
Abstract
Autoimmune uveitis is a complex group of sight-threatening diseases that arise without a known infectious trigger. The disorder is often associated with immunological responses to retinal proteins. Experimental models of autoimmune uveitis targeting retinal proteins have led to a better understanding of the basic immunological mechanisms involved in the pathogenesis of uveitis and have provided a template for the development of novel therapies. The disease in humans is believed to be T cell-dependent, as clinical uveitis is ameliorated by T cell-targeting therapies. The roles of T helper 1 (Th1) and Th17 cells have been major topics of interest in the past decade. Studies in uveitis patients and experiments in animal models have revealed that Th1 and Th17 cells can both be pathogenic effectors, although, paradoxically, some cytokines produced by these subsets can also be protective, depending on when and where they are produced. The major proinflammatory as well as regulatory cytokines in uveitis, the therapeutic approaches, and benefits of targeting these cytokines will be discussed in this review.
Introduction
The model of experimental autoimmune uveitis/uveoretinitis (EAU) in rodents is used as an animal model for human uveitis. The classical model of EAU is induced by active immunization with a retinal antigen (Ag) emulsified in complete Freund's adjuvant (CFA), a mineral oil supplemented with heat-killed mycobacteria. In all but the most susceptible mouse and rat strains, an injection of pertussis toxin must be given as an additional inflammatory stimulus (Agarwal and Caspi 2004). This may mimic the putative uveitogenic stimulus that is thought to trigger uveitis in humans, which is believed to involve an exposure to a retinal or crossreactive Ag, combined with an infectious event that provides innate inflammatory danger signals. Uveitogenic retinal proteins include retinal arrestin (soluble Ag), interphotoreceptor retinoid-binding protein (IRBP), rhodopsin, recoverin, phosducin, and retinal pigment epithelium-derived RPE-65. Irrespective of the eliciting Ag, available experimental evidence suggests that the immunological mechanisms driving the resultant disease are similar. Various animal models of uveitis have recently been reviewed (Caspi 2006; Horai and Caspi 2010). Of the available models, the mouse model of EAU induced with IRBP is the best characterized and the most widely used. The typical histological appearance of EAU resembles that of human uveitis, with inflammatory infiltrates in the vitreous, retina, and choroid and damage to the photoreceptor cell layer (Fig. 1).
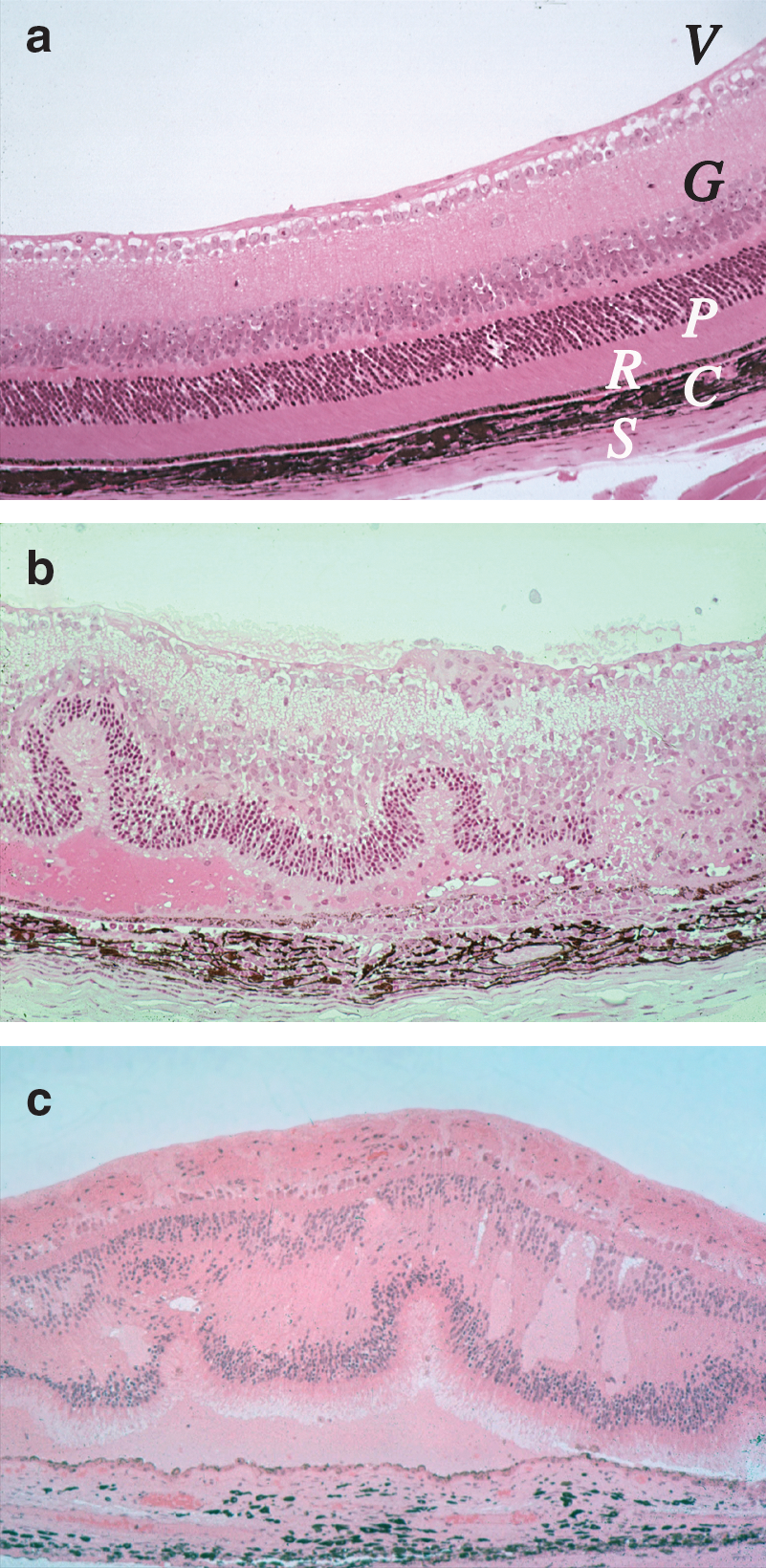
Histological appearance of uveitis in human and mouse
Adaptive/effector T cells from EAU-induced animals can pass the disease to naïve, genetically compatible recipient animals by adoptive transfer. The donor T cells are activated with the immunizing Ag in vitro and are infused into recipient animals. The recipients develop a destructive disease rapidly, usually within a week. The adoptive transfer model allows to avoid the use of adjuvant in the recipients and is useful to analyze the effector mechanism(s) of the disease, mimicking the clinical situation where the patient presents with an immune response that is already ongoing (Caspi 2006; Horai and Caspi 2010).
