others 2004). TNF-α was checked with IL-1β at a concentration often found in pig sera in the first weeks after weaning.
Our results indicate that components of the type I IFN family are constitutively expressed in pigs and actively involved in the homeostatic control of weaning stress. The latter consists of poor absorption of nutrients and electrolytes, related morphological alterations of the gut (shortening of villi, elongation of crypts), worsening of local immunity parameters, abrupt breakage of sow/piglet interactions, and environmental changes (transfer to weaning cages), which represent as many physical, emotional, nutritional, and immunological stressors with additive or synergic, potentially dangerous effects in terms of animal health and welfare (Wattrang and others 1998; McCracken and others 1995; Hameister and others 2010). Interestingly, the main metabolic responses at weaning (decreased plasma glucose, increased plasma glucagon, fibrinogen, and IL-1) were shown to be diet-independent (McCracken and others 1995).
The IFN-α response under conventional field conditions in this study could not be accounted for by the clinical and environmental conditions of pigs, nor was there serological or molecular evidence of the most common viral infections of swine; the lack of viral infections was confirmed by the unabated growth in both groups of pigs (data not shown). Also, the extent of the IFN-α response was shown to be different in subjects weaned at 3 versus 4 weeks of age. In fact, the IFN-α response on day +6 occurred at higher frequency in group 2 pigs (early weaning), in agreement with previous reports about the age-dependence of the weaning stress (Hameister and others 2010). This actually implies in younger pigs a larger and/or more prolonged homeostatic control action and a higher energy expense to the detriment of the weight gain; this was actually confirmed by the greater mean weight of group 1 pigs at 48 days of age (+1.5 kg on average, P < 0.01) (data not shown).
As for intracellular IFN-α in PBMC, positive samples showed on average a prevalence of 2% IFN α-positive cells in flow cytometry assays, which must be compared with a much lower prevalence in 2-month old animals, far away from weaning stress (Amadori and others 2010). The apparent molecular mass values of serum IFN-α point at the presence of different subtypes and/or isophorms of IFN-α, as also described in a previous study based on gel filtration chromatography (Piasecki 1988). Further, the additional, intracellular 58 and 41 kDa components in PBMC are consistent with the sizes of a dimer and glycosilated monomer, respectively, in agreement with previous data about constitutive expression of IFN-α in human PBMC (Greenway and others 1995). Further, constitutive expression of both dimers and nonglycosilated IFN-α monomers had been revealed by radio-immunoprecipitation in PBMC of 65-day old pigs, far away from weaning stress (Amadori and others 2010). The low molecular mass component in serum and NP-40 lysates of PBMC (<14 kDa) was evidenced in 3 distinct gel filtration chromatography assays and could not be unambiguously interpreted. It could be a biologically active, truncated fragment of an unknown IFN-α subtype or another low molecular weight component of swine type I IFNs, such as the recently described 139 aa ovine IFN-δ (the shortest natural type I IFN presently known) (Cochet and others 2009). It is clear though that much additional work will be required before a complete understanding of the phenomena occurs.
The serum IFN-α response could be offset against the pattern of constitutive gene expression around weaning. IFN A5/6 and A12 genes showed the highest frequency of expression, whereas lower frequencies were observed for IFN A1, A2, and A7/11 genes. The same genes were also expressed at similar frequencies and intensity at day 12 after weaning, whereas no IFN-α gene expression was observed on day +6, in the presence of a serum IFN-α response in most pigs under study. Such a transient inhibition of gene transcription is probably a feed-back mechanism for limiting the serum IFN-α response at the time of its own peak. On the basis of previous data obtained in virus infection models, the effector mechanism is likely to involve a transient block to IFN regulatory factor 3 function (Baigent and others 2002; Bossert and others 2003). Likewise, the increased IFN-α protein expression on day +6 suggests that downstream signaling components such as JAK/STAT may also be altered. Interestingly, IFN-α gene expression was almost lacking in 2 unweaned, low-stress control piglets (Table 3), which is further circumstantial evidence of a correlation between environmental stressors and activation of the type I IFN system.
By a careful comparison of in vivo IFN-α gene expression data, a certain discrepancy can be observed between 3-week and 70- to 80-day-old animals (compare data in Table 3 with time 0 data in Tables 4 and 5). In particular, one of the older pigs (pig 1 in Table 4) showed at time 0 the expression of IFN A4, whereas none of them expressed A12. This indicates that constitutive expression of type I IFNs may be affected by both age and metabolic phase of animals. Also, only IFN A9 was shown to be absolutely virus induction-dependent in vitro; as a result, A9 gene expression at time 0 in pig 2 (Table 5) was probably due to a recent virus infection in vivo. The correlation between the MDBK and the ELISA assays for IFN-α was not high, which points at a certain bias related to the IFN-α subtype mixtures in serum samples (see Tables 4 and 5).
In general, beyond the IFN-α genes constitutively expressed (time 0 in Tables 4 and 5), further genes were expressed after in vitro NDV stimulation, with or without priming (Tables 4 and 5). However, gene expression was often shown also in control, unstimulated PBMC cultures, and the relevant levels of expression (ΔCt values) were often similar to those of NDV-stimulated cultures. The same concept holds true of PBMC cultures submitted only to priming. However, only NDV-stimulated PBMC cultures gave rise to IFN-α protein release in their supernatants. Therefore, IFN-α release by swine PBMC can be largely explained in terms of posttranscriptional control activities. Our findings are reminiscent of both constitutive and virus-induced expression of type I IFN in humans: In fact, IFN A5 and beta genes are constitutively expressed in human PBMC, whereas multiple subspecies are expressed in the same cells after viral induction; constitutive expression is associated to minute extracellular IFN concentrations, which are outside the usual detection range of routine assays (Lallemand and others 1996).
