
Abstract
The present study evaluated any possible association among Interleukin-1-beta (IL-1B)-511 and IL-1 receptor antagonist (IL-1RN) variable number tandem-repeat (VNTR) genotypes, haplotypes, and IL-1B expression with risk for hepatitis B virus-related hepatocellular carcinoma (HBV-HCC) development in India. For this, 406 subjects (153 controls, 67 inactive HBV-carriers, 65 patients with chronic-active HBV, 62 HBV-cirrhotics, and 59 subjects with HBV-HCC) were enrolled in the study. Polymerase chain reaction (PCR)–restriction fragment length polymorphism, reverse transcriptase–PCR, and enzyme-linked immunosorbent assay methods were used for assessing polymorphism, mRNA, and protein levels, respectively, of IL-1. The study revealed no significant association of IL-1B(−511) CT and TT genotypes, while a significant positive association of the IL-1RN (VNTR) 1/2 genotype with HCC development, among controls and carriers. Besides, 2/2 genotypes acted as a potential risk factor for hepatitis and subsequent cirrhosis development, among the same groups. Furthermore, the IL-1 haplotypes 2 and 3 were found to be significant protective factors for hepatitis and subsequent HCC development, among controls. However, haplotype 4 shared a significant negative association with hepatitis only. Moreover, proinflammatory IL-1B levels significantly and steadily elevated with the disease progression to HCC, as compared to controls. These preliminary findings indicate a key role of IL-1 in the HBV-mediated disease chronicity, in the Indian population.
Introduction
A
IL-1 is a proinflammatory Th1 cytokine with multiple biological effects. The IL-1 gene family (including IL-1α, IL-1β, and IL-1 receptor antagonist (IL-1RN) is located on chromosome 2q13.21, spanning a region of 430 kb (Um and others 2003), encoding 3 proteins, which comprise the agonists IL-α, IL-β, and their naturally occurring inhibitor, IL-1 receptor antagonist (IL-1Ra/IL-1RN) (Dinarello 1996). The IL-1Ra gene (IL-1RN) contains an 86-bp variable number tandem-repeat (VNTR) polymorphism in intron 2 (Hutyrova and others 2002), and it acts as a competitive inhibitor (Tarlow and others 1993). The polymorphisms are located within the regulatory regions of the genes and are, therefore, of potential functional importance by modulating IL-1 protein production (Belfer and others 2004). IL-1B, because of its interaction with IL-6 and tumor necrosis factor (TNF), plays a vital role in central and peripheral immune response. A biallelic (C/T) polymorphism in the (Interleukin-1beta) IL-1B promoter region (position−511) was identified, which affects IL-1B secretion in vitro (Pociot and others 1992). This marker was found to be associated with susceptibility to numerous malignancies such as gastric cancer (El-Omar and others 2000; Ito and others 2007), pancreatic cancer (Barber and others 2000), breast carcinoma (Jin and others 1997), esophageal, and colorectal carcinomas (Ito and others 2007), but the association results have been inconsistent across different studies. While studying the relationships between polymorphism of the IL-1β promoter region −511C/T and the IL-1RN gene polymorphisms and susceptibility to chronic hepatitis B in the Chinese population, an association was reported between these polymorphisms and chronic hepatitis B infection (Zhang and others 2004). Recently, allele 2 of the IL-1RN intron 2 has been reported as a resistant marker of HBV infection, suggesting the role of the IL-1 polymorphisms in the pathogenesis of developing chronic hepatitis B. This allele is associated with enhanced IL-1β production in vitro and in vivo. Therefore, Zhang and others hypothesized that high production of IL-1β may help increase the production of other cytokines such as IL-2, IL-6, and TNF-α and trigger the complex immunological processes to eliminate the virus. The −511C allele of the IL-1β gene was identified as a genetic marker for the development of HCC in patients with chronic HBV infection. This was supported by an analysis that revealed that the IL-1B −511 C allele was significantly increased in patients with HCC in comparison to controls (Hirankarn and others 2006). However, literature is scanty regarding the association of IL-1B and IL-1RN (VNTR) with hepatitis B disease progression from asymptomatic carrier to the carcinoma state worldwide, with absolutely no reports from India. Thus, the present study was designed to evaluate role of the IL-1B(−511C>T) and IL-1RN (VNTR) polymorphism, mRNA, and the protein levels with the HBV-HCC risk in India.
Materials and Methods
Subjects
A total of 406 subjects were enrolled in this study.
Group 1 included 153 healthy subjects showing no clinical signs and symptoms, to suggest any form of disease, and underwent HBsAg screening so were designated as the control group. The subjects of this group were the relatives or the attendants of the patients attending the Hepatology out patient department (OPD) at the Postgraduate Institute of Medical Education and Research, Chandigarh, India. Group 2 comprised of 67 inactive HBV carriers with HBsAg positive for a period of 6 months or more, but with normal levels of transaminases, were HBeAg negative or anti-HBe positive, had serum HBV DNA <105 copies/mL and no clinical symptoms of liver disease along with no radiological evidence of cirrhosis or varices on endoscopy. Group 3 consisted of 65 patients with chronic-active HBV characterized by HBsAg positive for a period of 6 months or more along with raised transaminases >1.5×upper limit of normal (ULN) and had serum HBV DNA more than 105 copies/mL and imaging techniques showing no evidence of cirrhosis or no varices on endoscopy. Group 4 included 62 HBV-related cirrhotics with HBsAg positive for a period of 6 months or more along with raised transaminases >1.5× ULN and had serum HBV DNA more than 105 copies/mL and imaging techniques showing evidence of cirrhosis or varices on endoscopy, and ascites is detectable by ultrasonography. Group 5 included 59 subjects with HBV-HCC characterized by concomitant finding of 2 imaging techniques showing nodule larger than 2 cm with arterial hypervascularization or by one positive imaging technique showing hypervascularization associated with a-fetoprotein levels higher than 400 ng/mL (Zhou and others 2006). Also, the serum HBV DNA was more than 105 copies/mL. Groups 2, 3, 4, and 5 comprised of subjects attending the Hepatology OPD at the Postgraduate Institute of Medical Education and Research, Chandigarh, India.
