
Abstract
Besides functions of the interleukin-6 (IL-6)/gp130 cytokine family in immunology, IL-6 signaling has influence on memory processes. IL-6 acts on target cells via a membrane-bound IL-6 receptor (IL-6R) and subsequent association with the signal-transducing protein gp130. While gp130 is expressed on all cells in the body, IL-6R is expressed in only on few cells such as hepatocytes and some leukocytes. Cells lacking IL-6R were shown not to be responsive to the cytokine. Interestingly, a soluble form of the IL-6R in complex with IL-6 can stimulate cells that do not express the membrane-bound IL-6R. This signaling pathway has been called IL-6 trans-signaling. IL-6 trans-signaling can specifically be blocked by a soluble gp130 protein (sgp130Fc) without affecting IL-6 classic signaling via the membrane-bound IL-6R. Transgenic mice expressing sgp130Fc in the blood, but not in the central nervous system, were analyzed for hippocampus-dependent and independent memory, together with exploratory- and anxiety-related behavior. Transgenic animals did not show impaired hippocampus-dependent or independent learning and memory. However, compared to wild-type animals, they showed reduced exploratory behavior and an increased thermal pain threshold, indicating that these effects depend on IL-6 trans-signaling. These results bear important consequences for the therapeutic blockade of IL-6 activity in autoimmune diseases.
Introduction
B
There are at least 2 different IL-6-driven signaling pathways. The first is classical signaling, in which IL-6 acts via a membrane-bound IL-6 receptor (mIL-6R, CD126) and causes the recruitment of a dimer of gp130 coreceptor molecules (CD130), thereby initiating the activation of intracellular signaling via the JAK/STAT pathway. IL-6 classical signaling is mainly limited to some cell populations such as hepatocytes and some leukocytes, which express mIL-6R on their surface. An alternative mechanism is called trans-signaling, where a soluble form of the IL-6R (sIL-6R) forms an IL-6/sIL-6R complex with IL-6, capable to induce IL-6 responses via binding to gp130, enabling IL-6 to stimulate cells that lack an endogenous mIL-6R (Marz and others 1999). Although there is evidence that IL-6R mRNA is expressed in many neurons throughout the CNS (Schobitz and others 1993; Gadient and Otten 1994), most of them fail to respond to IL-6 alone (Marz and others 1998, 1999). This lack of responsiveness can be overcome by the addition of sIL-6R (Marz and others 1999). As the number of membrane-bound gp130 molecules is relatively constant on all cell types, including neuronal cells (Hibi and others 1990; Marz and others 1997; Islam and others 2009), the trans-signaling pathway seems to be of importance for IL-6 signaling in the CNS (Islam and others 2009). A soluble form of the gp130 receptor (sgp130) is found in body fluids, including the CNS (Marz and others 1997). sgp130 has been shown to act as a competitive inhibitor of IL-6 trans-signaling without affecting the classical signaling mediated via the membrane-bound IL-6R (Diamant and others 1997; Tanaka and others 2000; Jostock and others 2001; Jones and others 2011).
It has been somewhat contradictory whether IL-6 signaling has an influence on memory processes. On the one hand, there is evidence that overproduction of IL-6 is associated with cognitive decline in humans (Luterman and others 2000; Marsland and others 2006) and leads to suppressed hippocampal long-term potentiation (Bellinger and others 1995; Li and others 1997; Tancredi and others 2000) and impaired learning (Heyser and others 1997; Samuelsson and others 2006) in rodents. On the other hand, acute intranasal administration of IL-6 had a beneficial effect on sleep-dependent memory in healthy human subjects (Benedict and others 2009). However, the total absence of IL-6 signaling in IL-6 knockout mice led to impaired hippocampus-dependent (Baier and others 2009) and hippocampus-independent (Hryniewicz and others 2007; Baier and others 2009) learning.
In our group, we recently generated transgenic mice overexpressing a soluble gp130 protein dimerized by an Fc portion (sgp130Fc), in which transgene expression is driven by the hepatic PEPCK promoter (Rabe and others 2008). As the PEPCK promoter is only expressed after birth of the animals, it does not influence embryonic development, and the animals constitute a model in which IL-6 responses mediated by trans-signaling are specifically blocked, whereas IL-6 responses mediated via the membrane-bound IL-6R remain unaffected (Jostock and others 2001; Rabe and others 2008). The aim of the present study was to investigate the effects of this trans-signaling blockade on hippocampus-dependent and hippocampus-independent memory, in combination with the assessment of exploratory- and anxiety-related behavior of the animals.