Recently, we have developed an alternative model to IRBP/CFA-induced uveitis. Dendritic cells (DC) are professional Ag-presenting cells capable of stimulating naïve T cells, and are likely to be the main Ag-presenting cells in the early stages of EAU induction. A model of EAU was developed by injection of matured splenic DC loaded with the major uveitogenic peptide of IRBP into naïve wild-type mice (Tang and others 2007). Compared with the classical EAU model induced by active immunization with IRBP or its peptide in CFA, duration of the disease is shorter, the pathology appears to be less severe, and the inflammatory infiltrate has a predominantly granulocytic rather than mononuclear cell composition. Importantly, EAU elicited with Ag-pulsed DC is not only clinically distinct from CFA-induced EAU, but also is driven by unique effector mechanisms that will be discussed later. This model may offer new insights into the heterogenous nature of human uveitis.
Autoreactive effector CD4+ T cells have been associated with the pathogenesis of inflammatory and autoimmune disorders such as multiple sclerosis, rheumatoid arthritis, Crohn's disease, and uveitis. Naïve CD4+ T cells differentiate into effector subsets depending on the nature of the environment in which exposure to the Ag occurs. Several T cell effector phenotypes have been defined, known as T helper 1 (Th1), Th2, or Th17 and the more recently defined Th9 subset. Early studies suggested that the interferon (IFN)-γ-producing Th1 subset is responsible for the pathology of uveitis, whereas the interleukin (IL)-4-producing Th2 subset is regulatory. More recent studies have lead to a broader paradigm, including the new subsets Th17 and Th9. Each Th subset requires particular cytokines and transcription factors for its differentiation and maintenance, and each has its own cytokine signature, appropriate to its effector function. Proinflammatory cytokines produced by non-T cells are also critical in determining the lineage choice of differentiating Th cells (Fig. 2).
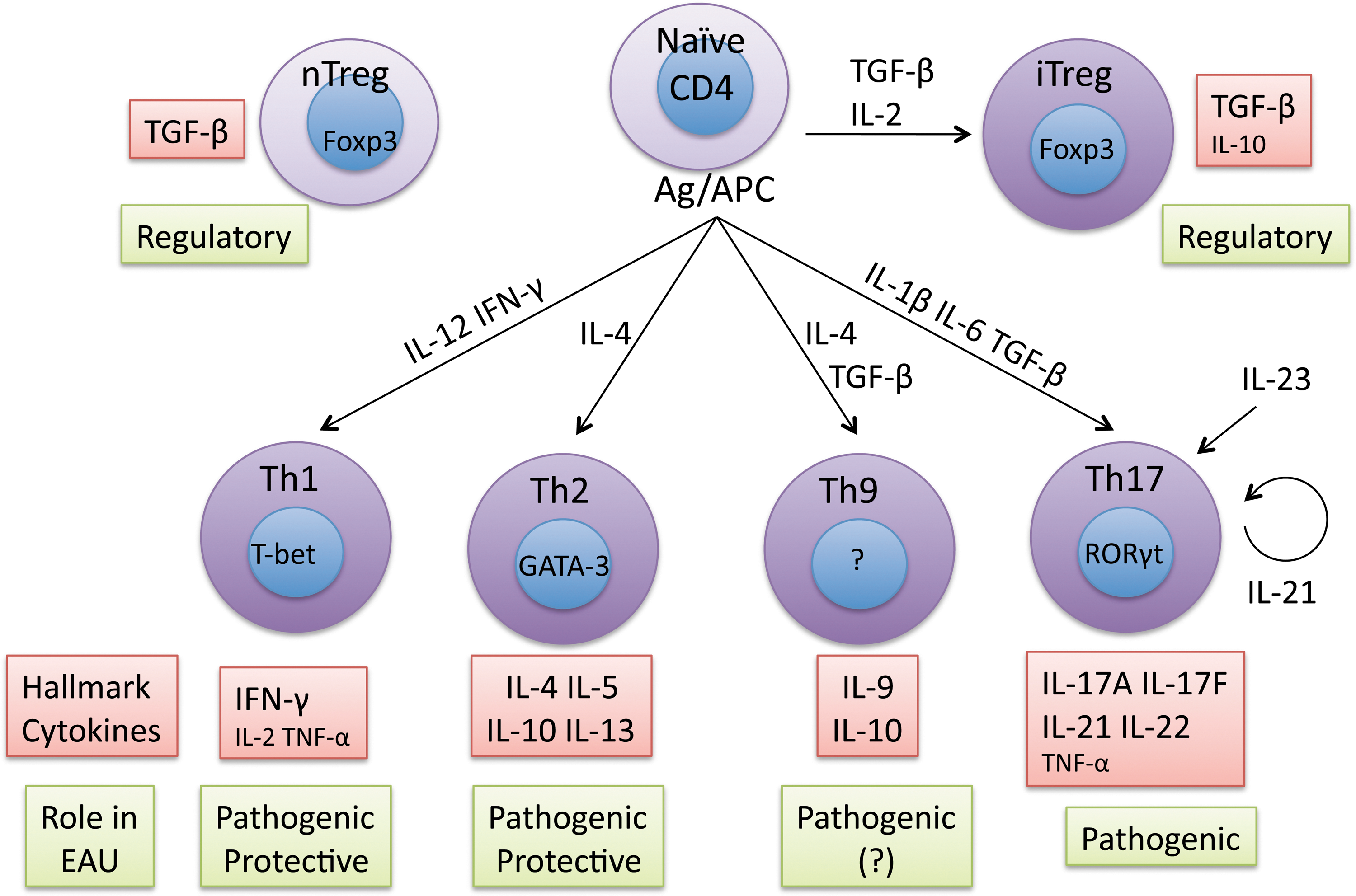
Cytokines and differentiation of effector T cell lineages associated with uveitis. Shown are CD4 T cell differentiation scheme based on currently available information from the literatures and role(s) of each subset in the animal models of EAU. IL, interleukin; TGF-β, transfoming growth factor β; Ag, antigen; TNF-α, tumor necrosis factor α; IFN-γ, interferon-γ; Th cells, T helper cells; Treg cells, regulatory T cells; APC, antigen presenting cells.
