
Abstract
Oat β-glucan can counteract the increased risk for Herpes Simplex Virus 1 (HSV-1) infection in mice, the effects of which have, at least in part, been attributed to macrophages. However, the specific responses of macrophages to oat β-glucan treatment in this model have yet to be elucidated. We examined the effects of varying doses of oat β-glucan on the pro-inflammatory cytokine response in both peritoneal and lung macrophages with and without exposure to HSV-1 infection in vitro. Peritoneal and lung macrophages were obtained from mice and cultured with varying concentrations of oat β-glucan (0 (control), 10, 100, and 1,000 μg) for 24 h and supernatants were collected. A standardized dose of HSV-1 was added for a second 24 h incubation period after which supernatants were again collected. Samples were analyzed for interleukin-1β (IL-1β), IL-6, and tumor necrosis factor α (TNF-α) using enzyme linked immunosorbent assay (ELISA). In most cases, oat β-glucan resulted in a dose-dependent increase in pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α) in lung and peritoneal macrophages with and without exposure to HSV-1 infection. When comparing across macrophage source, this response was greater for IL-1β and IL-6 in peritoneal macrophages and for TNF-α in lung macrophages. This may be a mechanism for the decreased risk for HSV-1 infection following oat β-glucan feedings in mice.
Introduction
Soluble oat β-glucan is a structural polysaccharide (∼2×106 molecular weight) found in the cell walls of the bran layer and endosperm fractions of the whole seed. Structurally, it not only consists of linear chains of β-
The purpose of this study was to examine the effects of varying doses of oat β-glucan on the release of pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α) from both peritoneal and lung macrophages with and without exposure to HSV-1 infection in vitro. Our data are the first to indicate that oat β-glucan can increase the release of pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α) from both peritoneal and lung macrophages in response to HSV-1 infection in vitro.
Materials and Methods
Macrophage cultures
Macrophages were obtained from 6-week-old male ICR mice, purchased from Harlan Sprague Dawley. Peritoneal macrophages were harvested by washing the peritoneal cavities of 20 mice (2 mice were pooled to give a sample size of 10) with 5 mL of RPMI 1640 (Gibco BRL, Grand Island, NY) supplemented with 10% fetal bovine serum and 2% penicillin, streptomycin, and
Oat β-glucan
Following adherence of macrophages, 200 μL/well of RPMI 1640 supplemented with 10% fetal bovine serum and 2% penicillin, streptomycin, and
HSV-1 infection of macrophages
Following treatment of macrophages with oat β-glucan, 50 μL of a standardized dose of HSV-1 KOS strain (7–10 pfu/cell) contained in RPMI 1640 supplemented with 10% fetal bovine serum and 2% penicillin, streptomycin, and
Cytokine protein analysis
Enzyme linked immunosorbent assay (ELISA) was used to measure IL-1β, IL-6, and TNF-α in the macrophage culture supernatants (R&D Systems, Minneapolis, MN). The assay was performed according to manufacturer's instructions. Results were reported as pg/mL.
Statistical analysis
Statistical analyses were performed using a commercially available statistical package from SigmaStat (version 2.03, SigmaStat; SPSS, Chicago, IL). Differences in cytokine release across oat β-glucan dose in both the preinfection and postinfection cultures were compared using a one-way analysis of variance (ANOVA) with Student-Newman-Keuls post hoc analysis (P<0.05). To compare the postinfection cytokine levels to preinfection levels a one-way ANOVA was performed on the values of the difference (post–pre) to determine if the increase in cytokine release following HSV-1 infection was affected by oat β-glucan. Data are presented as mean (±SEM).
