
Abstract
Avian influenza virus is endemic in many regions around the world and remains a pandemic threat, a scenario tied closely to outbreaks of the virus in poultry. The innate immune system, in particular the nucleic acid-sensing toll-like receptors (TLRs) -3, -7, -8, and -9, play a major role in coordinating antiviral immune responses. In this study we have investigated the use of TLR ligands as antivirals against influenza A in chickens. The TLR7 ligand poly-C inhibited low-path influenza A growth in the chicken macrophage cell line HD-11 more effectively than poly(I:C), which acts via TLR3. The TLR7 ligand 7-allyl-8-oxoguanosine (loxoribine) inhibited influenza A replication in vitro and in ovo in a dose-dependent manner. Treatment of primary chicken splenocytes with loxoribine resulted in the induction of interferons-α, -β, and -λ, and interferon-stimulated genes PKR and Mx. These results demonstrate that nucleic acid-sensing TLR ligands show considerable potential as antivirals in chickens and could be incorporated into antiviral strategies.
Introduction
V
In mammals, the first line of antiviral defence is coordinated by pattern recognition receptors (PRRs) that recognize pathogen-associated molecular patterns unique to particular pathogens (Heil and others 2003). A subset of the family of PRRs known as Toll-like receptors (TLRs) is largely responsible for recognizing viral nucleic acids and activation of antiviral cellular responses. TLRs-3, -7, -8, and -9 are considered the antiviral TLRs, activation of which leads to activation of transcription factors nuclear factor kappa B (NF-κB), interferon regulatory factor (IRF)-3 and IRF-7, resulting in production of the antiviral type I interferon (IFN) cytokines. Type I IFN (IFN-α and IFN-β in humans) bind to the IFN-α receptor, resulting in downstream effects including the shutdown of RNA and protein synthesis in virally infected cells, apoptosis of virally infected cells, and the induction of IFN-stimulated genes (ISGs) that coordinate downstream antiviral measures (Wathelet and others 1998).
7-allyl-8-oxoguanosine (loxoribine) is an analog of the DNA nucleoside guanosine. Loxoribine is immunostimulatory through TLR7-dependent activation of NF-κB in mouse and human cells (Heil and others 2003; Lee and others 2003), resulting in production of type I and type III IFNs, in addition to interleukin (IL)-6 and IL-12 (Pope and others 1995; Lee and others 2003). Here we investigate the use of TLR7 ligands as antivirals in chicken models of influenza infection. Our results demonstrate the ability of loxoribine to inhibit influenza A in both in vitro and in ovo models of chicken influenza A infection, and demonstrate the potential for TLR7 ligands to be incorporated into antiviral strategies.
Materials and Methods
Materials
Poly(I:C), poly-C and loxoribine were purchased from Invivogen (San Diego, CA).]
Cell lines
The continuous chicken macrophage cell line HD-11 was maintained in RPMI media supplemented with 10% (v/v) fetal calf serum (FCS), 10 mM HEPES, 2 mM L-glutamine, 100 U/mL penicillin, and 100 μg/mL streptomycin. Madin–Darby Canine Kidney (MDCK) cells (ATCC CCL-34) were maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with 10% (v/v) heat-inactivated FCS, 2 mM
Virus
Influenza A/Puerto Rico/8/34 (H1N1) (PR8) was passaged in 10-day embryonated specific pathogen-free chicken eggs. Allantoic fluid was harvested, aliquoted, and stored at −80°C for inoculations. Virus titer was measured by plaque assays, whereby serial 10-fold dilutions of viruses were incubated on MDCK cells for 30 min and then cells were overlayed with media containing 1% agar. Two days after infection, plaques were observed by staining with crystal violet.
Harvesting of embryo allantoic fluid
Eggs were chilled for 1 h at 4°C and swabbed with 70% ethanol to disinfect the shells. Eggshells were removed from above the air space, and a sterile Pasteur pipette used to remove ∼1 mL of allantoic fluid from each egg.