Materials and Methods
Animals
All experimental procedures were conducted in accordance with the German Law of Animal Protection. The experiments were carried out on 10 male transgenic C57Bl/6N mice overexpressing sgp130 under the control of the PEPCK promoter [sgp130Fc; for details: Rabe and others (2008)] and 10 age-matched male C57Bl/6N wild-type (wt) mice. All mice were housed in plastic cages in a sound- and light-attenuated housing room under a constant light–dark cycle (light phase 08:30–20:30; 21±1°C) with ad libitum access to food and water. All behavioral experiments were conducted during the first hour of the light phase. Animal behavior was video-recorded on the hard disk of a personal computer using Noldus software EthoVision Basic (
Thermal threshold
Thermal pain threshold was assessed with the IITC Hot Plate Analgesia Meter (World Precision Instruments). All mice were placed on a 50°C warm hot plate surrounded by 20-cm-high Plexiglas walls. The latency to a nocifensive behavior (defined as shaking and/or licking the hindpaw or vocalizing) was measured, and the animals were immediately removed from the hot plate.
Elevated plus maze
Anxiety and exploratory behavior were assessed in the elevated plus maze (EPM). The EPM was made of dark gray plastic and was mounted 90 cm above the dark gray floor of a sound- and light-attenuated recording chamber. The center (5×5 cm) of the maze was connected to 4 arms (42.5×5 cm); the walls of the 2 closed arms were made of white opal plastic (39-cm high). Animals were put on the maze for 5 min, and the following parameters were analyzed: total time spent in the open and closed arms, total distance moved, and frequency of entering the center.
Open field
Activity monitoring was conducted for 5 min in a square-shaped (50×50 cm) open field (OF) that was made of dark gray plastic. The following parameters were assessed: total distance moved and the time spent in the border zone.
Morris water maze
The Morris water maze (MWM) is a classical test to investigate spatial and thus hippocampus-dependent memory consolidation (Morris 1984). The test was performed in a dark gray circular pool (diameter 120 cm) filled with water at a temperature of 24±0.5°C with 4 landmarks (black geometrical symbols) arranged around the water maze. For analysis, the maze was virtually divided into 4 quadrants. In the target quadrant, a transparent escape platform (diameter 10 cm) was placed 1 cm beneath the water surface. With an interval of one week, 2 independent MWM experiments with different target locations were performed for all animals. Each experiment consisted of 3 subsequent training days with 3 training sessions and 1 test (reversal) day with 2 sessions. When animals did not find the platform within 90 s during the training sessions, they were placed on it for additional 10 s. On the probe day, the platform was removed, and 2 test trials were run with a duration of 60 s and with starting points at 2 randomly chosen quadrants. The following parameters were determined for each session: escape latency until the mice reached the platform or its position (in probe trials), mean distance to the platform, time spent in the border zone as percentage of the recording time, and total distance covered by the mice.
Novel object recognition memory
A well-established test to investigate hippocampus-independent memory performance is the novel object recognition memory (NORM) test (Ennaceur and others 1997). This test utilizes the natural tendency of rodents to explore novel things more than familiar ones. In our study, an experiment comprised 2 sessions of each 300 s performed in the animals' home cage: During the first session (learning session), 2 identical objects were presented to the animal, and 1 h later (retrieval session), one of the familiar objects was replaced by a novel one. The following parameters were assessed for both sessions: cage exploration as percentage of the total time. For the retrieval session, a recognition index was calculated for each animal: (time novel object)×100/[(time novel object)+(time familiar object)].
Statistical analyses
All statistical analyses were performed with the standard software package GraphPad Prism 4. The number of animals per group was n=10. Either unpaired t-tests were run to detect genotype (wt versus sgp130Fc) differences, or 2-factor-repeated-measurement analysis of variances with the factor genotypes (wt versus sgp130Fc) and day of the experiment. When appropriate, post hoc Bonnferroni t-tests were calculated. The level of significance was set at P<0.05.
Results
Levels of sgp130Fc in the periphery and in the CNS
The sgp130Fc transgenic mice have been shown to express microgram amounts of the sgp130Fc protein in the plasma (Rabe and others 2008). We now wanted to assess whether sgp130Fc is also found in the CNS. As shown in Fig. 1, we compared the levels of the sgp130Fc protein in the serum (Fig. 1A) and in brain homogenates after perfusion (Fig. 1B). In the serum of the sgp130Fc transgenic mice, the levels ranged between 20,000 and 30,000 ng/mL, whereas in the brain of the same animals, only background levels of up to 30 ng/mL were detected. We conclude from these data that the sgp130Fc protein does not cross the blood–brain barrier under unchallenged conditions, and that therefore the sgp130Fc concentrations in the serum are responsible for the effects observed in the transgenic mice, in which IL-6 trans-signaling was blocked.