Cytokines play an important role in maintaining lymphocyte homeostasis under conditions of health and disease. Intraocular expression of cytokines has been studied in patients with uveitis, with reports of increased levels of inflammatory cytokines and decreased levels of regulatory cytokines (de Boer and others 1992, 1994; Sakaguchi and others 1998; Perez and others 2004; Takase and others 2006). The roles of various cytokines and how they affect the critical checkpoints of uveitis, as studied in animal models and to a lesser extent in patients, are shown in Table 1 and discussed in the following sections.
AqH, aqueous humor; BD, Behçet's disease; VKH, Vogt-Koyanagi-Harada disease; N/A, information not available; EAU, experimental autoimmune uveitis; IFN, interferon; NK, natural killer; IL, interleukin; DC, dendritic cells; NKT, natural killer T cells; PBMC, peripheral blood mononuclear cells; TGF-β, transfoming growth factor β; TNF-α, tumor necrosis factor α; Treg cells, regulatory T cells; APC, antigen presenting cells.
Th1 Cells and Cytokines in Uveitis (IFN-γ and IL-12)
IL-12, composed of 2 heterodimeric subunits, p35 and p40 is produced by DC and macrophages, is a key Th1-inducing cytokine. The roles of IL-12 and of IFN-γ, the main signature cytokine of the Th1 lineage, have been intensively studied in EAU models in the 1990s. At that time, the Th17 subset (discussed ahead) had not yet been described, and the Th1 subset was thought to be the major pathogenic effector T cell subset in uveitis. An IRBP-specific uveitogenic T cell line polarized to the Th1 phenotype in the presence of IL-12 and producing massive amounts of IFN-γ was highly uveitogenic in naïve recipient animals (Xu and others 1997). Contrary to expectations, however, targeting endogenous IFN-γ either by a neutralizing antibody or by genetic deletion did not confer resistance to EAU but rather exacerbated the disease (Caspi and others 1994; Jones and others 1997), suggesting that IFN-γ has a protective role in the pathogenesis of uveitis. Adding to the confusion, more recent data demonstrated that an IFN-γ-producing effector T cell is required for disease induction in the EAU model mediated by Ag-pulsed DC (Tang and others 2007). In the aggregate, these findings show that IFN-γ can be pathogenic or protective, depending on the stage of disease at which it is produced and the model of EAU that is being examined.
The seemingly paradoxical role of IFN-γ as both a protective and a proinflammatory cytokine is clarified by the finding that it is early production of IFN-γ that is protective. Early administration of recombinant IL-12 at the time of immunization for EAU (but not later) prevented the development of disease and dampened the induction of adaptive IRBP-specific responses through an IFN-γ-dependent effect (Tarrant and others 1999). Similarly, IFN-γ elicited from natural killer T cells (NKT) by administration of the invariant T cell receptor ligand α-GalCer (again at the time of immunization, but not later) had a protective role in EAU (Grajewski and others 2008) and inhibited the subsequent adaptive Th1 as well as Th17 responses. Thus, production of IFN-γ early in the response—mostly from innate immune cells—inhibits the subsequent adaptive responses to the uveitogenic Ag. A recently published study identifies yet another potential pathway for protection by early IFN-γ, by identifying granulocyte macrophage colony stimulating factor (GM-CSF) produced by Ag-specific effector T cells as a necessary cytokine for pathology in the central nervous system (CNS), in the model of experimental autoimmune encephalomyelitis (EAE) (Codarri and others 2011). IFN-γ strongly represses the development of GM-CSF producing effector T cells, which may explain why early production of IFN-γ at the time of effector T cell development can confer protection. Although GM-CSF-producing uveitogenic T lymphocytes have not yet been demonstrated to be involved in pathogenesis of uveitis, the EAU and EAE disease models share many mechanisms of inflammatory tissue damage, so this may apply also to EAU.
Th2 and Th9-Associated Cytokines (IL-4, IL-10, IL-13, and IL-9)
Th1 and Th2 responses counter-regulate each other. When the Th1/Th2 dichotomy first emerged in the 1990s and Th1 cells were shown to be pathogenic in uveitis, it was proposed that Th2 cells, as counter-regulatory to Th1, would be protective. A requirement of both IL-4 and IL-10 for induction of protective oral tolerance to retinal Ag in EAU was in line with a regulatory role of these cytokines (Rizzo and others 1999). Further, presence of cells producing IL-4 and IL-10 (hypothesized at that time to be Th2-type) in late-stage EAU was associated with disease resolution (Keino and others 2001), although their function was not directly demonstrated and today we might classify them as regulatory T (Treg) cells rather than Th2 cells. The suppressive function of such Th2-like cells could be ascribed to their IL-10 production, as IL-10 suppresses activation and function of uveitogenic effector cells in vitro and in vivo (Xu and others 1997; Rizzo and others 1998; Agarwal and others 2008). Earlier in the response, Th2 cytokines could affect disease development by tipping the balance away from Th1 development. However, as with Th1, the role of Th2 cells is perhaps not as clear-cut as was first assumed. Genetic resistance of mice to EAU was not clearly associated with their ability to mount a Th2 response (Sun and others 1997). In human uveitis, mixed Th1/Th2 profiles or even elevated levels of Th2 cytokines (IL-4, IL-10, and IL-13) were reported in serum of active Behçet's patients (Raziuddin and others 1998; Aridogan and others 2003), and raised the possibility that the Th2 responses could also be harmful to ocular tissues (Caspi 2002). In keeping with this, adoptively transferred Th2 cells can induce inflammation in the eye, although they were less pathogenic than Th1 and required immunosuppressed hosts (Kim and others 2002). Thus, as with IFN-γ, it is likely that the timing and cellular source of the Th2 cytokines would determine whether Th2 cells may play a pathogenic or a protective role.
Th9 cells are a recently described T cell phenotype that can be induced in presence of IL-4 plus transfoming growth factor β (TGF-β) and that produces IL-9 and IL-10 (Dardalhon and others 2008; Veldhoen and others 2008). The role of Th9 cells was tested for its capacity to induce ocular inflammation in an adoptive transfer model. Although in vitro activated Th9 cells were capable of inducing inflammation in recipient mice, the Th9 phenotype did not seem to be stable in vivo, as expression of IL-9 in the donor cells was not detected in the eye or lymph nodes (Tan and others 2010). Thus, the role of this cytokine and of the cell subset producing it in the pathogenesis of uveitis requires further investigation.