Exclusion criteria (Groups 2–5)
Following patients were excluded from the study if the (1) Patient was HIV +ve (2) Patient was anti HCV +ve (3) Patient had any other viral marker in blood or any evidence of hemochromatosis and Wilson's disease.
The protocol of the study was approved by the Institute's Ethics Committee. Informed consent was obtained from all the subjects involved in the study. Six milliliters of venous blood was collected from all the subjects.
DNA isolation
High-molecular-weight genomic DNA was isolated from the peripheral blood samples using the standard sodium perchlorate method as described by Daly and others (1996) and was suspended in a TE buffer (pH 8.0; 10 mM Tris and 1 mM EDTA) and finally used for IL-1B and IL-1RN (VNTR) genotyping.
Genotyping of IL-1B(−511)
The IL-1B(−511C>T; rs16944) polymorphism was determined by a polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP)-based method (Zhang and others 2004). The primers used were as follows: IL-1B(−511) forward primer: 5′-GTTTAGGAATCTTCCCACTT-3′; IL-1B(−511) reverse primer:5′ TGGCATTGATCTGGTTCATC-3′ (Sigma-Aldrich Pvt. Ltd.). Briefly, the genomic DNA (100 ng) was amplified in a total volume of 10-μL reaction mixture consisting of 1×PCR buffer(50 mM KCl; 1.5 mM MgCl2; 20 mM Tris HCl, pH8.4), 2 mM MgCl2, 0.6U Taq DNA polymerase, (MBI Fermentas, Life Sciences, USA), 1 μM primers, and 200 μM each dNTP. The PCR conditions consisted of initial denaturation at 94°C for 2 min, 35 amplification cycles with denaturation at 94°C for 45 s, annealing at 55.7°C for 45 s, and elongation at 72°C for 45 s. Final extension was carried out at 72°C for 10 min. Each set of reaction included an internal (β-actin), positive, and negative controls. To detect the C→T variant at position −511, the PCR amplification product (304 bp) was digested with 1U of Ava1 (37°C for 1 h) and then electrophoresed on a 3% agarose gel. On RFLP analysis of IL-1B(−511), the presence of 2 bands of 190 bp and 114 bp depicted the wild genotype (CC), and the uncut band of 304 bp showed the variant (TT) genotype. However, the detection of 3 bands of 304 bp, 190 bp, and 114 bp after restriction digestion confirmed the presence of a heterozygous (CT) genotype. β-actin was used as the internal control and amplified as a 350-bp product.
Genotyping of the IL-1RN penta-allelic VNTR polymorphism
The polymorphism analysis of IL-1RN (VNTR) (Fig. 2) resulted in amplicons of 5 different sizes depending on the number of repeats: 240 bp=2 repeats (allele 2), 325 bp=3 repeats (allele 4), 410 bp=4 repeats (allele 1), 500 bp=5 repeats (allele 3), and 595 bp=6 repeats (allele 5). Further, for the purpose of statistical analysis and because of the rarity of alleles 3, 4, and 5, this polymorphism is treated as biallelic by dividing alleles into short and long categories, in which the short allele has 2 repeats (allele 2), and the long allele has 3 or more repeats (alleles 1, 3, 4, and 5) (Machado and others 2001). β-actin was used as the internal control and amplified as a 350-bp product.
Haplotype and linkage disequilibrium analysis
The construction and analyses of haplotypes of each individual were inferred using the PHASE algorithm (Stephens and others 2001), which uses a Bayesian approach incorporating a priori expectations of haplotypic structure based on population genetics and coalescence theory. Haploview version 2.0.2 (Whitehead Institute for Biomedical Research) was used to estimate linkage disequilibrium (LD). The LD was expressed as Lewontin's D (ID'I) and the LD coefficient r2, between all pairs of biallelic loci.
Measurement of blood IL-1B levels
Peripheral venous blood was collected in sterile heparinized tubes, and peripheral blood mononuclear cells (PBMCs) were isolated by using the standard Histopaque (Sigma) density-gradient centrifugation method (Boyum 1984). Their viability was assessed by trypan dye exclusion test and were later stored at −80°C. PBMCs were lysed in PBS through repeated freeze–thaw cycles before the determination of the IL-1B levels (105 cells/well) using an enzyme-linked immunosorbent assay (ELISA) kit (BD OptEIA™ Set Human IL-1B; BD Biosciences Pharmingen). Sensitivity of the kit was 3.9 pg/mL.
Reverse transcriptase–PCR analysis of IL-1B
Total RNA was isolated from PBMCs (2×105 cells) by using a HiPurA™ Total RNA Miniprep Purification Spin kit (HiMedia Laboratories Pvt. Ltd.), and cDNA synthesis was carried out from the purified and intact total RNA (100 ng) according to the manufacturer's instructions (RevertAid™ First Strand cDNA Synthesis Kit; MBI Fermentas, Life Sciences). IL-1B expression was evaluated by real-time reverse transcriptase (RT)–PCR analysis by using sequence-specific primers. Ten microliters PCR mixture consisted of 5 μL of 1×Absolute QPCR SYBR Green Mix (consisting of 3 mM MgCl2 and DNA polymerase), 4 μL reaction mixture consisting of 0.3 μM of both forward and reverse primers (Sigma-Aldrich Pvt. Ltd.), and water, to which 1 μL of cDNA was finally added. In optimized PCR, the initial denaturation step was carried out for 2 min at 95°C. The PCR conditions used are as follows: initial denatuation for 15 min at 95°C; amplification (40 cycles): 30 s at 94°C, 30 s at 64°C (IL-1B) or 68°C (β-actin) annealing °C; synthesis: 20 s at 72°C; melting (1 cycle): 1 s at 95°C, 1 min at 73°C/66°C, and continuous melting at 95°C. The sequences of the IL-1B (Lichtenstein and others 1997) and β-actin (Primer3 Input v 0.4.0) primers used are shown in Table 1. The expected PCR products of size 249 bp and 318 bp were obtained for IL-1B and β-actin (internal control), respectively, when electrophoresed on a 2% agarose gel.