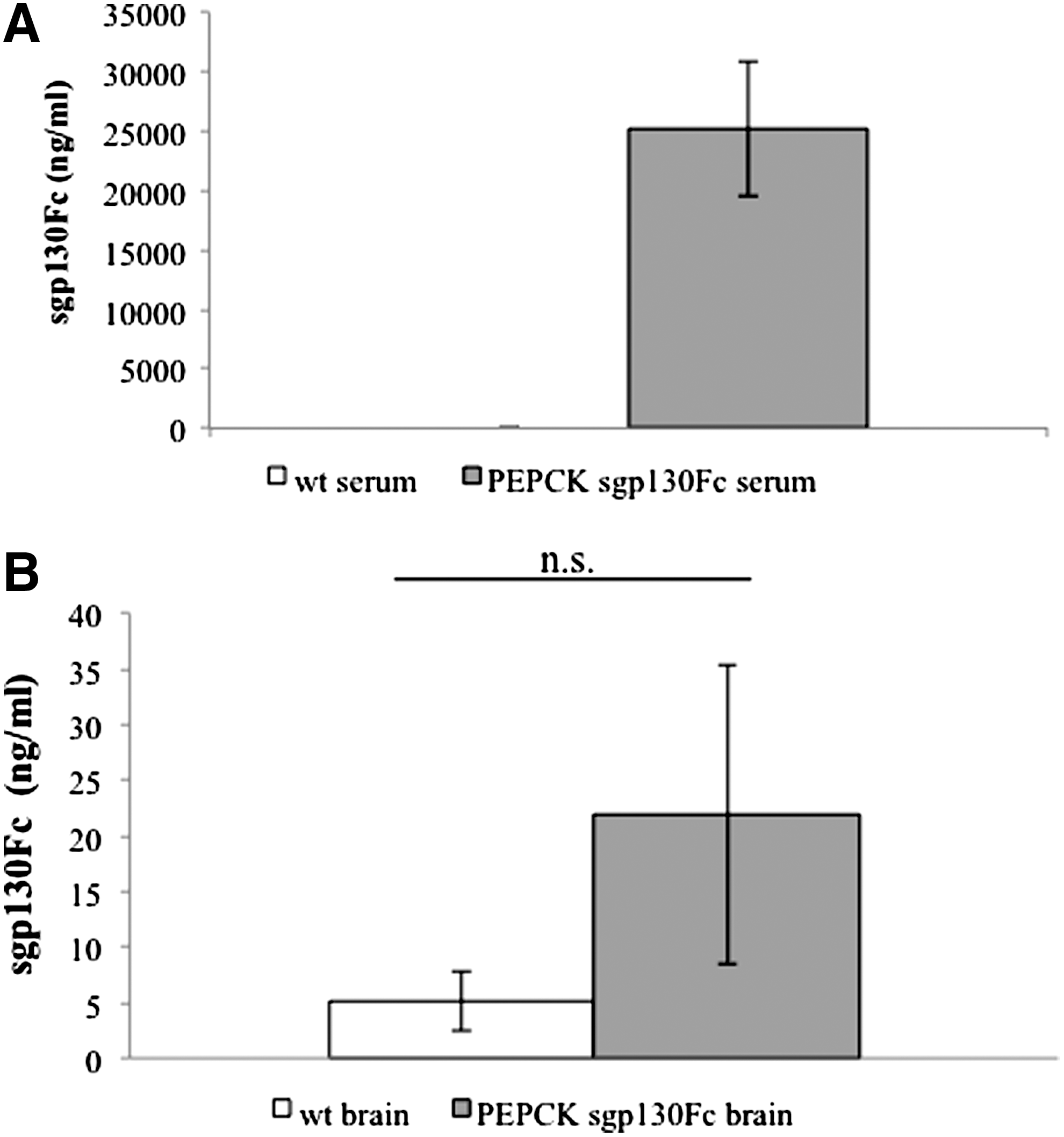
Levels of sgp130Fc protein in serum and brain homogenates of sgp130Fc transgenic mice.
Hippocampus-dependent memory
The results from MWM experiments are shown in Fig. 2. Typically, the primary outcome measure in this test is the latency, which the animals needed to reach the platform (training sessions) or the target quadrant (probe session). In a first analysis, the large variability in the results of the sgp130Fc animals led us to examine more closely the results of the individual animals. This revealed that while all wt mice (black circle) and 7 of the 10 sgp130Fc mice (black squares) learned the task (Fig. 2A), 3 sgp130Fc mice (empty squares) pursued a different strategy and kept to the border of the water maze (Fig. 2B) without trying to reach the platform (Fig. 2C). There were no differences between the wt mice and the learning sgp130 mice, whereas the nonlearning sgp130 mice differed significantly from both other groups.