Results
Peritoneal macrophages
Interleukin-1β
We examined IL-1β release from peritoneal macrophages following culture with various doses of oat β-glucan [0 (control), 10, 100, and 1,000 μg] (Fig. 1A). Statistically significant elevations were found for 10 μg (∼100 pg/mL), 100 μg (∼300 pg/mL), and 1,000 μg (∼250 pg/mL) versus 0 μg (0 pg/mL; P<0.05). Further, both 100 μg and 1,000 μg increased IL-1β levels compared with the 10 μg dose (P<0.05). We then cultured these macrophages with HSV-1 and again examined the cytokine response. Our data indicate increases in IL-1β release for 10 μg (∼300 pg/mL), 100 μg (∼900 pg/mL), and 1,000 μg (∼900 pg/mL) compared with 0 μg (0 pg/mL; P<0.05; Fig. 1A) with the largest increase observed for 100 μg and 1,000 μg (P<0.001). We also compared the difference between pre- and postinfection (post–pre) cytokine levels to determine if the increase in inflammation following HSV-1 infection was affected by oat β-glucan. Statistically significant differences were seen for 10 μg (∼200 pg/mL), 100 μg (∼600 pg/mL), and 1,000 μg (∼650 pg/mL) versus 0 μg (0 pg/mL; P<0.001). And both 100 μg and 1,000 μg increased IL-1β levels versus the 10 μg dose (P<0.001).
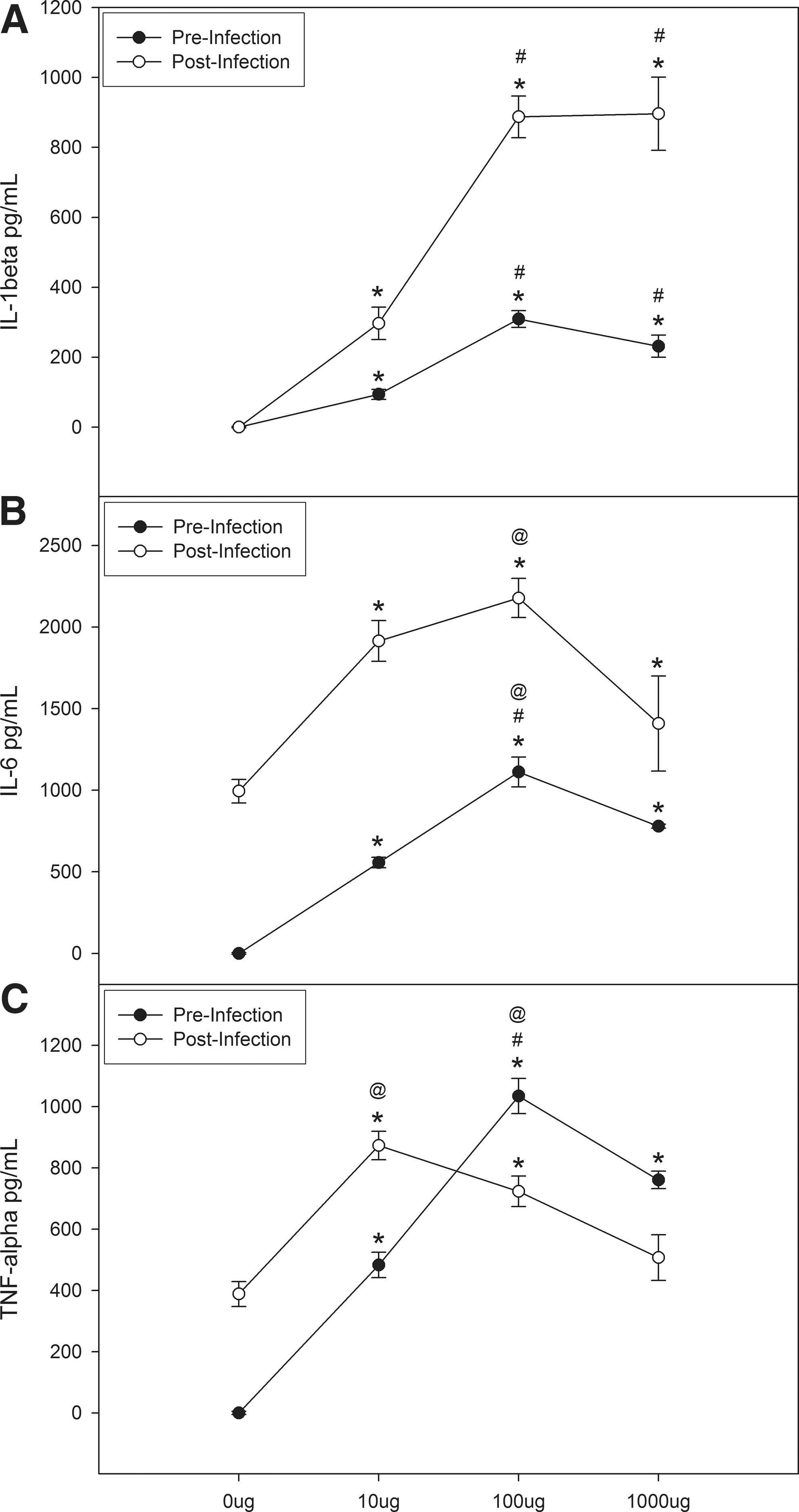
Oat β-glucan increases interleukin 1β (IL-1β)
Interleukin-6
A similar response was found for IL-6 release (Fig. 1B); significant differences were found for 10 μg (∼600 pg/mL), 100 μg (∼1,200 pg/mL), and 1,000 μg (∼800 pg/mL) compared to 0 μg (0 pg/mL; P<0.05), and maximal increases were observed with 100 μg (P<0.05). Following infection with HSV-1, significant differences were found for 10 μg (2,000 pg/mL), 100 μg (2,200 pg/mL), and 1,000 μg (1,400 pg/mL) versus 0 μg (1,000 pg/mL; P<0.05) with the greatest elevation observed at the 100 μg dose (P<0.001; Fig. 1B). The response following HSV-1 infection (post–pre) was affected by only the 10 μg dose (∼1,400ρg/mL versus ∼950ρg/mL (0 μg)).
Tumor necrosis factor-α
For TNF-α, elevations were found for 10 μg (∼500 pg/mL), 100 μg (∼1,000 pg/mL), and 1,000 μg (∼800 pg/mL) compared with 0 μg (0 pg/mL; P<0.001; Fig. 1C) and the greatest increases were seen with 100 μg and 1,000 μg (P<0.001). After infection with HSV-1, significant increases were found for 10 μg (∼900 pg/mL) and 100 μg (∼750 pg/mL) versus 0 μg (∼400 pg/mL; P<0.001) and maximal cytokine release was observed at the 10 μg dose (P<0.05; Fig. 1C). When we examined the difference (post–pre) in cytokine levels across oat β-glucan dose we found that both 0 μg (∼400ρg) and 10 μg (∼400ρg) had increased TNF-α release versus 100 μg and 1,000 μg.
Lung macrophages
Interleukin-1β
In lung macrophages, only the 1,000 μg (∼10 pg/mL) dose of oat β-glucan resulted in a significant increase in IL-β versus 0 μg (0 pg/mL; P<0.001); however, the magnitude of this response was much less than that observed for peritoneal macrophages (Fig. 2A). Similarly, following culture with HSV-1, IL-1β release was increased at the 1,000 μg (20 pg/mL) concentration versus 0 μg (0 pg/mL; P<0.001; Fig. 2A) and again the extent of this response was much less than in peritoneal macrophages. Likewise, we found that the 1,000 μg concentration had elevated levels of IL-1β compared with the other doses (P<0.05) after analyzing the difference between pre- and postinfection levels.