Hemagglutination assays
Hemagglutination (HA) assays were carried out in U-bottom 96-well plates. Serial 2-fold dilutions of virus samples were mixed with an equal volume of a 0.5% suspension (v/v) of chicken erythrocytes and incubated at 4°C for 30 min. Wells containing an adherent, homogeneous layer of erythrocytes were scored as positive.
Treatment of eggs with loxoribine
Loxoribine (at indicated doses) was delivered to 10-day-old embryonated specific pathogen-free eggs by injection into the allantoic fluid through the eggshell membrane. Eggs were sealed with heated parafilm and incubated at 37°C. Sixteen hours post-treatment, eggs were inoculated with virus and incubated at 37°C for indicated time periods.
Preparation of splenocytes
Primary chicken splenocytes were prepared as described previously (Jenkins and others 2009). Briefly, spleens were removed from 5-week-old specific pathogen-free chickens and single-cell suspensions prepared and strained through a 70 μm nylon cell strainer onto prewarmed Lymphoprep solution. Cell suspensions were centrifuged at 1100 g max for 20 min at room temperature. The splenic mononuclear cell layer was collected, washed in phosphate-buffered saline (PBS), and resuspended in 10 mL of DMEM with 5% (v/v) FCS.
Stimulation of chicken splenocytes with loxoribine
Splenocytes were seeded in 12-well plates at 1×106 cells/well in DMEM, 10% FCS, with 100 U/mL penicillin and 100 μg/ML streptomycin (splenocyte media). Splenocytes were stimulated with indicated concentrations of loxoribine in splenocyte media.
RNA isolation, reverse transcription, and quantitative real-time polymerase chain reaction
RNA was harvested using Tri-reagent (Sigma) according to manufacturer's instructions. One microgram of extracted RNA was treated with DNase (Sigma) according to manufacturer's instructions and reverse-transcribed to complementary DNA (cDNA) using a Reverse Transcription kit (Promega). Quantitative real-time polymerase chain reaction (QRT-PCR) experiments were conducted on an ABI Prism 7700 Sequence Detection System (Applied Biosystems). The comparative threshold cycle (C t) method was used to derive fold change gene expression. All results were normalized to chicken glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Primers and probes were designed using Primer Express software (Applied Biosystems). Where possible, the probe sets were designed across intron:exon boundaries. PCR cycling was performed as follows: 95°C for 1 min, followed by 40 cycles of 95°C for 15 s, 60°C for 30 s and 72°C for 30 s.
QRT-PCR probes
Probes for the detection of chicken IFN-α, IFN-β, IFN-λ, and GAPDH have been described previously (Karpala and others 2008a, 2008b). Sequences for detecting chicken PKR and chicken Mx are given in Table 1.
Tissue culture infectious dosage50 analysis
Ten-fold dilutions of supernatants containing virus were added to a 96-well tissue culture plate. MDCK cells were added to all wells and the plates were incubated for 1 h at 37°C, 5% CO2. The medium was discarded and replaced with virus growth media (Eagle's minimum essential media, 0.3% bovine serum albumin, 1 mM Hepes, 5 mM glutamine, 100 U/mL penicillin, 100 μg/mL streptomycin, and 5 μg/mL trypsin). The plates were incubated for 4 days and scored for cytopathic effect. The infectious titer was calculated by the method of Hawkes (1979).
Statistics
Statistics were performed using a 2-tailed Student t-test.
Results
Influenza A/Puerto Rico/8/34 (PR8) virus growth was measured in the immortalized chicken macrophage cell line, HD-11, in the presence and absence of the TLR3 ligand [poly(I:C) and TLR7 ligands (poly-C) and loxoribine]. Cells were pretreated with ligands for 6 h to induce an antiviral response, and then infected with PR8. When infected at an multiplicity of infection (m.o.i.) of 10, cells pretreated with poly-C showed a large decrease in tissue culture infectious dosage50 (TCID50) titers at 72 h compared to untreated control cells (Fig. 1A). Cells treated with poly(I:C) supported virus growth at a level comparable to untreated cells. The effect of loxoribine on PR8 virus growth in HD-11 cells (virus m.o.i. 10) was tested over a range of loxoribine concentrations (Fig. 1B), with 0.1 mM of loxoribine resulting in significant reduction in virus growth compared to untreated cells.