Statistical analysis
Genotype frequencies were calculated as the number of participants with a particular genotype divided by the total number of participants. The Hardy–Weinberg equilibrium was also tested. Based on the genotype distribution data, power of the study was found to be more than 80%.
The statistical significance was depicted by using Pearson P-value. Association between genotypes/haplotypes with HCC risk was calculated as odds ratios (ORs) with 95% confidence intervals (CIs), and their statistical significance was determined by chi-square test using SPSS (v.15 for Windows) and PHASE v2.1 software, respectively. Further, the OR was adjusted by logistic regression analysis. The nonparametric Kruskal–Wallis one-way analysis of variance and Student's t-test were used to determine the significance of differences in continuous variables using GraphPad Prism software, while multiple comparisons were done by Bonferroni test. Pearson's correlation analysis was used to estimate the correlations between different parameters. The association of independent variables with cytokine concentration was assessed by multiple regression analysis using SPSS software. A P-value under 0.05 was considered statistically significant.
Results
On analyzing the demographic characteristics of the subject groups, the subjects found to have HBV-HCC showed the highest mean as well as median age with respect to the other categories, showing that the HBV-HCC is a disease of later half of life. Moreover, an increased gender ratio (male: female) revealed a distinct male preponderance for HBV-HCC in the present study in all the mentioned groups (data not shown).
Also, the liver function test profile depicted that nearly all the investigated parameters (aspartate and alanine aminotransferase and alkaline phosphatase and albumin/globulin ratio) were significantly elevated as the disease progresses to cirrhosis, with reference to the controls, thus reflecting an enhancement in the degree of disease severity (data not shown).
IL-1B(−511C>T) polymorphism analysis
In Fig. 1, the PCR-RFLP products of IL-1B(−511) of some of the representative samples are shown having a random distribution of the 3 genotypes viz. wild, heterozygous, and variant.
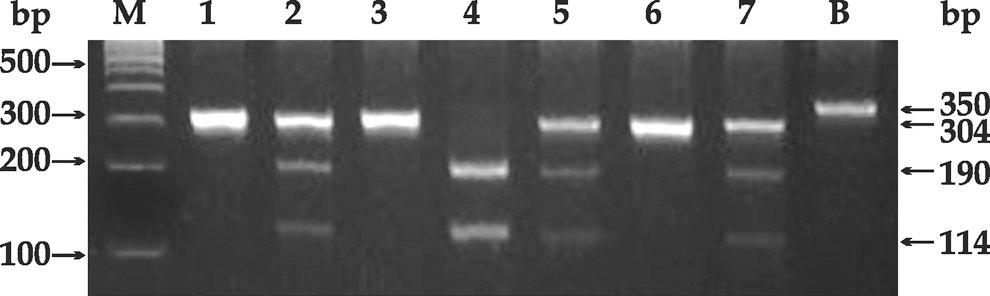
Polymerase chain reaction (PCR)–restriction fragment length polymorphism analysis of IL-1B(−511) of some representative samples. The PCR product was digested with 1U of Ava1 (37°C for 1 h) and electrophoresed on a 3% agarose gel. In Lane M: 100-bp ladder; lanes 1, 3, and 6: variant genotype (304 bp); lanes 2, 5, and 7: heterozygous genotype (304 bp, 190 bp, and 114 bp); lane 4: wild genotype (both 190 bp and 114 bp); Lane B: β-actin amplification product (350 bp).
Genotype distribution of IL-1B(−511C>T)
The genotype distribution analysis of the polymorphic forms of the IL-1B(−511) (Table 2) showed that the CT genotype was the most prevalent genotype in all the investigated groups. Besides, the IL-1B (CC) genotype was maximally present in the HBV-cirrhotics (9.68%), was comparable in case of inactive HBV-carriers (7.81%) and subjects with chronic-active HBV (7.69%), while it was present least in subjects with HBV–HCC (3.39%). Further, the subjects with HBV-HCC showed the highest distribution (54.24%) for the IL-1B (TT) genotype in comparison to the other groups. Also, this distribution was significantly more (P<0.01) than the minimal frequency of 32.26%, present in HBV-cirrhotics. Besides, the incidence of IL-1B(CT) genotype distribution was almost equivalent in inactive HBV carriers (48.44%) and the subjects with chronic-active HBV (49.23%), while it was found to be least in the HCC group (42.37%). Moreover, the findings were consistent with the Hardy–Weinberg rule.
Data represented as number of subjects (genotype distribution,%).
P<0.01 w.r.t. HBV-cirrhotic.
HBV, hepatitis B virus; HCC, hepatocellular carcinoma; w.r.t., with respect to.
Association of the IL-1B(−511C>T) genotypes with the HBV-HCC risk
The OR for the association between IL-1B(−511) genotypes and the carcinoma progression (Table 3) showed that both the CT and TT genotypes were found to be nonsignificantly associated with HBV-HCC development, among controls and carriers.
Data represented as odds ratio (95% confidence interval).
ORa=unadjusted odds ratio and ORb=odds ratio adjusted with age, gender, bilirubin, total protein, A/G, AST, ALT, and ALP.
IL-1RN (VNTR) polymorphism analysis
The PCR analysis of the IL-1RN (VNTR) as depicted in Fig. 2 showed the presence of different allelic combinations of the 86-bp sequence. A total of 5 different alleles can be seen at 240 bp (allele2), also termed as the short allele, while the long allele comprises 325 bp (allele4), 410 bp (allele1), 500 bp (allele3), and 595 bp (allele5) representing 3, 4, 5, and 6 copies of the 86-bp sequence in intron 2 of the IL-RN gene. PCR was performed to look for the distribution of the different allelic combinations of the 86-bp sequence. β-actin was used as the internal control and amplified as a 350-bp product.