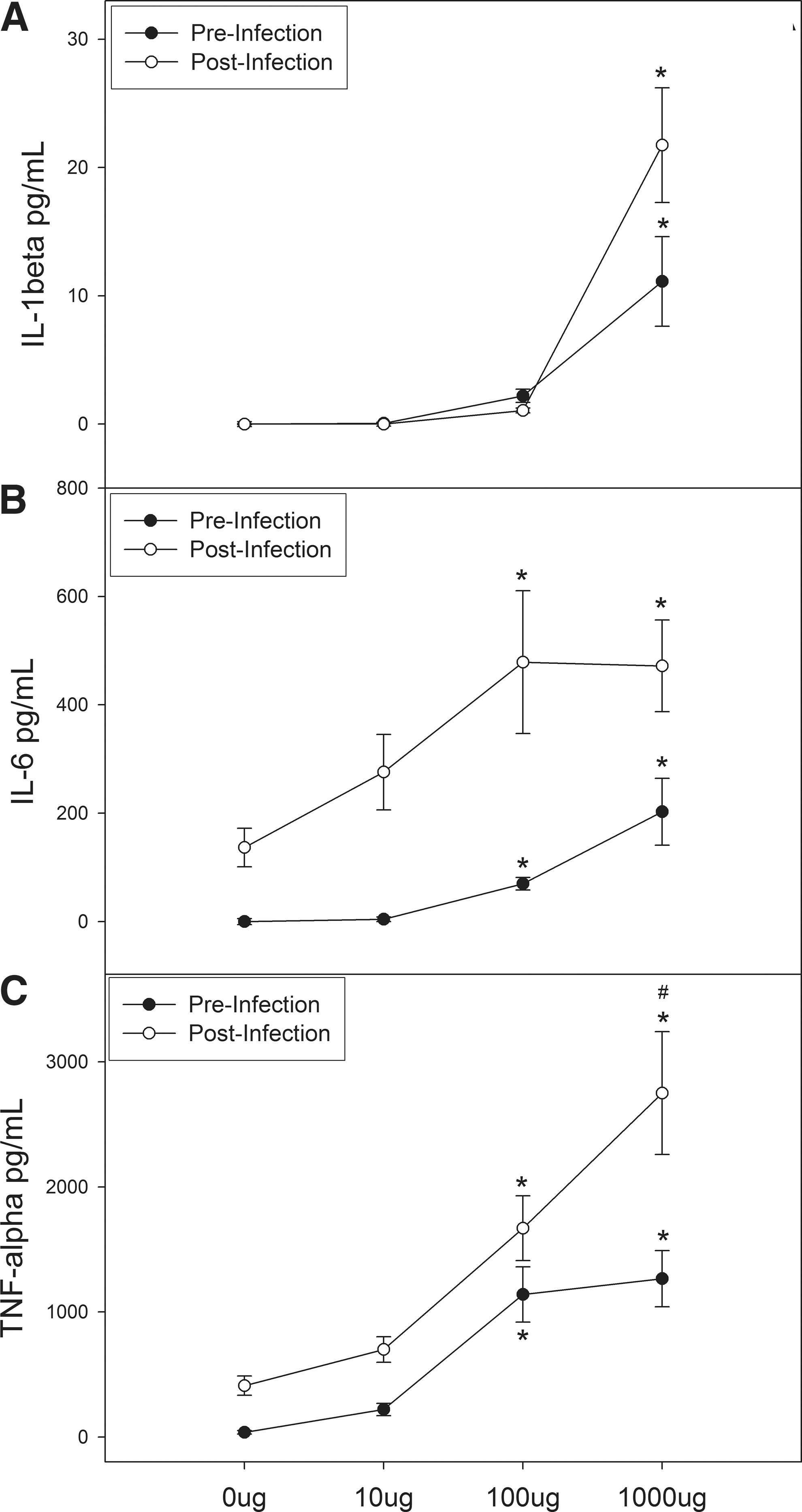
Oat β-glucan increases IL-1β
Interleukin-6
For the IL-6 response in the lungs, statistically significant increases were found for 100 μg (∼75 pg/mL) and 1,000 μg (∼200 pg/mL) versus 0 μg (0 pg/mL; P<0.05; Fig. 2B). When HSV-1 infection was added to the culture, we found elevated levels of IL-6 with the 100 μg (500 pg/mL) and 1,000 μg (500 pg/mL) doses (P<0.05) versus 0 μg (∼150 pg/mL; Fig. 2B); however, as seen with IL-β, the magnitude of this response was much less than that observed for peritoneal macrophages. We found no significant differences in the oat β-glucan response after comparing the difference between pre- and postcytokine levels.