Also, IFN-γ release was shown to be absolutely NDV-dependent in our in vitro model, which points at a role of pig NK cells (CD3−, CD8α+, CD5−) (Saalmuller and others 1994; Krog and others 2003); instead, a possible role of Ag-specific memory T cells can be substantially ruled out, as NDV infects only avian species.
On the whole, the accumulated data are consistent with a model of synergic or complementary stimulation of type I IFNs in swine PBMC, whereby in vivo signals can induce and sustain a constitutive response; the latter could be possibly affected by lipopolysaccharide (LPS) levels in plasma, ranging in healthy outbred pigs between 50 and 200 pg/mL under different conditions of environmental infectious pressure (Candotti and Amadori, unpublished results); LPS can, in fact, induce type I IFN gene transcription by TLR-4 signaling (Pietras and others 2006), and a further regulation of constitutive expression can be accomplished by nitric oxide (negative regulator) (Guillemard and others 1999), plasminogen activation inhibitor type 2 (positive regulator) (Antalis and others 1998), and Th2 cytokines such as IL-4 and IL-10, which exert a strong posttranscriptional control on type I IFN genes (Varano and others 2000); if this were the case in our pig model, the shift to a predominant pro-inflammatory cytokine milieau after weaning would circumvent the possible negative regulation by Th2 cytokines and induce the secretion of minute amounts of type I IFNs. This is actually seen in vitro in murine macrophage cultures after exposure to inflammatory stimuli such as LPS, IFN-γ, or IL-12 (Gessani and others 1989). In the presence of a constitutive IFN response, virus-specific signals (probably mediated by Toll-like-Receptors and/or RIG-like helicases) (Saito and Gale 2007) would induce the transcription of the A9 gene (Tables 4 and 5); this could be the signal for efficient translation of IFN-α genes, maybe obtained by stabilizing the corresponding mRNAs. Further, if the A9 gene is already expressed as a result of in vivo exposure to a viral agent (see pig 2 in Table 5), then an in vitro virus stimulation could shut down the transcription of all IFN-α genes and cause the release of preformed IFN-α protein from PBMC. This would explain the complete discrepancy between gene and protein expression in vitro observed in the aforementioned pig (Table 5) and could partly account, in general, for virus-induced IFN-α release.
The results obtained in the experiments of in vitro IFN induction by inflammatory cytokines are relevant to the in vivo scenario after weaning, in which IL-1β plays a major role in the response to weaning stress (McCracken and others 1995; Pié and others 2004). In previous studies in our lab, porcine IL-1β in a wide range of concentrations could induce low-titered IFN-α in whole blood cultures (Amadori, unpublished results). Our results in this study confirm that IL-1β may induce the expression of IFN A9 gene in vitro and the release of minute amounts of IFN-α proteins, which is somehow reminiscent of the in vivo conditions at weaning; IL-1β did not instead induce IFN-γ (Table 5). Thus, on the basis of our in vitro findings, IL-1β could contribute to the IFN-α response at weaning, also detectable as low-titered cytokine in serum. In particular, IFN-α production in the intestinal tract after weaning could be accounted for by a generalized inflammosome reaction in enterocytes, leading to the release of biologically active IL-1β after caspase 1 activation (Martinon and others 2009); interestingly, marked upregulation of IL-1β mRNA occurs between day 0 and 2 after weaning in most parts of the intestine in pigs (Pié and others 2004). Owing to the above, a note of caution should be expressed about the actual origin of IFN-α-producing cells after weaning. In particular, we cannot rule out a possible contribution of intestinal, intraepithelial and/or lamina propria leukocytes, and/or of mesenteric lymph nodes, which were shown to sustain a systemic IFN-α response to a viral infection, also detectable in serum (Riffault and others 2001).
The above functions of type I IFNs in pigs outline a resilient system aimed at efficiently coping with microbial and nonmicrobial stressors. In this regard, within type I IFN system, synergism and cooperation between IFN-β and IFN-α genes are probably of paramount importance and a priority of future studies. Such a complex homeostatic system is likely to involve both transcriptional and posttranscriptional effector mechanisms, including the conversion of prestored IFN dimers (Amadori and others 2010) into mature monomers and their subsequent release. Further, the constitutive expression of both type I and gamma IFNs in PBMC (Amadori and others 2010) suggests that pigs need to exert a fine tuning and control of the inflammatory response under conditions of frequent, heavy exposure to air-driven LPS (Zhiping and others 1996); also, the emergence of lean type, rapid growth pig breeds with very high plasma concentrations of reactive oxygen metabolites (Brambilla and others 2002) further compounds this issue. Therefore, future comparative studies on rural versus lean type pig breeds could provide evidence of a rapid adaptation of the IFN system to the outcomes of genetic selection in farm animals, which obviously proceeds at a much faster pace than phylogenetic evolution.
Footnotes
Acknowledgments
The authors wish to thank Drs. Paolo Candotti and Nicola Martinelli (Istituto Zooprofilattico Sperimentale, Brescia, Italy) for the assistance in blood samplings. The skilful technical assistance of Mr. Antonio Cristiano, Mr. Michele Testa, and Mrs. Cinzia Mantovani are gratefully acknowledged. This study was partly funded by the Italian Ministry of Health, grant PRC2007003, and a EU EPIZONE grant.
Author Disclosure Statement
No competing financial interests exist.