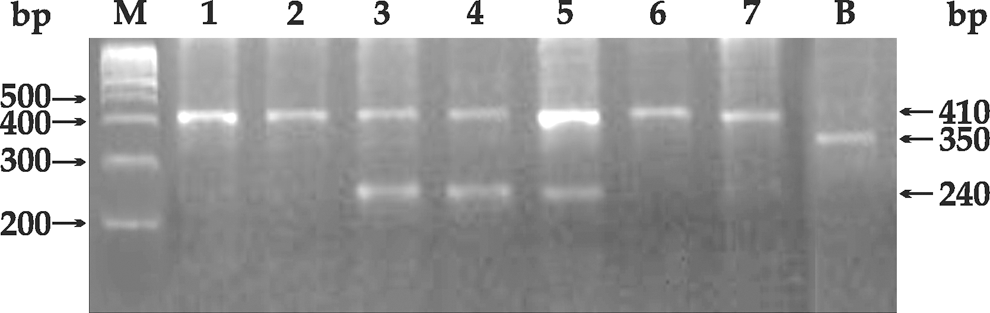
PCR-analysis of IL-1RN (variable number tandem repeat) of some representative samples. In Lane M: 100-bp ladder; Lanes 1, 2, 6, and 7: 1/1 (410 bp) genotype; Lanes 3–5: 1/2 (410 bp and 240 bp) genotype; Lane B: β-actin amplification product (350 bp). IL-1RN, interleukin-1 receptor antagonist.
Genotype distribution of IL-1RN(VNTR)
The genotype distribution for the IL-1RN (VNTR) polymorphic forms (Table 4) showed that among all the genotypes, 1/1, 1/2, and 2/2 forms were the most prevalent ones in all the categories. Further analysis revealed that the 1/1 genotype has a maximum frequency in the inactive HBV-carrier subjects (62.69%) as compared to the controls (52.94%), subjects with chronic-active HBV(28.13%), HBV-cirrhosis (33.33%), and HBV–HCC (37.29%). Also, the frequency distribution observed in the control population was significantly more, in comparison to the subjects with hepatitis (P<0.001), cirrhosis (P<0.01), and HCC (P<0.05). In the hepatitis B group, 56.25% of the subjects showed the presence of the 1/2 genotype, which is significantly higher than controls (38.23%; P<0.05) and HBV-carriers (20.89%; P<0.001). Also, the controls had significantly more 1/2 genotype distribution (P<0.01), with respect to (w.r.t.) carriers. Besides, with 20.89% of 1/2 genotype distribution, the carrier subjects were observed to have significantly lesser distribution in comparison to the cirrhotic (46.67%; P<0.01) and the HCC (50.85%; P<0.001) categories. However, the 1/3 genotype was observed to be maximal in the carriers (2.98%), while it was comparable (2.20%) in the control population and minimal (1.56%) in the hepatitis group. The 1/5 genotype was found to be present only in HBV-carrier subjects (2.98%). The 2/3 genotype was almost equivalently present in the carrier (1.50%) and HCC groups (1.69%). Moreover, the 2/2 genotype was found to be maximum in the HBV-cirrhotic group (20%), which was found to be significantly more as compared to the controls (5.15%; P<0.001) and the hepatitis (14.06%; P<0.05) groups. Also, this genotype was significantly more in the hepatitis category (P<0.05), as compared to the control population.
Data represented as number of subjects (genotype distribution,%).
P<0.05, ** P<0.01, and *** P<0.001; w.r.t. control; + P<0.05, ++ P<0.01, and +++ P<0.001 w.r.t. inactive HBV-carrier.
Association of the IL-1RN (VNTR) genotypes with HBV-HCC risk
The OR for the association between the IL-1RN (VNTR) genotypes and the carcinoma progression taking 1/1 genotype as reference (Table 5) showed that the 1/2 genotype shared a significant positive association (P<0.05 and P<0.001, respectively) with HBV-HCC development with the control and HBV-carrier groups as reference, the later being at ∼2-fold higher risk as compared to controls. Besides, 2/2 genotype was found to be a potential risk factor for hepatitis (P<0.01) and subsequent HBV-cirrhosis development (P<0.001), both among controls and carriers.
Data represented as odds ratio (95% confidence interval).
OR a=unadjusted odds ratio and ORb=odds ratio adjusted with age and gender.
P<0.05, ** P<0.01, and *** P<0.001 w.r.t. control ++ P<0.01 and +++ P<0.001 w.r.t. inactive HBV-carrier.
Haplotype distribution of IL-1B(−511) and IL-1RN(VNTR)
The haplotype distribution for IL-1B(−511) and IL-1RN (VNTR) (Table 6) showed that hap1 is maximally distributed in the chronic-active HBV (23.8%; P<0.001) and in the subjects with HCC (15.80%; P<0.05), which was significantly more in comparison to the controls(5.20%). While in case of cirrhotics, the haplotypic frequency of 6.80% was found to be significantly less w.r.t. the hepatitis B (P<0.001) and HCC (P<0.05) groups. As far as hap2 is concerned, it was found that the frequency of 30.80% in controls and 33.10% in inactive carriers was observed to be significantly higher as compared to the hepatitis (9.80%; P<0.001) and the subjects with HCC (7.90%; P<0.001). Also, the HBV-cirrhotics were found to have more incidence of the hap2 (29.70%), when compared to the hepatitis B (P<0.001) and HBV-HCC (P<0.001) groups. In the control population, 27.90% frequency of hap 3 was observed, which was higher than that of the carriers (15.30%; P<0.01) and the chronic-active HBV (18%; P<0.05) categories. On the contrary, in the cirrhotics, a frequency of 36.40% was found to be maximum when compared to the carriers (15.30%; P<0.001), the hepatitis (18%; P<0.001), and HCC (22.80%; P<0.05) groups. Moreover, the distribution of hap 4 was found to be 36.10% in controls, which was significantly less than that of the HBV-carrier (50.80%; P<0.01), hepatitis B (48.40%; P<0.05), and the HBV-HCC (53.50%; P<0.01) categories.
Data represented as number of chromosomes examined (haplotype distribution,%). *2=short allele (2 repeats) and L=long allele (3–6 repeats).
P<0.05, ** P<0.01, and *** P<0.001 w.r.t. control; +++ P<0.001 w.r.t. inactive HBV-carrier; ^^^ P<0.001 w.r.t. chronic-active HBV; # P<0.05 and ### P<0.001 w.r.t. HBV-cirrhotic.