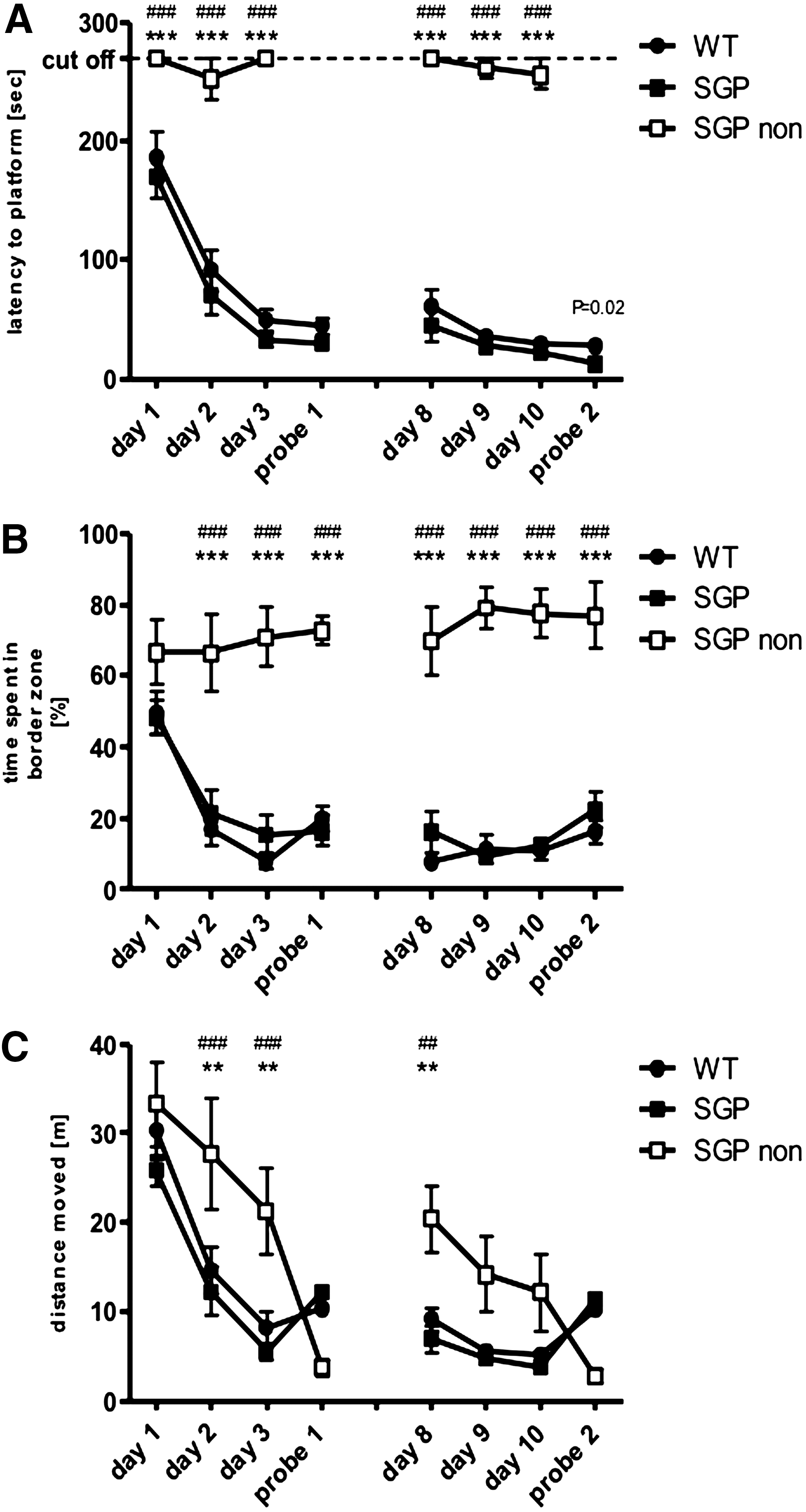
Blockade of IL-6 trans-signaling in the MWM test. Results from the MWM displayed as mean values±SEM; circles for WT animals, boxes for sgp130Fc mice. sgp130Fc showing no initial learning behavior are plotted as empty boxes. The experiment consisted of 2 phases: a training phase, which lasted for 3 days, and a probe trial. We performed 2 experiments (first experiment on left hand side; second experiment on right hand side) with an interval of 1 week. Shown are
Hippocampus-independent memory
The results from the novel object recognition test (NORM) are displayed in Fig. 3. There was no significant difference in learning between the 2 groups as measured by the recognition index (Fig. 3A, t-test, P>0.2). However, sgp130Fc mice spent significantly less time in exploring both objects in the learning (Fig. 3B; unpaired t-test, P=0.0033) as well as in the retrieval session (Fig. 3C, unpaired t-test, P=0.0255, n=10). Interestingly, the nonlearners from the MWM experiment (Fig. 2) did not differ from the other sgp130Fc mice in their exploring behavior or recognition index.