Th17 Lineage Cytokines (IL-17A, IL-17F, IL-21, IL-22, and IL-23)
As part of the contradictory findings concerning the role of IFN-γ in EAU, discussed above, it was also noted that mice lacking the IL-12p35 subunit exhibited exacerbated EAU disease scores, similar to IFN-γ-deficient mice, but surprisingly, the opposite phenotype was observed in the mice lacking the IL-12p40 subunit (Luger and others 2008). This finding and similar findings in other autoimmune disease models were clarified when IL-23, which shares the p40 chain with IL-12, was discovered and shown to be important in differentiation and maintenance of IL-17-producing CD4+ T cells, now known as Th17 cells. In addition to p40, which is shared with IL-12, IL-23 also contains a unique p19 subunit. IL-23p19-deficient mice exhibited protection from EAU, similar to p40-deficient animals (Luger and others 2008). It is now understood that the protection associated with genetic deficiency of the p40 subunit was due to the absence of IL-23 and Th17 responses, and not to the absence of IL-12 and Th1 responses. Th17 cells, which are promoted by IL-23, constitute a more recently identified inflammatory Th subset, and have provided a new understanding of effector mechanisms driving autoimmune diseases (Fouser and others 2008). Although IL-23 was first thought to be needed for initial commitment of T cells to the Th17 lineage, analogous to the role of IL-12 for the Th1 lineage, further studies revealed that IL-23 was not needed for Th17 differentiation from naïve T cells. Rather, Th17 cells are induced by combined effects of IL-6 and TGF-β, or IL-1β and TGF-β, and IL-23 is critical in survival and maintenance of Th17 cells (Bettelli and others 2006; Veldhoen and others 2006; Acosta-Rodriguez and others 2007; Manel and others 2008). Th17 lineage is a heterogenous population that produces IL-17A and IL-17F, IL-21, IL-22, tumor necrosis factor (TNF)-α, and several other cytokines, but not all Th17 cells produce the full complement of these cytokines (Korn and others 2009; Damsker and others 2010). The exact combination of cytokines expressed by Th17 cells may be affected by how they were initially induced and by the cytokine milieu in which they subsequently reside.
Th17 cells may be relevant to human uveitis. IL-23 was shown to be elevated in VKH disease and in Behçet's disease (Chi and others 2007, 2008) and IL-17A production from peripheral blood mononuclear cells (PBMC) was increased in patients with uveitis (Amadi-Obi and others 2007; Chi and others 2008). In animals, IL-17A-producing T cells are associated with the induction of EAU (Peng and others 2007). Neutralization of IL-17A by monoclonal antibodies protects mice from EAU, even when administered after the uveitogenic effector T cells have already been generated (Peng and others 2007; Luger and others 2008; Zhang and others 2009). However, IL-17A-deficient mice still develop EAU with scores only slightly lower than wild-type controls (Luger and others 2008), indicating that compensatory mechanisms exist in IL-17A-deficient animals. These results were confirmed by Yoshimura and others (2008) who additionally noted that the severity of ocular inflammation in IL-17A-deficient mice was initially equal to wild-type controls, but resolved faster after peak of disease had been reached. Importantly, however, in wild-type mice the blockade of IFN-γ and IL-4 in vivo resulted in exacerbation of disease with augmented IL-17 production. These results indicate that, despite existence of mechanisms that can compensate for its absence, IL-17A has a proinflammatory and pathogenic role in the classical model of EAU induced by active immunization of retinal Ag in CFA. In addition to conventional effector CD4+ T cells, IL-17A is also produced by CD8+ T cells (Peng and others 2007) and innate-like cells such as invariant NKT cells (Grajewski and others 2008) as well as γδ T cells (Cui and others 2009). γδ T cells are needed to develop full-blown EAU, but interestingly, treatment with recombinant IL-17 of animals immunized for EAU induction inhibited subsequent disease by suppressing IFN-γ-producing effector T cells (Ke and others 2009). This is reminiscent of the protective role of IFN-γ produced early in the process of EAU development, but at the same time also suggests that the enhancing role of γδ T cells on EAU is not due to their ability to produce IL-17.
The gene encoding IL-17F is adjacent to the gene encoding IL-17A, and IL-17F is coproduced by most of the cells that make IL-17A. In the models or arthritis and EAE, IL-17F is not required for pathogenesis (Iwakura and others 2011). We have observed that IL-17F-deficient mice were equally susceptible to EAU and systemic treatment of animals by anti-IL-17F monoclonal antibody did not protect from EAU, suggesting that IL-17F is dispensable for pathogenesis of EAU as well (Silver and others, unpublished data). In humans, IL-17F gene polymorphisms associated with uveitic diseases have been reported in Asian populations, but because the IL17F gene is closely linked to the IL17A gene, the significance of this finding is unclear (Jang and others 2008; Shu and others 2010).
IL-21 is a member of the IL-2 family of cytokines produced by activated CD4+ T cells and Th17 cells, and is an important factor for induction of follicular helper T cells that are important in eliciting the humoral immune responses (antibody production and germinal center formation). Increased levels of IL-21 in the sera and upregulation of its mRNA in the PBMC of patients with active chronic or recurrent active VKH disease have been reported (Li and others 2010). Animal studies show that IL-21 and IL-21R expression is upregulated in lymph nodes and spleens during EAU (Liu and others 2009) and that IL-21R-deficient mice are resistant to EAU (Wang and others 2011). Thus, IL-21 appears to have a role in promoting uveitis, possibly through increasing Th17 responses.
IL-22 belongs to the IL-10 family. It is largely produced by CD4+ T cells and natural killer (NK) cells and signals through 2 receptors: IL-10Rβ, which is ubiquitously expressed and the heterodimeric receptor IL-22R1, which is restricted to nonlymphoid cells such as epithelial cells and fibroblasts. The role of IL-22 has been controversial in different animal models and diseases. For example, it has been reported to mediate inflammation in arthritis and dermatitis, including psoriasis, whereas it was dispensable for EAE, and was protective in inflammatory bowel disease (Sanjabi and others 2009; Kreymborg and Necher 2010). IL-22 was found to be highly expressed in PBMC of uveitis patients, and in culture could damage primary human retinal pigment epithelial cells by decreasing total tissue resistance and inducing apoptosis (Li and others 2008). On the other hand, recent data indicated a protective role for IL-22 in the mouse EAU model, where disease was ameliorated by treatment with recombinant IL-22. Mechanism of protection appeared to include inhibition of uveitogenic Ag presentation by DC of treated mice (Shao and others 2011). Unpublished data from our laboratory using IL-22-deficient mice and IL-22 neutralizing antibodies also support a protective role of IL-22 in EAU (Mattapallil, Rigden and others, unpublished data). Thus, the role of IL-22 in uveitis may differ between humans and laboratory animals and requires further investigation.