Tumor necrosis factor-α
Finally, for TNF-α release from lung macrophages (Fig. 2C) significant increases were found with the 100 μg (∼1,100 pg/mL), and 1,000 μg (∼1,300 pg/mL) doses compared with 0 μg (0 pg/mL; P<0.001). And contrary to what was observed for IL-1β and IL-6 in lung macrophages, the overall magnitude of this response appeared greater than that observed for peritoneal macrophages. A similar response was found following infection with HSV-1; 100 μg (∼1,600 pg/mL) and 1,000 μg (∼2,800 pg/mL) increased TNF-α compared with 0 μg (∼400 pg/mL; P<0.001) and maximal elevation was observed with the 1,000 μg dose (P<0.05; Fig. 2C), and again this response was greater than that of peritoneal macrophages. The post–pre analysis indicates that the increase in TNF-α following addition of HSV-1 is influenced by the 1,000 μg (∼1,800ρg) dose of oat β-glucan but not by the 10 or 100 μg doses.
Discussion
Previous work with soluble β-glucan derived from oats suggests that it can enhance resistance to various viral, protozoan, and bacterial infections that is mediated, at least in part, by its ability to activate macrophages (Estrada and others 1997; Yun and others 1998, 2003; Davis and others 2004b; Murphy and others 2008a). We have shown that oat β-glucan feedings can offset the increase in susceptibility to HSV-1 respiratory infection that occurs with stressful exercise (Davis and others 2004b), the benefits of which are lost following depletion of lung macrophages (Murphy and others 2008a). However, the specific antiviral responses of macrophages following treatment of oat β-glucan in this model or other virus challenge models are largely unknown. Likewise, there is no information regarding possible differences in the macrophage response in various tissue locations. These are the first data to indicate that oat β-glucan can increase the release of pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α) from both peritoneal and lung macrophages under control conditions and in response to HSV-1 infection in vitro. Further, we report that this response varies according to the macrophage source; the response was greater in peritoneal macrophages for both IL-1β and IL-6 and in lung macrophages for TNF-α.
The first part on this study examined the effects of varying doses of oat β-glucan on the release of pro-inflammatory cytokines from both lung and peritoneal macrophages in vitro. Macrophages have been demonstrated to possess β-glucan receptors such as CR3 and dectin-1 that recognize and mediate phagocytosis of glucan particles (Goodridge and others 2009). Following encounter with β-glucans, macrophages can enhance their transcription and translation of a wide range of gene products (Mueller and others 2000). For example, it has been shown to activate nuclear factor-κB (NFκB) in macrophages and trigger its nuclear translocation (Xu and others 2011). In this study, the pro-inflammatory cytokine (IL-1β, IL-6, and TNF-α) response to oat β-glucan was increased, generally in a dose-dependent manner, in both peritoneal and lung macrophages. When comparing across macrophage source the response appeared to be greater in peritoneal macrophages for both IL-1β and IL-6 and in lung macrophages for TNF-α. Plasticity is a well-known property of macrophages (Holt and others 2008; Galli and others 2011; Lopez-Castejon and others 2011), therefore it is possible that macrophages at different tissue locations may not display the same magnitude of change in response to a similar stimulus. Our findings are consistent with those of Estrada and others (1997) that report an oat β-glucan-induced increase in IL-1β and TNF-α from peritoneal macrophages in vitro. However, this is the first study to examine the pro-inflammatory cytokine response of macrophages to oat β-glucan treatment from 2 different tissue sources and the first report of this response in lung macrophages.