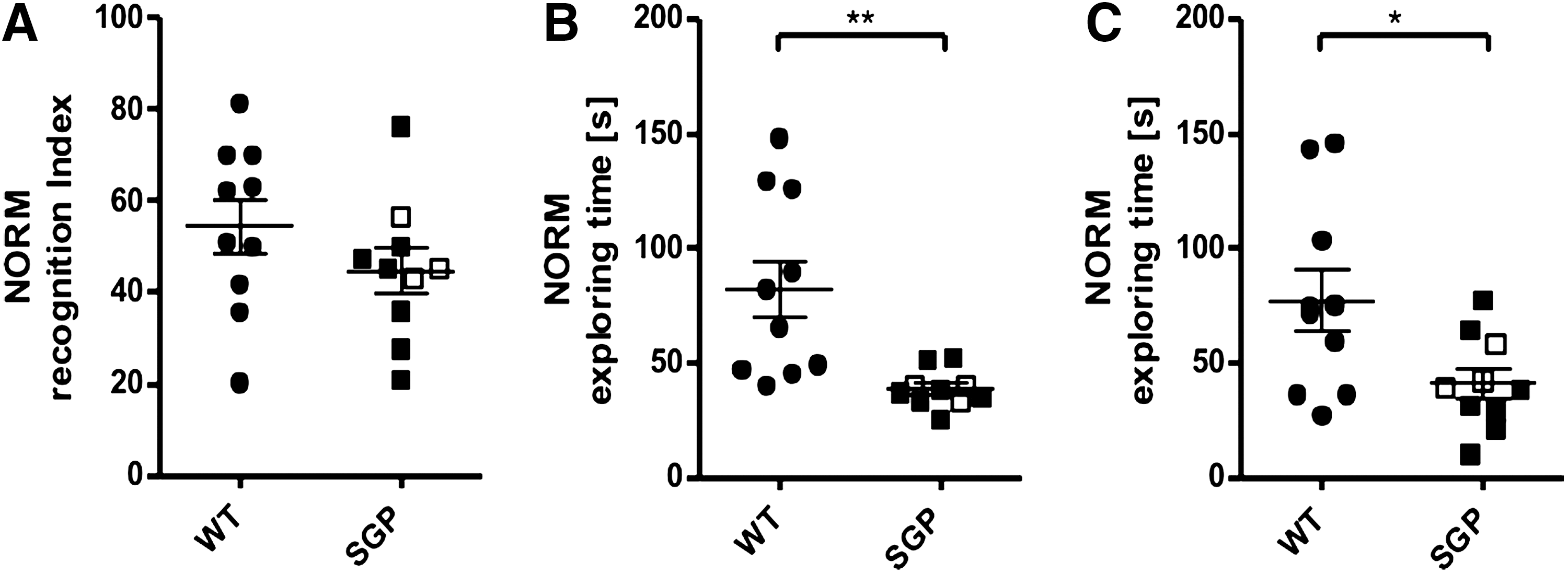
IL-6 trans-signaling in novel object recognition. Results from the novel object recognition test (NORM) as individual measures, mean values±SEM; circles for WT animals, boxes for sgp130Fc mice. sgp130Fc mice not learning in the MWM are indicated by empty boxes.
Anxiety behavior
To control for the influence of stress- and anxiety-related behavior, the OF and EPM were performed. Although sgp130Fc mice seemed more agitated than wt animals during handling, this was not reflected by increased locomotive activity as measured by total distance moved in any of the performed behavioral experiments (Fig. 4). Furthermore, sgp130Fc mice did not differ significantly from wt animals in the OF concerning the time amounts spent in the border zones (Fig. 4A, B). Likewise, analysis of the behavior in the EPM did not reveal any significant differences between wt and sgp130Fc mice (Fig. 4C–F). Of particular interest is that the MWM nonlearners of the sgp130Fc group did not show any extreme anxiety-related behavior in any of the experiments (Fig. 2).