Balance and Interplay of Th1/Th17 Responses Can Determine the Course of EAU
Human uveitis is clinically heterogeneous even though patients often respond to the same retinal Ag(s). Both Th1 and Th17 responses have been associated with human uveitis (Arayssi and Hamdan 2004; Amadi-Obi and others 2007; Chi and others 2007, 2008). Extrapolating from data in animal models, it is conceivable that at least some of those differences may be attributed to differences in the effector T cell lineage driving disease.
As mentioned in Introduction, there exist 2 alternative models of EAU: the classical one induced by immunization with IRBP in CFA (CFA EAU), and a more recently developed model induced by infusion of uveitogenic IRBP peptide-pulsed DC (DC EAU) (Tang and others 2007; Luger and others 2008). Importantly, reminiscent of the heterogeneity of human uveitis, these models differ in severity and clinical course of the disease, in the type of inflammatory infiltrate recruited into the diseased eyes and most importantly, they have a different cytokine dependence. Although both IFN-γ- and IL-17A-producing cells are detected in the inflamed eye, CFA EAU is prevented and reversed by treatment with anti-IL-17A antibodies, indicating dependence on the Th17 effector lineage, and conversely, DC EAU cannot be induced by infusion of uveitogenic DC into IFN-γ-deficient recipients, indicating dependence on the Th1 effector lineage. Thus, autoimmune uveitis can be driven either by a Th1 or by a Th17 response. This notion was further supported by adoptive transfer experiments showing that Th17 and Th1 can each serve as a standalone effector phenotype competent to elicit disease in the absence of the reciprocal effector response in the recipient. Namely, Th1 cells induced disease in hosts treated with anti-IL-17A antibodies, and Th17 cells induced EAU in hosts deficient in IFN-γ (Luger and others 2008). The notion of Th17 cells as standalone effectors might perhaps be blurred by the finding that Th17 cells convert to IFN-γ-producing Th1-like cells in uveitis (Shi and others 2008) and that such conversion may be important for the ability of Th17 cells to induce tissue pathology (Hirota and others 2011).
Nevertheless, the ability of Th17 cells from IFN-γ-deficient donors to induce full-blown EAU in IFN-γ-deficient recipients (Luger and others 2008) argues that IFN-γ itself is not the cytokine that confers pathogenicity on such converted Th17 cells.
Although Th1 as well as Th17 cells induced EAU, each caused recruitment of a different population of inflammatory leukocytes into the eye. Notably, infusion of Th1-polarized cells resulted in a mononuclear infiltrate in the eyes of mice with EAU. In contrast, transfer of Th17-polarized cells resulted in a neutrophilic infiltrate, as also seen in IFN-γ-deficient mice with EAU (Su and others 2007; Luger and others 2008). This is in keeping with the cytokine and chemokine profile typical of the respective responses. IFN-γ is known to attract a mononuclear infiltrate, whereas IL-17 induces IL-8 (or its mouse equivalent, KC) and attracts a neutrophilic infiltrate (Laan and others 1999; Kerr and others 2008).
The dominant effector phenotype driving pathology appears to be influenced by the conditions under which Ag is first recognized by the immune system. In CFA EAU, which is Th17 dependent, the Ag is first recognized in the context of mycobacteria present in CFA. In DC EAU, which is IFN-γ dependent, the Ag is presented by the DC that had been matured with lipopolysaccharide (LPS) and anti-CD40 (Tang and others 2007; Luger and Caspi 2008). This can be duplicated in vitro by stimulating T cells in presence of bone marrow-derived DC matured with bacterial endotoxin (LPS), mycobacterial extract, or the yeast polysaccharide zymosan. T cell receptor stimulation in the presence of LPS-matured DC resulted in a dominant production of IFN-γ by the T cells, whereas mycobacterial extract produced a more IL-17A-dominated response. Zymosan skewed the response even more strongly to IL-17A, with very little IFN-γ production (Fig. 3).
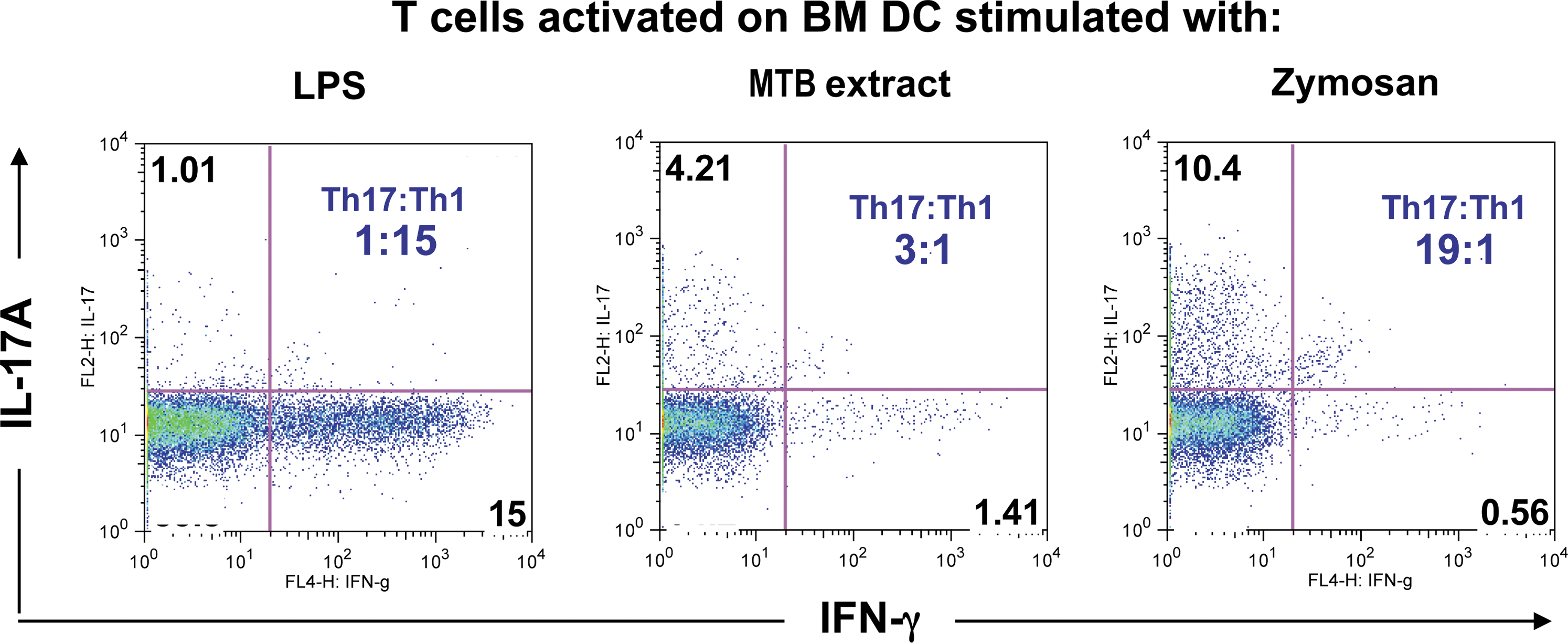
Innate stimulation of APC determines Th1/Th17 balance. Dendritic cells (DC) cultured from bone marrow (BM) using accepted methods were incubated overnight with LPS, Mycobacterium tuberculosis (MTB) extract, or the yeast polysaccharide zymosan. After washing the DC monolayers, sorted naive T cells were added in presence of anti-CD3 Ab for 6 days. Cells were stained for CD4 and intracellularly for IL-17A and IFN-γ after a 4 h PMA: phorbol myristic acetate/Ionomycin and brefeldin A pulse. Shown is cytokine production by CD4+ cells (Isabelle Suffia and others, unpublished data). LPS, lipopolysaccharide; PMA, phorbol myristic acetate.