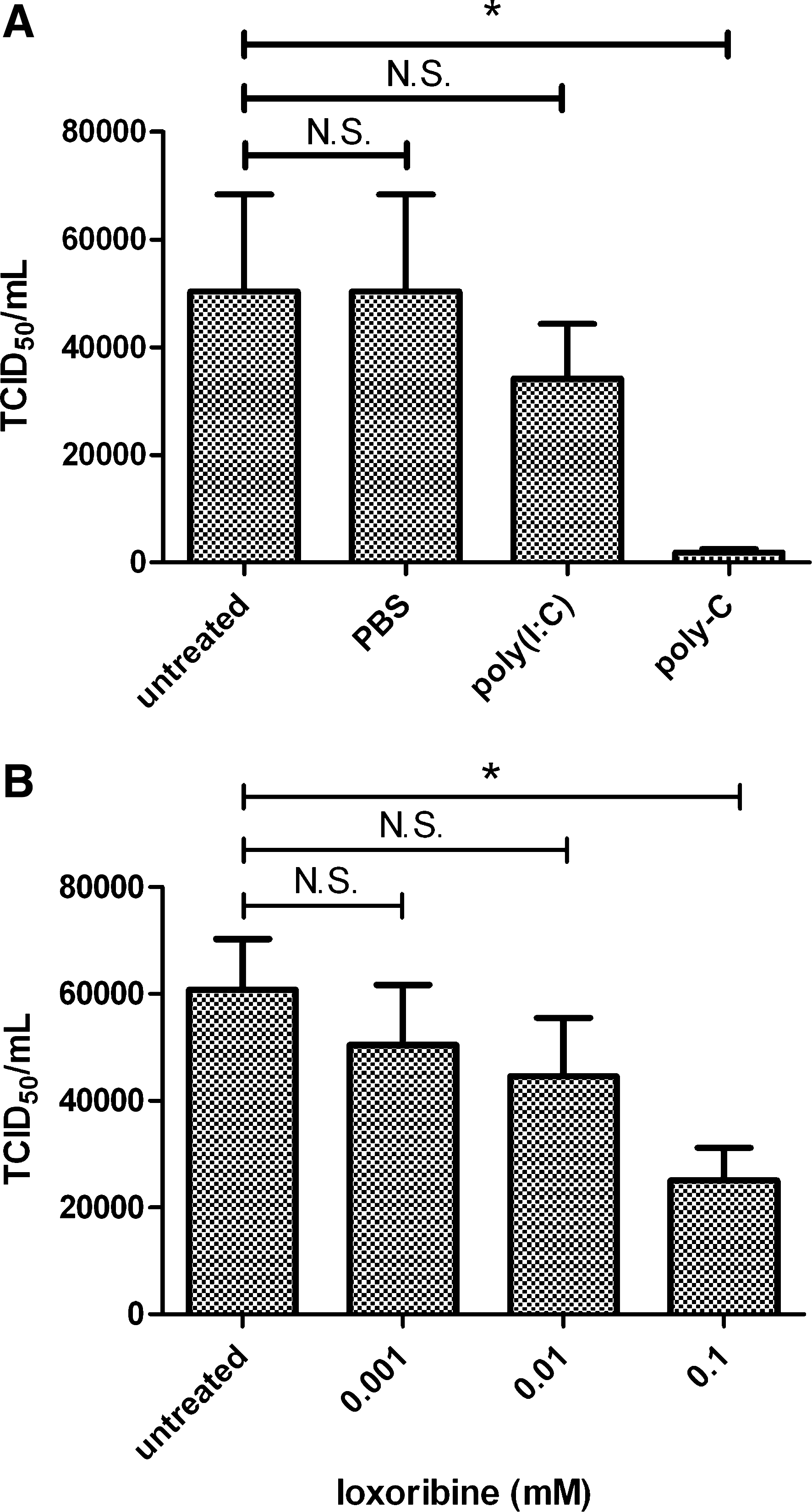
TLR 7 ligand inhibits influenza A/PR8 replication in chicken cells.