Association of IL-1B(−511C>T) and IL-RN (VNTR) haplotypes with HBV-HCC risk
The OR for the association between the IL-1B(−511) and IL-1RN (VNTR) haplotypes and the carcinoma progression (Table 7), with all wild haplotypes as reference, showed that haplotypes 2 and 3 were found to be significant protective factors (P<0.001) for chronic-active hepatitis and subsequent HBV-HCC development (P<0.001 and P<0.05, respectively), with controls as reference. However, as far as haplotype 4 is concerned, it was found to be significantly in negative association (OR=0.30; P<0.05) with the subjects with hepatitis only. To further investigate that whether the 2 SNPs act as independent predictors of disease risk/protection or effect each other (as a consequence of LD), the LD was estimated. The LD analysis between the IL-1B(−511) and IL-1RN (VNTR) loci, done separately in control and the diseased populations, showed a weak LD (D′=0.04; r 2=0.001 and D′=0.206; r 2=0.01), respectively, in the two.
Data represented as odds ratio (95% confidence interval).
ND=not determined due to a single subject having this haplotype.
ORa=unadjusted odds ratio and ORb=odds ratio adjusted with age and gender.
P<0.05 and *** P<0.001 w.r.t. control.
Analysis of IL-1B levels in various categories
On assaying the spontaneous/unstimulated levels of IL-1B in the PBMCs (Fig. 3), a significant (P<0.001) and steady increase was evident in the IL-1B levels from a median of 38.94 pg/mL in the control population to 179.1 pg/mL in subjects with HCC. Besides, the levels found in the HCC group were also found to be significantly higher as compared to the carriers (87.08 pg/mL; P<0.01) and the chronic-active HBV category (110.41 pg/mL; P<0.01).
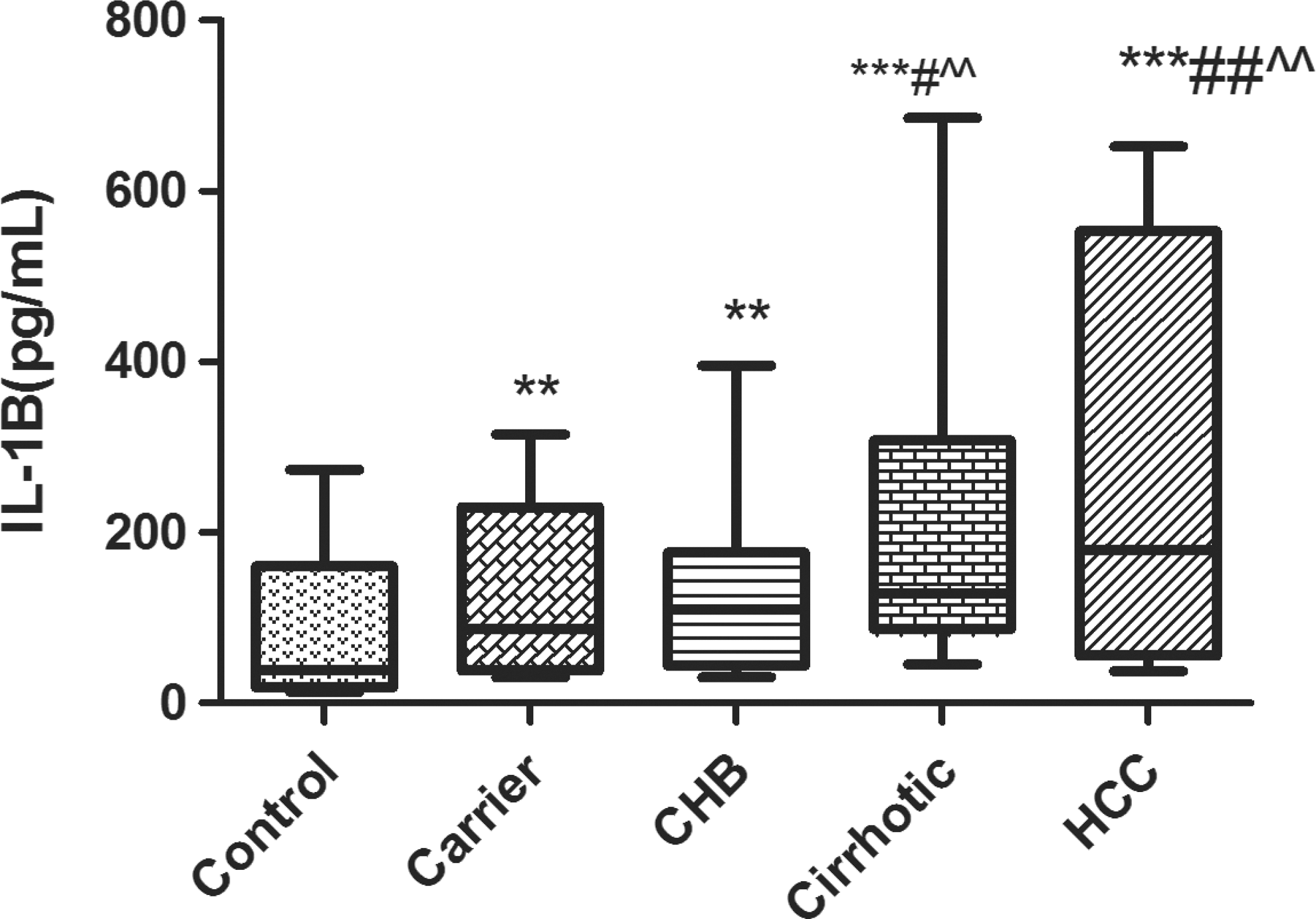
IL-1B levels in various categories. Box and whisker plot depicting median value (middle line) and whiskers showing minimum to maximum values. **P<0.01 and ***P<0.001 w.r.t. control; # P<0.05 and ## P<0.01 w.r.t. inactive HBV-carrier; ^^ P<0.01 w.r.t. chronic-active.