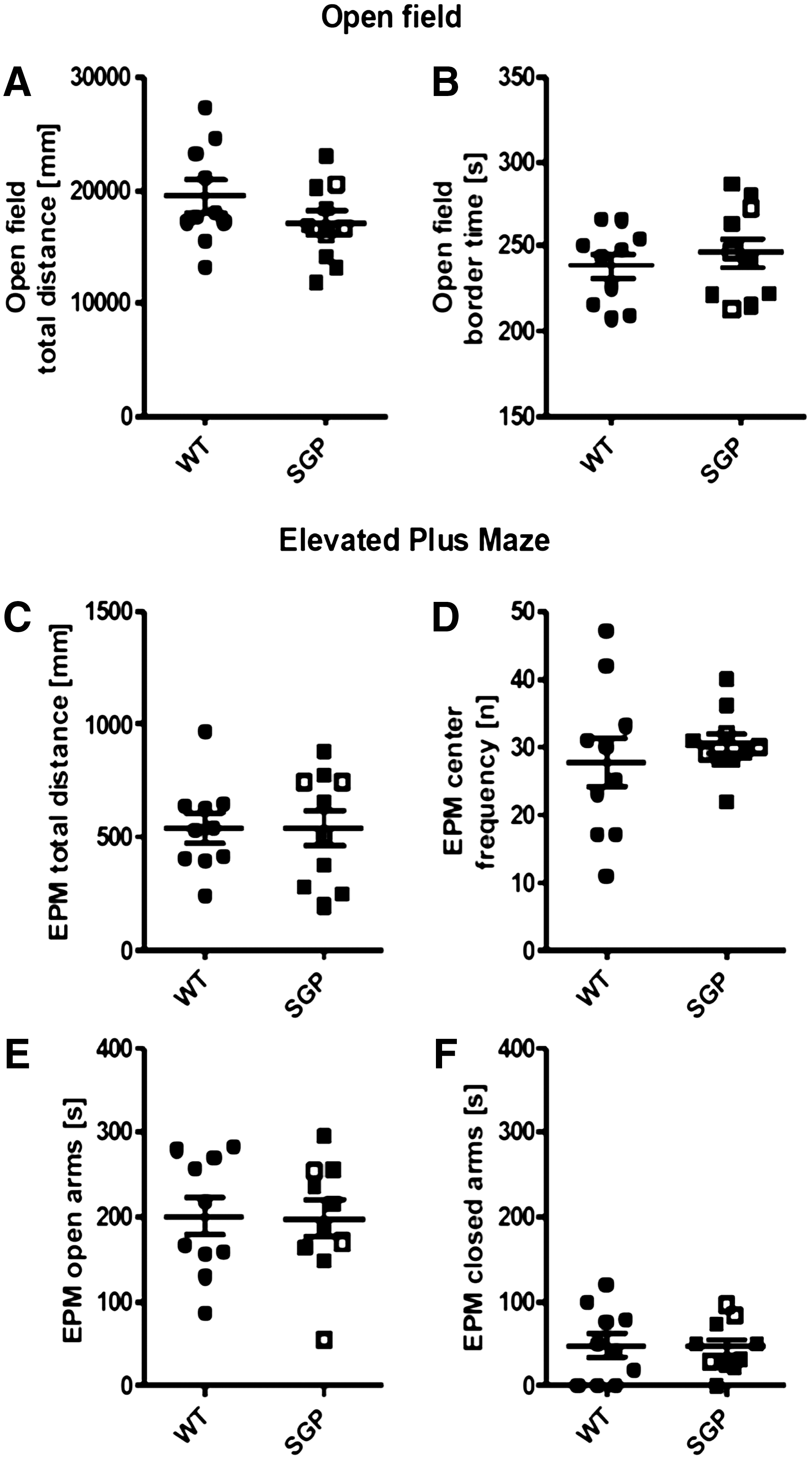
Results from testing anxiety behavior in the open field.
We next assessed the thermal pain threshold using a hot-plate analgesia meter instrument. Mice were placed on a hot plate, and the latency to a nocifensive behavior was measured. As shown in Fig. 5, in comparison to wt mice, sgp130Fc animals displayed a slight, but statistically significant, increased pain threshold (Fig. 5).