We tested the antiviral properties of loxoribine in ovo. Ten-day-old chicken embryos were injected with loxoribine 24 h before virus infection to allow time for a protective antiviral response to occur. The concentrations of loxoribine used were guided by previous administration of loxoribine to 18-day old chicken embryos (Jenkins and others 2009). Pre-treatment of 10-day old chicken embryos with loxoribine, followed by infection with 500 plaque forming units (pfu) PR8 virus for 24 h, resulted in a concentration-dependent decrease in PR8 virus levels in virally-infected embryos (Fig. 2A). The largest inhibition of virus growth was observed for eggs treated with 50 μg loxoribine. To determine whether loxoribine could inhibit PR8 virus during periods of peak virus infection, the growth of PR8 virus in chicken embryos was measured in a time- and dose-dependent manner. Chicken embryos were infected with 50 to 50,000 pfu of PR8 virus, with virus levels measured by HA assay 24, 48, and 72 h postinfection. For all dosage groups, PR8 virus was detected at 24 h, with large increases in virus levels detected at 48 h (Fig. 2B). For eggs infected with 50 pfu of virus, maximum virus titers were observed after 72 h. For other treatment groups (eggs infected with 500, 5,000 or 50,000 pfu), maximum virus titers were observed at 48 h.

TLR 7 ligand loxoribine inhibits influenza A/PR8 replication in chicken embryos.
The effect of loxoribine on the growth of PR8 virus in 10-day-old chicken embryos was tested 24 h and 72 h postinfection with 50 pfu PR8 virus/egg. Consistent with results observed in Fig. 2A, loxoribine (10 μg/egg) inhibited PR8 growth after 24 h, with virus levels measured by HA assay (Fig. 2C) and TCID50 assay (Fig. 2D). Pretreatment of chicken embryos with loxoribine also resulted in significant inhibition PR8 virus after 72 (Fig. 2C, D). As quantitated by TCID50 assays, loxoribine caused an 85% and 95% reduction in virus titer at 24 and 72 h, respectively, compared to control eggs treated with PBS. While there was a difference in HA titers between eggs infected with 500 pfu/virus for 72 h (Fig. 2B, C), HA titers from eggs treated with loxoribine were consistently lower than HA titers from eggs treated with PBS, within the same experiment. We investigated the downstream genes induced by loxoribine that may mediate antiviral properties in chicken embryos. We have previously demonstrated that the treatment of primary chicken splenocytes with loxoribine results in cytokine production, and that cytokine profiles of splenocytes are consistent with cytokine profiles of chicken embryos administered loxoribine in ovo (Jenkins and others 2009). Primary chicken splenocytes were stimulated with 2 different concentrations of loxoribine (1 mM and 10 mM) for 6 and 24 h. After these times cellular RNA was purified and used for QRT-PCR analysis of splenocyte gene expression levels (Fig. 3). After 6 h, loxoribine stimulated increases in IFN-α and IFN-β mRNA expression levels at low doses, while levels of Mx and PKR were elevated at 6 h by high doses of loxoribine. At 24 h, IFN-α and IFN-β were elevated in chicken splenocytes treated with the higher dose of loxoribine, whereas IFN-λ was elevated in both low- (∼2-fold) and high-dose groups. Of the ISGs, levels of PKR were elevated at 24 h by both low and high doses of loxoribine.