Moreover, as shown in Fig. 4, the IL-1B levels (median±SEM) were found to be markedly decreased in subjects with the variant genotype (11.69 pg/mL; P<0.01), as compared to the CT genotype individuals (35.08±22.7 pg/mL), in the control group. However, HBV-carriers and the diseased categories displayed no significant changes in the levels, among all the 3 genotypes. On Pearson correlation analysis of IL-1B levels, with different parameters (IL-1B genotypes, bilirubin, total protein, AST, ALT, ALP, and A/G), none of them were found to be significantly correlated with IL-B.
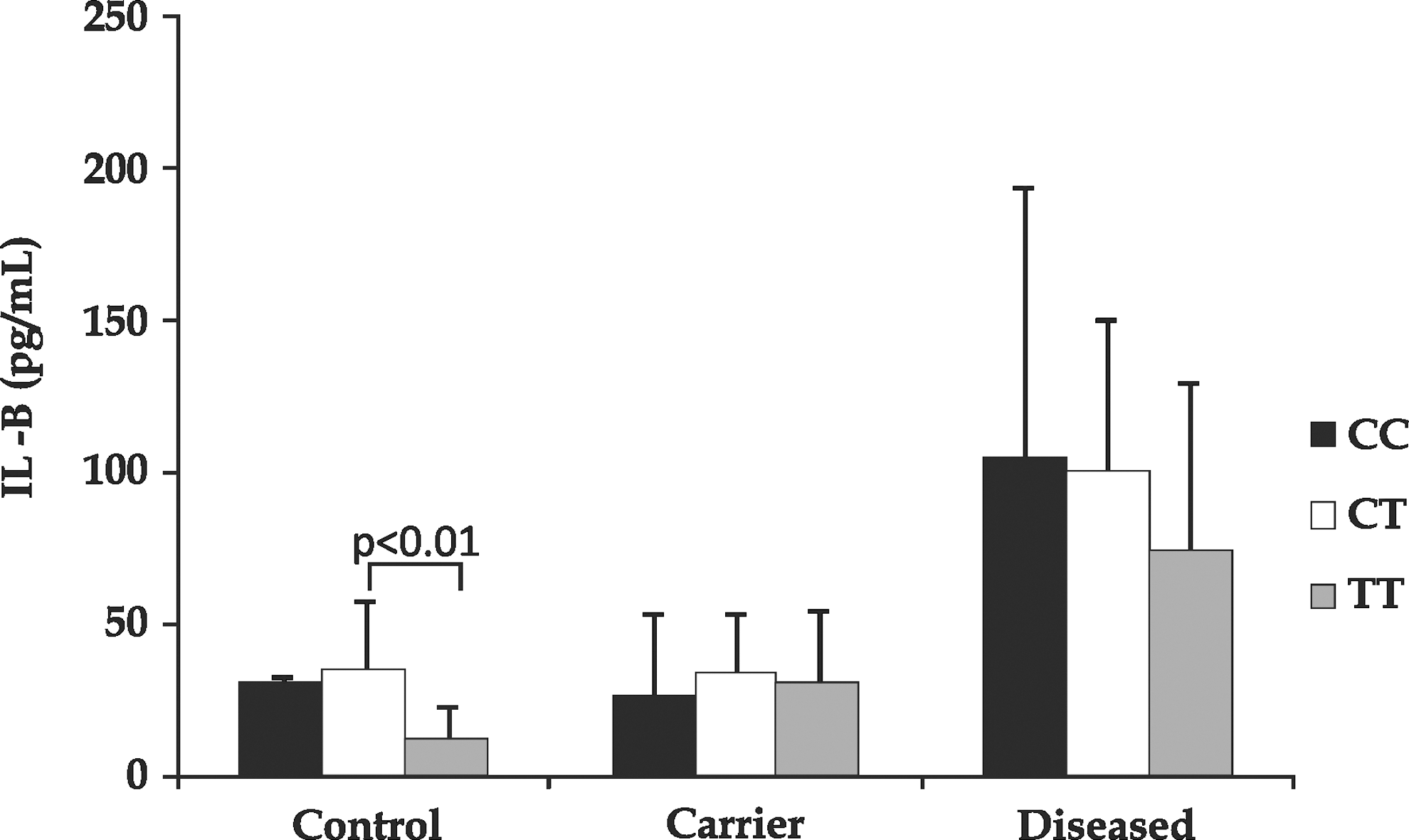
IL-1B levels in control, carrier, and the diseased groups with different IL-1B(−511C/T) genotypes. Data are represented as (median±SEM) pg/mL.
RT-PCR analysis of IL-1B
The mRNA expression (normalized with β-actin) of IL-1B when studied by RT-PCR (Fig. 5) revealed no significant change, in any of the categories.