The second part of this study sought to examine the effects of varying doses of oat β-glucan on the release of pro-inflammatory cytokines from macrophages following HSV-1 infection in vitro. This was done to begin to examine the specific antiviral responses of macrophages that may play a role on the benefits of oat β-glucan on susceptibility to respiratory infection (Davis and others 2004b); pro-inflammatory cytokines have been implicated in playing a necessary role in the pathogenesis of viral infections, including HSV-1, and macrophages are primary sources of these mediators (Van Reeth 2000; Mogensen and others 2004; Murphy and others 2008b, 2011b; Iijima and others 2011; Shi and Pamer 2011). In fact, we recently reported an increase in susceptibility to HSV-1 respiratory infection in IL-6 knockout mice; IL-6 knockout mice showed an increase in morbidity, mortality, and lung viral titers that was associated with a decrease in macrophage antiviral activity (Murphy and others 2008b). Further, both IL-1β and TNF-α are produced by virally infected macrophages and interact with various cells of the immune system to eliminate viral infections (Van Damme and others 1987). However, to date there are no reports on the influence of oat β-glucan on the pro-inflammatory macrophage cytokine response to any viral pathogen. Our findings show for the first time that incubation of peritoneal and lung macrophages with oat β-glucan results in an increase in the release of pro-inflammatory cytokines in response to HSV-1 infection. Consistent with the first part of this study, a greater response occurred for IL-1β and IL-6 in peritoneal macrophages and for TNF-α in lung macrophages. As previously stated, these findings are not unusual given the plasticity of macrophages (Holt and others 2008; Galli and others 2011; Lopez-Castejon and others 2011).
It is worth noting that in general, the macrophage cytokine response appeared greater in the supernatants from infected cells versus the noninfected cells for both peritoneal and lung macrophages. To compare the postinfection cytokine levels to preinfection levels we analyzed the values of the difference (post–pre) to determine if the increase in cytokine release following HSV-1 infection was affected by oat β-glucan. Our data indicate that this response was influenced by treatment, although it varied according to the dose. However, these data should be interpreted with caution as the same cells were used for both noninfected (first 24 h incubation) and infected treatment (second 24 h incubation) as opposed to parallel cultures. It is also important to point out that, for peritoneal macrophages in particular, there appeared to be an inhibition of cytokine release in both noninfected and infected samples with the highest dose of oat β-glucan. While we did not explore this finding the most likely explanation is that the 1,000 μg dose of oat β-glucan may have resulted in some toxicity.
Given the necessary role of inflammation in resistance to HSV-1 infection (Mogensen and others 2004; Murphy and others 2008b, 2011b) an increase in pro-inflammatory cytokines from macrophage cells during infection, in conjunction with other known alterations in macrophage functions including intrinsic antiviral resistance, may contribute to the decrease in susceptibility to infection that has been associated with oat β-glucan consumption in rodents (Davis and others 2004a, 2004b). However, it is important to note that resistance to infection was not examined in this study and therefore conclusions on whether the oat β-glucan induced increase in pro-inflammatory cytokines can specifically alter susceptibility to HSV-1 infection cannot be made. In addition, the mechanisms for the increase in pro-inflammatory cytokines were not examined. Previous studies using β-glucan from other sources have implicated NFκB and other transcription factors in this response (Xu and others 2011). Although no studies of this nature have been carried out with β-glucan derived from oats it is likely to affect the same or similar pathways. Future investigations should examine specific pathways by which oat β-glucan can stimulate cytokine release in this model and production of other cytokines that may influence susceptibility to HSV-1 infection. Further, the macrophage cytokine response to other viral pathogens or macrophage activators were not examined; therefore, the observed effects of oat β-glucan cannot be generalized to other pathogens or compared with other known macrophage activators.
In conclusion, this study suggests that oat β-glucan can increase the macrophage pro-inflammatory cytokine response to HSV-1 infection in vitro, the magnitude of which varies according to the source of macrophages. This along with other known changes in macrophage function may be a mechanism for the decreased susceptibility to viral infection following orally administered β-glucan feedings in mice.
Footnotes
Acknowledgment
This work was funded by a grant from the American College of Sports Medicine.
Author Disclosure Statement
No competing financial interests exist.