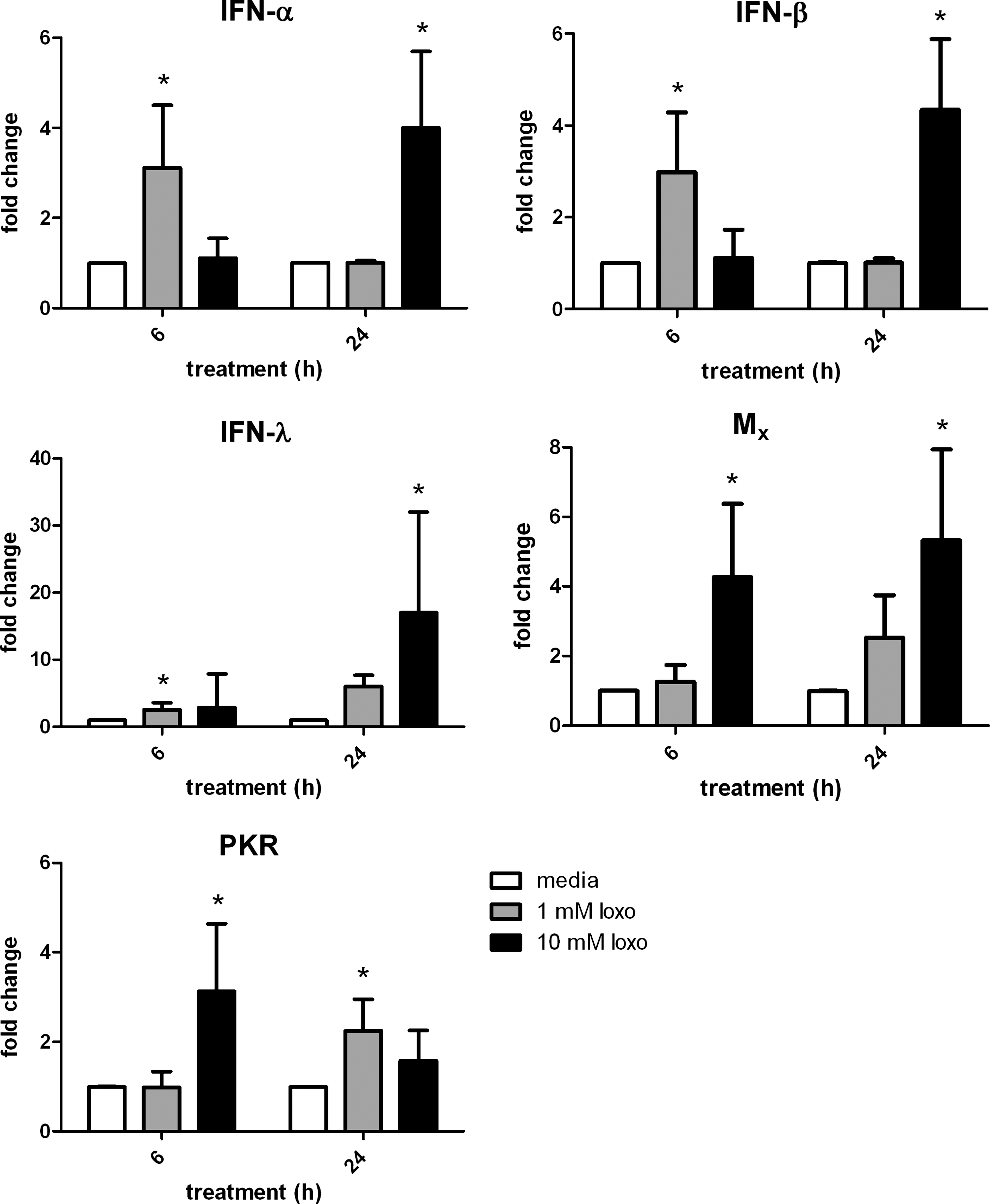
Loxoribine induces antiviral genes in primary chicken splenocytes. Primary chicken splenocytes were treated with media (white bars), 1 mM loxoribine (gray bars), or 10 mM loxoribine (black bars) for 6 h or 24 h. Quantitative real-time polymerase chain reaction was used to measure changes in gene expression. Results represent means from splenocytes isolated from 3 chickens. Error bars represent standard error. *p<0.05 compared to untreated cells.