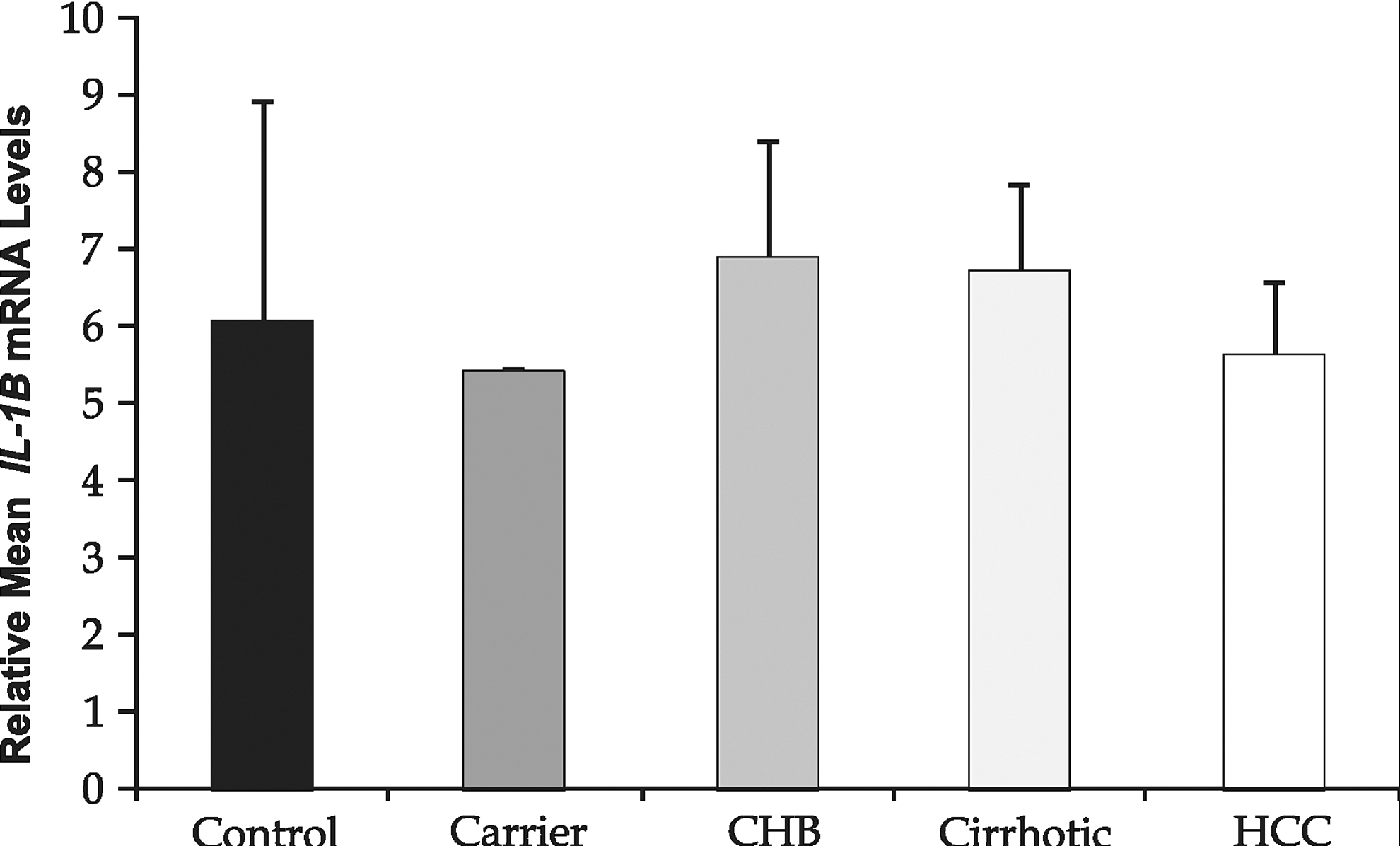
Reverse transcriptase–PCR analysis of IL-1B (normalized with β-actin) of some representative samples. Data are represented as mean±SD. Graph represents relative change in IL-1B mRNA expression.
Discussion
HBV accounts for the majority of HCC cases in the world, with the incidence of HbsAg carriers constituting nearly 2% to 11% of the Indian population (Manilal and others 2010). The HBV infection causes acute and chronic liver disease, leading to a wide array of clinical manifestations from inactive carrier to the carcinoma state. This differential response of the HBV infection in an individual, in part, is dependent upon various immunological factors, including the innate and adaptive immune response against the viral infection. Cytokines, the small proteineous entities, are considered to be the key mediators, regulating a myriad of inflammatory immune responses.
Out of these cytokines, IL-1, a key proinflammatory Th1 cytokine, has been documented to play an important role in HBV disease pathology. However, literature is scanty regarding the role of IL-1 in HBV pathogenesis in the Indian population. With this overview, the present case–control study was planned, which attempted to assess the association between the IL-1B(−511) and IL-1RN(VNTR) genotypes, haplotypes, and the mRNA and protein levels of this cytokine during the progression of hepatitis B infection from the asymptomatic carrier to the carcinoma state in the north Indian population. For this purpose, we initially conducted the polymorphism analysis of IL-1B(−511) and found that the CT genotype (54.90%) was the predominant form in the control population, similar to 59% and 46.7%, respectively, reported in the north Indians (Kaur and others 2007; Mittal and others 2007), 51% in the Polish (El-Omar and others 2000), 57%, 45%, and 46%, respectively, in the Japanese (Tanaka and others 2003; Ikehara and others 2006; Migita and and others 2007), and 43.3% in the Italian Caucasian population (Borzani and others 2010). However, this finding differed from the studies, where the CC genotype distribution, which varied from 48% to 50%, was observed to be maximal (Machado and others 2001; El-Omar and others 2003; Trajkov and others 2005). Also, in our study, we have observed that the variant (TT) genotype frequency of 54.24% was found to be highest in the HCC category, which differed from the studies, where the CT genotype was the predominant form (41.1%, 50.3%, and 51%, respectively) (Tanaka and others 2003; Chen and others 2005; Migita and others 2007). However, unlike our study, another study (Hirankarn and others 2006) reported the IL-1B-511 CC genotype to be significantly more in patients with HCC, with controls as reference.
The genotype distribution for the IL-1RN VNTR polymorphic forms showed that in the control population, the 1/1 genotype frequency of 52.94% was found to be the most prevalent one. This was observed to be in accordance with the studies conducted in Taiwanese (Chen and others 2005), in Americans (El-Omar and others 2003), in Portuguese (Machado and others 2001), in Germans (Glas and others 2004) and in the Japanese population (Kanemoto and others 2000; Ito and others 2007; Migita and others 2007). On the contrary, a study reported the predominance of 2/2 genotype (73.3%), among controls (Mittal and others 2007), while the frequency of 1/2 genotype (50%) was found to be highest in a study in the north Indian controls (Bid and others 2008). In our study, the incidence of the 1/2 genotype (50.85%) was maximal, compared to the other genotypes in subjects with HCC, consistent to that observed in another study (Chen and others 2005) in the same group (86%). However this varied from the study (Tanaka and others 2003) where the 1/1 genotype (88.8%) was the predominant form evident in the said group. Also, contrary to the present study, no significant difference in the IL-RN genotype frequencies was observed among Japanese patients afflicted with various stages of liver disease (Migita and others 2007).