The clinical and immunological differences between 2 models of uveitis, induced with the same Ag in genetically identical mice under different conditions of innate stimulation, could shed light on the complex and heterogeneous nature of human uveitis and provide a more comprehensive representation of ocular diseases of autoimmune origin. Although no animal model covers full spectrum of human uveitis, each variant has unique characteristics that may contribute to understanding different aspects of human disease.
Other Proinflammatory Cytokines
In addition to IL-12 and IL-23, which are produced by Ag-presenting cells and are key proinflammatory mediators of Th1 or Th17 responses, IL-1, IL-6, and TNF-α are major proinflammatory cytokines produced by various cell types, including lymphocytes, monocyte/macrophages, DC, and ocular resident cells, and play central roles during inflammation. The presence of these cytokines in the ocular fluid of uveitic patients was shown 2 decades ago, and they have been considered as major inflammatory mediators driving pathology at the site of the disease (Wakefield and Lloyd 1992).
The nonredundant requirement for IL-1 in uveitis was demonstrated directly by the complete protection of EAU in mice deficient for type I IL-1 receptor (IL-1RI). In fact, resistance to EAU of mice deficient for MyD88 appeared to be due to lack of IL-1 signaling rather than lack of signaling through MyD88-dependent Toll-like receptors, indicating the pivotal role of this cytokine (Su and others 2005). IL-18, a member of the IL-1 family, also signals through MyD88. IL-18 polymorphisms in BD patients have been reported (Jang and others 2005; Lee and others 2006). However, lack of IL-18 did not affect EAU development (Jiang and others 2001; Su and others 2005).
IL-6 has been shown to be a critical mediator for induction of inflammation. IL-6 has recently been shown to be a critical factor for Th17 differentiation. In uveitis patients, elevated levels of IL-6 were reported in the aqueous humor or vitreous fluid (de Boer and others 1992; Perez and others 2004; Yoshimura and others 2009). Not surprisingly, IL-6-deficient mice were resistant to EAU, and systemic treatment of animals with anti-IL-6R antibody showed suppression of disease, although intraocular treatment was not effective (Yoshimura and others 2009; Hohki and others 2010). Thus, IL-6-dependent Th17 differentiation may play an important part in pathogenesis of EAU.
The inflammatory role of TNF-α has been demonstrated in many autoimmune diseases, including arthritis, psoriasis, and Crohn's disease. The serum level of TNF-α is elevated in patients with active Behcet's disease (Evereklioglu and others 2002; Lee and others 2003). Neutralization of TNF-α in the EAU model was effective in suppressing disease (Sartani and others 1996; Dick and others 1998). TNFR1-deficient mice were resistant to EAU, because TNF signaling is required for macrophage migration to inflammatory site (Raveney and others 2009). It was also reported that TNF-α, as well as vascular endothelial growth factor and IL-1β, may contribute to the breakdown of the blood–retinal barrier (BRB) in EAU and in patients with uveitis, possibly through opening of tight junctions and increased vesicular transport within the endothelial cells (Luna and others 1997). Thus, TNF-α has been a major target for treating inflammatory and autoimmune diseases, including uveitis as described ahead.
Anti-Inflammatory/Regulatory Cytokines
Regulatory cytokines play a critical role in modulating activation of lymphocytes and controlling inflammation. TGF-β, IL-10, IL-27, and IL-35 have all been shown to have suppressive activity in autoimmune diseases.
TGF-β is a pleiotropic cytokine that shows regulatory as well as inflammatory activity, depending on the context of other cytokines present in the same environment (Sanjabi and others 2009). For the regulatory function, TGF-β is a key factor to induce Treg cells that are important for peripheral tolerance. In combination with IL-2, TGF-β promotes differentiation of induced Treg cells (Chen and others 2003). On the other hand, TGF-β was also recently identified as a critical cytokine for Th17 and Th9 differentiation when acting in concert with other cytokines (IL-1β or IL-6 for Th17 and IL-4 for Th9) (Veldhoen and others 2006, 2008). TGF-β is abundant in ocular fluids, where it is found mostly in the form of TGF-β2. In uveitic patients, mature TGF-β levels in aqueous humor were reported to be reduced (de Boer and others 1994), its lack conceivably promoting disease and lending support to its potentially regulatory function. TGF-β suppresses the acquisition of effector functions by autopathogenic T cells (Xu and others 2003). Recent data from our group demonstrated that TGF-β together with retinoic acid, which is also abundant in the eye due to its role in the chemistry of vision, coordinately support in situ conversion of naïve T cells into Treg cells within the ocular microenvironment (Zhou and others, unpublished data).