Discussion
This study is the first to demonstrate the antiviral properties of the TLR7 ligand loxoribine in chickens, inhibiting growth of low-pathogenic influenza A PR8 virus in 10-day-old chicken embryos and inducing a range of antiviral genes in chicken splenocytes. We have previously shown that loxoribine stimulates production of IL-6 and IFN-α in chicken splenocytes (Jenkins and others 2009). We have also shown that in ovo administration of loxoribine to E18 chicken embryos resulted in elevated levels of IL-6 that can be detected in chicken spleens 7 days posthatch (Jenkins and others 2009). Collectively, these results demonstrate the ability of loxoribine to induce a significant antiviral response in chickens.
The ability of loxoribine to induce antiviral cytokines and to block low-path influenza suggests that it could be incorporated into and compliment existing antiviral avian influenza strategies. Vaccination of poultry to prevent avian influenza continues to be performed routinely in South-East Asia, where outbreaks of avian influenza in poultry regularly occur. Loxoribine has been used as an adjuvant to enhance antitumor vaccines in mice (Pope and others 1994), with adjuvant activities attributed to activation of natural killer cells and adjuvant effects on B cells. The antiviral properties of loxoribine, coupled to its ability to induce known immuno-modulators such as IFN-λ in chicken embryos, suggest a role for loxoribine as an antiviral adjuvant in poultry.
An interesting finding from this study is that loxoribine induces ISGs Mx and PKR. Upon virus infection, PKR dimerizes and becomes active, phosphorylating and activating the translation initiation factor eIF2α which in turn halts protein synthesis and virus production [reviewed by Wolff and Ludwig (2009)]. The influenza virus NS1 gene segment is known to antagonize the RNA-mediated dimerization of PKR in mammals, therefore promoting virus replication, and can also antagonize antiviral pathways by blocking retinoic acid-inducible gene 1 protein (RIG-I) signaling. The apparent lack of RIG-I in the chicken genome (Barber and others 2010) increases the likelihood that receptors such as PKR play critical roles in chicken antiviral responses.
In primary chicken splenocytes, Mx and PKR were elevated by the higher dose of loxoribine at 6 h post-stimulation, suggesting that the type I IFN pathway was induced swiftly. At the lower dose of loxoribine, type I IFN was elevated at 6 h, which may have driven ISG production observed at 24 h. Both type I IFN and ISGs were up-regulated at 24 h, suggesting a bimodal induction of IFN responses.
An intriguing finding from this study is that while activation of TLR3 signaling pathways results in the production of antiviral effector molecules (Karpala and others 2008a), HD-11 cells treated with a TLR3 ligand [poly(I:C)] did not support virus replication less than control HD-11 cells (Fig. 1A). This result would suggest that influenza A is capable of blocking antiviral signaling pathways downstream of TLR3 signaling, a hypothesis that we are currently investigating further.
An attractive feature of loxoribine as an antiviral therapy is that while it stimulates TLR7, it is stable in vivo and not subject to degradation like naturally occurring nucleic acids. Antiviral and antitumor activities of loxoribine have been demonstrated in mouse models of diseases, and loxoribine has been trialed as an antitumor treatment in human patients with advanced cancers (Agarwala and others 2000). Interestingly, immunostimulatory short interfering RNAs have also shown promise recently in as anticancer and antiviral therapeutics. The conjugation of siRNAs to CpG oligonucleotides that activate TLR9 result in potent immune responses that facilitate antitumor treatment in mice (Kortylewski and others 2009). The expanding use of polymers, such as those synthesized using Reversible addition-fragmentation chain transfer technology, as vehicles for in vivo siRNA delivery (Zhang and others 2007) creates opportunities for immunostimulators such as loxoribine to be complexed to in vivo delivery vehicles to enhance antiviral and antitumor therapies. The results of this present study and others would suggest that loxoribine possesses antiviral properties worth incorporating into future anitiviral strategies.
Footnotes
Acknowledgments
We thank Susanne Wilson and Terri O'Neil for supplying eggs.
Author Disclosure Statement
The authors have no conflicts of interest to disclose.