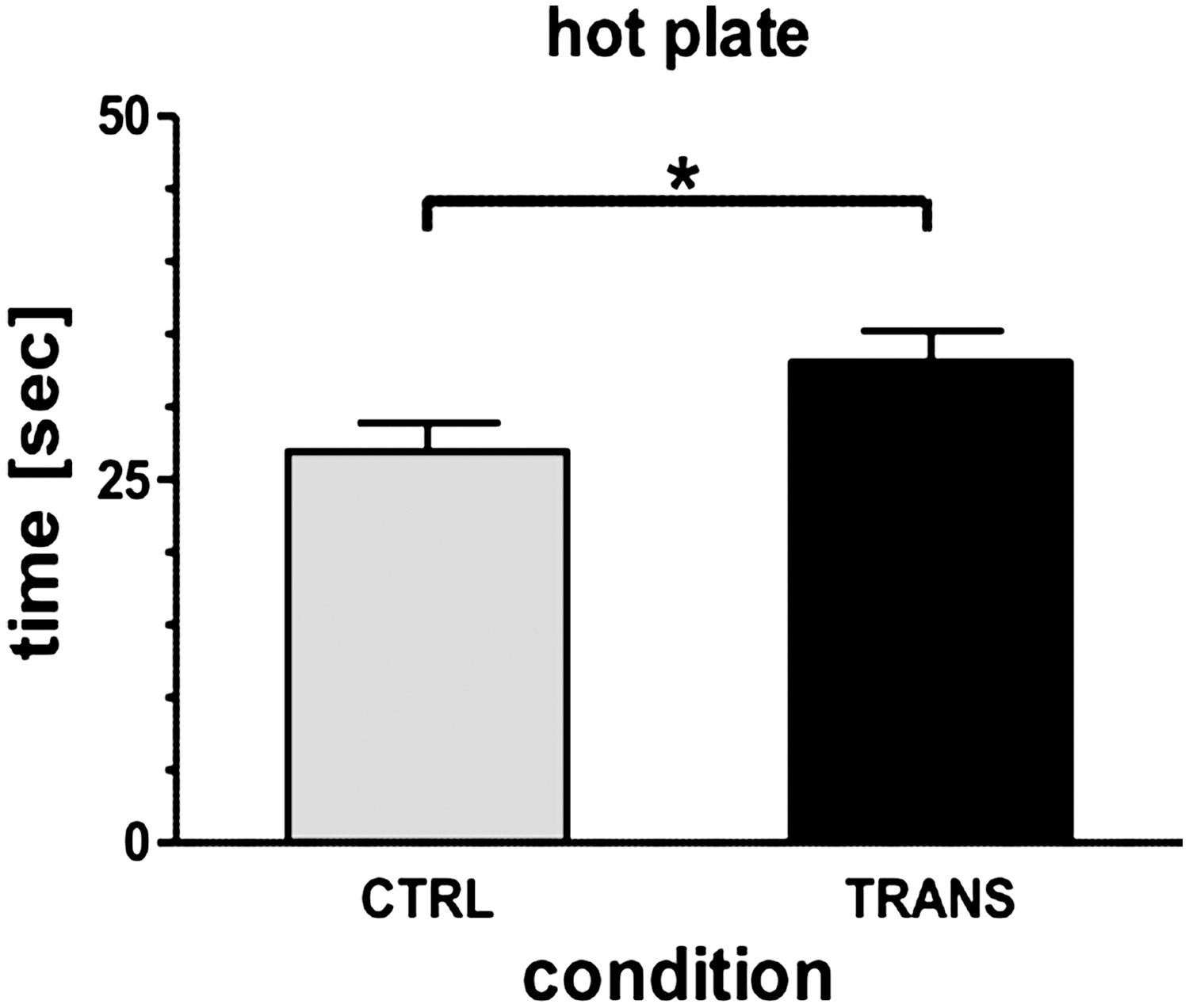
Decreased heat sensitivity in sgp130Fc transgenic mice. Thermal pain threshold was assessed by placing mice on a 50°C hot plate surrounded by Plexiglas and recording the time to nocifensive behavior. *P<0.05. CTRL, control mice; Trans, sgp130Fc transgenic mice.
Discussion
In our study, we found that the investigated transgenic animals, overexpressing sgp130Fc, did not show generally impaired hippocampus-dependent or hippocampus-independent learning and memory as assessed by the MWM test and novel object recognition, respectively. However, there are some general differences in their behavior in comparison to wt animals of the same genetic background. First, there are some unspecific changes, as reduced body weight and exploratory behavior. Secondly, sgp130Fc mice show an increased thermal pain threshold, and last but not least, 3 out of 10 sgp130Fcmice showed despair behavior in the MWM, without any differences in anxiety, exploratory, or learning behavior in the other tests.
To interpret these differences, it is of importance to understand that in the investigated sgp130Fc animals, the expression of the sgp130Fc transgene was driven by the liver-specific PEPCK promoter (Rabe and others 2008). As PEPCK is only expressed in the liver, sgp130Fc is secreted into the blood and, apparently, does not have access to the CNS (Fig. 1). Concerning functions outside the CNS, sgp130Fc transgenic mice hence can serve as a model to evaluate the physiologic consequences of a therapeutic life-long blockade of IL-6 trans-signaling (Lemmers and others 2009; Jones and others 2011), but not IL-6 classical signaling (Jostock and others 2001). In addition, as the PEPCK promoter is only activated after birth of the animals (Ballard and Hanson 1967), the overexpression of sgp130Fc begins postpartum, and it thus can be assumed that embryologic development is unaffected. However, although trans-signaling is supposed to be the main IL-6 pathway in the CNS, and there is evidence that the gp130Fc fusion protein might pass the blood–brain barrier under inflammatory conditions (Linker and others 2008), it is likely that the effects seen in this study in the investigated sgp130Fc animals are not mediated by central nervous mechanisms.