Furthermore, on analyzing the OR for the association between the IL-1B(−511) genotypes and the carcinoma progression, it was observed that both the IL-1B-511(CT) and (TT) genotypes were found to be in no significant association with HBV-HCC development among controls and carriers. Similar to our results, a study (Zhang and others 2004) showed no difference in the IL-1B allele or the genotype frequencies between patients with HBV and the controls. This finding, however, differed from a study (Tanaka and others 2003), where the IL-1B-511(TT) genotype was found to be a significant risk factor (OR=2.19) for HCC development, among controls. Another study (Ito and others 2007) suggested that the IL-1B-511 heterozygotes were found to be associated with significantly reduced risk of gastric (OR=0.48) and colorectal (OR=0.34) carcinomas.
Further, in our study, the association analysis of the IL-1RN (VNTR) genotypes and the HCC risk, with 1/1 genotype as reference, showed that 1/2 genotype was found to be a potential risk factor (ORa=1.88 and ORa=4.10, respectively) for HCC development, among controls and carriers. Consistent to this, in another study (Machado and others 2001), carriers of IL-1B-511Tand IL-1RN*2 homozygotes were found to be risk factors for developing gastric carcinoma with OR=2.7; 95% CI=1.5–4.9 and OR=3.1; 95%CI=1.5–6.5, respectively. Similar findings were also observed in American patients with gastric cancer (El-Omar and others 2003). However, contrasting results were obtained in a study (Zhang and others 2004), where the carriage of the IL-1RN allele 2 was observed to have a significant protective role in HBV infection, among controls. On the other hand, no significant association was observed between 2/2 genotype and the liver disease progression in a Japanese study (Tanaka and others 2003), contrary to our study, where this genotype was found to be potentially associated with cirrhosis development (ORa=6.20 and ORa=5.04, respectively), among controls and carriers. Earlier, in a study done in Chinese patients with hepatitis B (Chan and others 2008), no significant association was found between the IL-1B and IL-RN polymorphisms and liver fibrosis.
On associating the IL-1 haplotypes with HCC risk, haplotypes 2 and 3 acted as potential protective factors for hepatitis and HCC development, among controls. However, haplotype 4 shared a significant negative association, only with the hepatitis group.
Besides, in this study, when analyzed in the control population, a weak LD was observed (D′=0.04; r 2=0.001) between the IL-1B-511 and IL-1RN (VNTR) loci, consistent to that observed in the Portuguese population (Machado and others 2001).
Activation of the host immune response in response to HBV infection results in increased production of proinflammatory cytokines such as IL-1 that determine the extent of disease chronicity. IL-1 is documented to be one such mediator of necroinflammation in the liver, regulating viral replication, disease activity, and prognosis. It not only regulates hepatocyte destruction but also elicits antiviral activity and stimulates the synthesis of acute-phase proteins (Sekiyama and others 1994). The hypothesis regarding the IL-1 genetic polymorphism and hepatocarcinogenesis is based on the assumption that the carriers of these genotypes are associated with increased levels of IL-1β in the liver in response to HBV infection and hepatocyte damage that may finally lead to the development of HCC. IL-1β is a proinflammatory cytokine as well as a tumor growth factor. There are several lines of evidence that support its role in tumor growth development. First, IL-1β can increase the production of prostaglandin E2 and hepatocyte growth factor (Bamba and others 1998). Second, IL-1β can induce angiogenesis, which is an important step in promoting tumor growth by either upregulating COX-2 or inducing nitric oxide and vascular endothelial growth factor (Chang and others 2005). Third, IL-1β can also attenuate interferon-induced antiviral activity and STAT1 activation in the liver (Tian and others 2000). On determination of the spontaneous IL-1B levels, significant (P<0.001) and steadily increased levels were evident in the subjects with HCC, as compared to the controls. This was consistent to an earlier study that also showed increased IL-1B levels in patients with fulminant hepatic failure with respect to the controls (Sekiyama and others 1994). Further, it was hypothesized (Zhang and others 2004) that high production of IL-1β may help increase the production of other cytokines such as IL-2, IL-6, and TNF-α and trigger the complex immunological processes to eliminate the virus. Further, elevated IL-1 also activates NF-κB, a transcription factor, documented to be often constitutively activated in development of HBV-HCC (Block and others 2003). This was further validated by the results obtained by us in a separate study, where a steady and significant increase (P<0.05) in NF-κBp65 expression was evident in subjects with chronic-active HBV and HCC, in comparison to the control population (data not shown).
To determine whether the IL-1B(−511C/T) genotypes influenced the levels of this cytokine, the IL-1B levels were compared among different genotypes. The levels were found to be markedly decreased (P<0.01) in controls with the TT genotype, w.r.t. those with the CT genotype. This finding further reconfirmed earlier documentation that the C allele accounted for higher IL-1B production (Hirankarn and others 2006). However, carriers and other diseased categories displayed a nonsignificant change in the levels, among the 3 genotypes.
No significant change was evident in the mean relative IL-1B mRNA levels, in any of the categories. However, a study done in liver biopsy samples of the patients with chronic hepatitis B (Shindo and others 1996) showed raised IL-1B transcript levels, in comparison to controls.
In conclusion, no potential association was evident between IL-1B(−511) genotypes and HCC risk, among controls and carriers. In case of IL-1RN(VNTR), 1/2 genotype acted as a potential risk factor for HBV disease chronicity and HBV-HCC development, among controls and carriers. Besides, 2/2 genotype was found to be a potential risk factor till cirrhosis development, among the same groups. However, IL-1 haplotypes 2 and 3 were found to be vital protective factors for hepatitis and subsequent HBV-HCC development, with controls as reference. Also, a significant and steady increase in proinflammatory IL-1B levels was evident with the disease progression to HCC, as compared to controls. Therefore, these findings suggest vital role of IL-1 in prediction of the HBV-induced disease transition in the Indian population, thereby aiding in modulation of the existing prognostic and therapeutic strategies. However, there is a need for larger comprehensive population-based studies to further substantiate these preliminary findings and to be conclusive.
Footnotes
Acknowledgment
The financial assistance by the Indian Council of Medical Research (ICMR) is highly acknowledged.
Author Disclosure Statement
No competing financial interests exist.