IL-10 was already mentioned above in the context of the Th2 response; however, IL-10 can be associated not only with the Th2 response. IL-10 is an immunomodulatory cytokine produced by various cell types, including conventional and Treg cells, B cells, and monocytes. Recently, studies identified IL-10 production in cells that otherwise show Th1 and Th17 phenotype, raising the concept that IL-10 production may be a general regulatory mechanism by which effector T cells mitigate their own inflammatory activity and control bystander tissue damage (Anderson and others 2007; Jankovic and others 2007; Xu and others 2009). Elevated levels of IL-10 were detected in the serum or ocular fluid in the patients with uveitis (Aridogan and others 2003; Takase and others 2006). In the model of EAU, treatment of mice with IL-10 after EAU induction ameliorated disease scores, and systemic neutralization of IL-10 during the effector phase enhanced EAU, supporting its protective role in uveitis (Rizzo and others 1998). Moreover, mice with transgenic overexpression of IL-10 in activated T cells or in macrophages constitutively were resistant to EAU (Agarwal and others 2008). Because of its anti-inflammatory activity, modulation of IL-10 or its signaling components can be an attractive therapeutic approach.
IL-27 is a recently identified cytokine of the IL-6/IL-12 family. It is composed of 2 subunits, p28 and Epstein-Barr virus (EBV)-induced gene-3 (Ebi-3). There is still a paucity of information about this cytokine in EAU. IL-27 receptor is a heterodimer composed of a unique IL-27R chain (WSX-1) and gp130, which is shared between IL-6 and IL-27 receptors. Sonoda and others (2007) reported that WSX-1-deficient mice developed reduced EAU scores, suggesting a pathogenic role of IL-27R signaling in ocular autoimmunity. In apparent contradiction, however, Amadi-Obi and others (2007) reported that IL-27 is constitutively expressed in retinal ganglion and photoreceptor cells and inhibits Th17 responses in EAU. Further, microglial cells in the eye constitutively secrete IL-27, whose expression is further upregulated during EAU, and photoreceptors constitutively express IL-27R and respond to IL-27 signals by producing IL-10 and suppressor of cytokine signaling proteins (Lee and others 2011), implying a protective role for IL-27 in ocular inflammation. These findings are in line with the results of others in CNS inflammation models, indicating that IL-27 is protective in EAE by regulating pathogenic Th17 cells (Batten and others 2006; Stumhofer and others 2006). Thus, similarly to the dual role of IFN-γ, IL-27 in EAU may also be pathogenic or protective, depending on when and where it is produced. It is conceivable that the apparently pleiotropic effects of IL-27 may in part be related to its ability to induce IFN-γ and promote the Th1 response.
Recent studies identified the IL-27p28 subunit of IL-27 as an antagonist of gp130 (Stumhofer and others 2010). Further clarification is needed as to whether the suppressive role of IL-27 in autoimmunity is related to the antagonistic effect of IL-27p28. In support of this notion, recent unpublished data from our laboratory demonstrated that systemic overexpression of p28 was protective in Th1- and in Th17-induced EAU models (Chong and others unpublished data).
IL-35 is a newly discovered cytokine produced by Treg cells as a result of cell–cell contact with their target cell. IL-35 is a heterodimer composed of 2 subunits: p35 (shared with IL-12) and Ebi-3 (shared with IL-27) (Collison and others 2007). IL-35 was shown to attenuate disease in the model of arthritis (Niedbala and others 2007). Although the role of IL-35 has not yet been reported in EAU, a suppressive function for this cytokine is predicted by the finding that mice lacking p35 showed exacerbated EAU (Luger and others 2008). In this context, it would be of interest to examine EAU susceptibility in mice deficient in the other IL-35 chain, EBI-3, but such studies had not been reported. However, it must be kept in mind that mice deficient in either of these subunits lack more than one cytokine, which may confound interpretation of the results.
Emerging Treatments of Clinical Uveitis Targeting Cytokines and Their Receptors
Established therapies for uveitis are based largely on nonspecific immunosuppression (corticosteroids, antimetabolites, and alkylating agents). However, because of the severe side effects of these treatments, it is important to develop new approaches based on increased understanding of basic disease mechanisms, so as to intervene more specifically in the pathogenic processes. Although involvement of many cytokines has been demonstrated in experimental uveitis (Fig. 4), targeting their activities clinically in humans is still quite limited. One of the hazards involves the pleiotropic nature of some cytokines and the possibility to elicit unexpected reactions. As an example, a clinical trial to treat multiple sclerosis that was undertaken on the basis of early data in mice showing that IFN-γ can have protective effects in EAE (similarly to EAU) resulted in exacerbation of the disease and had to be stopped (Panitch and others 1987).