Animals with a complete knockout of IL-6 signaling showed impaired hippocampus-dependent (Sparkman and others 2006; Baier and others 2009) and hippocampus-independent learning and memory (Hryniewicz and others 2007; Baier and others 2009), indicating that IL-6 signaling in general influenced these processes. However, it remained unclear whether IL-6 classic signaling or IL-6 trans-signaling was involved. Our study using peripheral blockade of trans-signaling by sgp130Fc showed that IL-6 trans-signaling in the periphery did not contribute to these processes. It will be interesting to study whether blockade of IL-6 trans-signaling specifically in the CNS will lead to similar effects as seen in the IL-6-deficient mice (Sparkman and others 2006; Baier and others 2009).
Three mice behaved abnormally in the test on hippocampus-dependent memory (Fig. 2). Although we do not have an explanation for this behavior, it is not uncommon that in transgenic animals, the observed phenotype is not 100% penetrant (Peters and others 1997; Maione and others 1998; Suthaus and others 2012). Interestingly, these 3 animals did not behave unexpectedly in the test on hippocampus-independent memory (Fig. 3) and in the test on anxiety behavior (Fig. 4).
Interestingly, profound changes in anxiety behavior (Armario and others 1998; Butterweck and others 2003) and reduced tolerance to psychological and immunological stressors as detected in IL-6 knockout mice (Bluthe and others 2000; Butterweck and others 2003; Swiergiel and Dunn 2006) were not found in the sgp130Fc mice. The increased thermal pain threshold, on the other hand, is in line with the findings that endogenous IL-6 is required for thermal hyperalgesia (Murphy and others 1999), involved in the modulation of muscular pain (Manjavachi and others 2010), and that neutralization of IL-6/sIL-6R is particularly successful in producing antinociception (Boettger and others 2010). In addition, a recent study achieved that ablation of gp130 in the nociceptive neuron. These mice showed significantly reduced levels of inflammation- and tumor-induced pain (Andratsch and others 2009). These effects apparently do not require central nervous mechanisms. This present study now demonstrates that this gp130 effect relies on IL-6 trans-signaling. Reduced exploratory behavior, as seen in both, sgp130Fc mice (this study) and IL-6 knockout mice (Butterweck and others 2003; Baier and others 2009), might be an IL-6 trans-signaling-related effect. Again, studies with animals with blockade of IL-6 trans-signaling in the CNS might help to clarify these issues. Such mice are currently being generated in our laboratory.
In summary, the animals investigated in this study overexpress the sgp130Fc protein exclusively outside the CNS and show behavioral changes that all can be explained with peripheral mechanisms. Therefore, the lack of impact on hippocampus-dependent and hippocampus-independent learning does not necessarily contradict the hypothesis of IL-6 trans-signaling being involved in these processes (Benedict and others 2009), but rather suggests that no efficient blockade of this pathway can be achieved in this animal experimental approach. This is an important finding, since it suggests that long-term sgp130Fc treatment of patients suffering from chronic inflammatory diseases will likely not lead to behavioral impairments due to peripheral blockade of IL-6 trans-signaling. Neutralization of the IL-6R by the monoclonal antibody tocilizumab is a therapy, which has been approved in more than 100 countries for the treatment of rheumatoid arthritis and other inflammatory diseases (Tanaka and others 2012). It remains to be shown whether trans-signaling is involved in the behavioral changes seen in IL-6-deficient animals (Baier and others 2009). Studies with animals in which the expression of the sgp130Fc transgene is under the transcriptional control of a promoter specific for the CNS are therefore warranted to further investigate the impact of IL-6 trans-signaling on learning and memory. Transgenic mice that express the sgp130Fc transgene under the control of the GFAP promoter have been generated in our laboratory and are currently under investigation. The comparison of the behavior of mice with peripheral and central blockade of IL-6 trans-signaling will help to understand the contribution of this important signaling pathway.
Footnotes
Acknowledgments
The research was supported by the Sonderforschungsbereich SFB 654 Plasticity and Sleep (projects B6, C5, and C8) by the Deutsche Forschungsgemeinschaft (DFG) and by the Cluster of Excellence Inflammation at Interfaces. We thank Mona Below for excellent technical support.
Author Disclosure Statement
No competing financial interests exist.