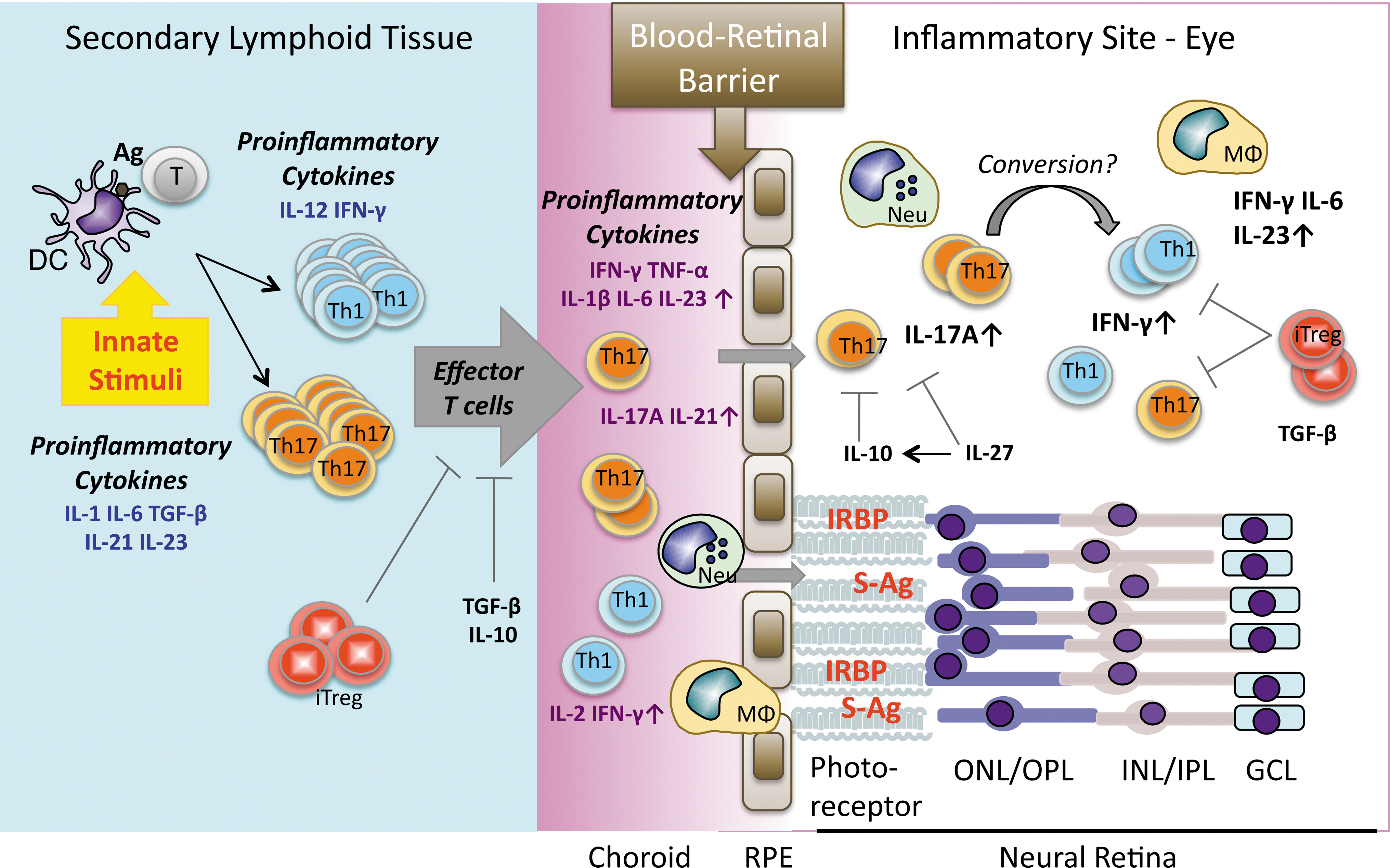
Cytokine networks in pathogenesis of uveitis. Presentation of Ag in the periphery in the presence of cytokines and innate (environmental) stimuli induces T cell activation, differentiation, and clonal expansion. Activated effector (uveitogenic) T cells then migrate and extravasate into the eye. Upon breakdown of blood–retinal barrier (BRB), leukocytes and lymphocytes (Th1 and Th17) that are recruited from circulation, as well as inflammatory cytokines, amplify the inflammation, resulting in uveitis. RPE, retinal pigment epithelium; ONL, outer nuclear layer; OPL, outer plexiform layer; INL, inner nuclear layer; IPL, inner plexiform layer; GCL, ganglion cell layer.
That said, the central involvement of T cells in pathogenesis of uveitis makes them a natural target for therapy. Activated T cells depend on IL-2 for their proliferation. Cyclosporin A (CsA), a T cell targeting drug of the macrolide family that blocks the IL-2 signaling pathway by inhibiting calcineurin, is now in general use for ocular inflammation. CsA was first shown to have a therapeutic effect in the rat EAU model before going to clinical trials (Nussenblatt and others 1985). The macrolides FK-506 (tacrolimus) and rapamycin (sirolimus) also target the IL-2 signaling pathway and are effective for some types of uveitis (Yang and others 2009). More recent studies have examined IL-2 receptor-directed therapy with monoclonal antibodies (daclizumab) as an approach to target activated T cells. This therapy has shown efficacy in advanced clinical trials (Nussenblatt and others 1999; Nussenblatt 2002; Yeh and others 2008). Interestingly, the possibility that such treatment might actually aggravate T cell-mediated autoimmunity because IL-2 is necessary for the maintenance and activity of Treg cells (at least in mice) was not fulfilled. The mechanism behind the therapeutic effects of daclizumab is complex and incompletely understood, but includes an enhancement in CD56-bright NK cells with inhibitory function (Li and others 2005). Unfortunately, despite its demonstrated efficacy in a number of clinical conditions, daclizumab has recently been discontinued due to insufficient market demand (
Other cytokine-based interventions currently in use for uveitis include neutralization of TNF-α or of IL-1, as well as augmentation of IFN-α. Interestingly, they seem to be particularly effective in uveitis accompanying Behçet's disease. TNF-α neutralization (infliximab, adalimumab, and etanercept) has been used for some time to treat various autoimmune diseases (Reimold 2002). Infliximab, a chimeric murine-human IgG1, reduced inflammation in uveitis that was refractory to conventional therapy (Jap and Chee 2008). In patients with Behçet's disease who had reduced numbers of circulating FoxP3+ Treg cells, successful infliximab treatment was accompanied by an increase in Treg cells (Sugita and others 2011). Inhibition of IL-1 signaling by antibodies to IL-1β or by recombinant human IL-1R antagonist (Anakinra, an endogenous inhibitor of IL-1 activity) also was effective in Behçet's patients (Dinarello 2011). Lastly, in analogy to IFN-β in multiple sclerosis, IFN-α has been effective in uveitis related to Behçet's disease. IFN-α has been approved in Europe (although not yet in the United States) for this indication (Imrie and Dick 2007).
Since the eye is a small and relatively closed organ, local therapies in the eye are an attractive approach that can obviate systemic side effects. Intravitreal injections or implants are already in use for such local therapies, and biological products can also be delivered into the eye. Locally produced IL-10 has been shown to be beneficial in animal models (Broderick and others 2005; Smith and others 2005). This opens the possibility for intraocular injection of other anti-inflammatory molecules, such as IL-27 and IL-35, or even in vitro generated Treg cells. For this purpose, it will be important to develop minimally invasive and highly efficient local drug delivery approaches.
Conclusions
Because human autoimmune uveitis is heterogenous and access to clinical material is limited, it has been difficult to dissect the basic mechanisms involved. EAU models have served as extremely useful platforms to understand the mechanisms that might drive uveitis in humans. Recent advances with the discovery of new cytokines and T cell subsets, as well as advances in cytokine biology, have shed light on effector mechanisms of uveitis at the cellular and molecular levels. Although results from recent studies suggest a pathogenic role of Th17 cells in uveitis, data from animal models of uveitis indicate that both Th1 and Th17 cells are pathogenic through production of their lineage-specific effector cytokines. Based on the current knowledge, targeting specific cytokine molecules or their signaling pathways could ultimately help in developing better treatment strategies for human uveitis.
Footnotes
Acknowledgment
This work has been supported by NIH/NEI Intramural Funding.
Author Disclosure Statement
No competing financial interests exist.