
Abstract
Endurance exercise can cause immunosuppression and increase the risk of upper respiratory illness. The present study examined changes in the secretion of T helper (Th) cell cytokines after endurance exercise. Ten highly trained road cyclists [mean±SEM: age 24.2±1.7 years; height 1.82±0.02 m; body mass 73.8±2.0 kg; peak oxygen uptake 65.9±2.3 mL/(kg•min)] performed 2 h of cycling exercise at 90% of the second ventilatory threshold. Peripheral blood mononuclear cells were isolated and stimulated with phytohemagglutinin. Plasma cortisol concentrations and the concentration of Th1/Th2/Th17 cell cytokines were examined. Data were analyzed using both traditional statistics and magnitude-based inferences. Results revealed a significant decrease in plasma cortisol at 4–24 h postexercise compared with pre-exercise values. Qualitative analysis revealed postexercise changes in concentrations of plasma cortisol, IL-2, TNF, IL-4, IL-6, IL-10, and IL-17A compared with pre-exercise values. A Th1/Th2 shift was evident immediately postexercise. Furthermore, for multiple cytokines, including IL-2 and TNF (Th1), IL-6 and IL-10 (Th2), and IL-17 (Th17), no meaningful change in concentration occurred until more than 4 h postexercise, highlighting the duration of exercise-induced changes in immune function. These results demonstrate the importance of considering “clinically” significant versus statistically significant changes in immune cell function after exercise.
Introduction
P
Stress-related changes in immune system function are typically mediated through activation of the sympathetic nervous system (SNS) and the hypothalamic-pituitary-adrenal axis. Activation of the SNS induces catecholamine release, with a delayed release of glucocorticoids. These stress hormones alter immune function after acute exercise and during periods of heavy training (Pedersen and others 1998), in response to acute and chronic psychological stress (Cohen and others 1991), and after surgery (Rosenberger and others 2009). The effects of exercise-induced stress hormones on T helper (Th) cell function during and after prolonged heavy-intensity exercise have not been fully elucidated.
Glucocorticoids and catecholamines influence Th cell cytokine release, principally by stimulating glucocorticoid and β2 adrenergic receptors, respectively (Calcagni and Elenkov 2006). In turn, these stress hormones influence the activity of cellular (Th1), humoral (Th2), and mucosal (Th17) branches of the immune response (Elenkov 2004; Calcagni and Elenkov 2006). Specifically, cortisol, adrenaline, and noradrenaline may suppress cytokine release from antigen-presenting cells and Th1 cells, while upregulating cytokine production from Th2 cells, thereby causing a shift toward Th2 cytokine production (Steensberg and others 2001; Lancaster and others 2004). The balance between Th1 and Th2 immunological responses is very important in maintaining optimum immune health. The Th1 pathway primarily acts against intracellular pathogens, particularly viruses and bacteria, while the Th2 pathway is believed to protect against extracellular pathogens such as parasites (Kidd 2003). Previous research has extensively examined changes in the balance of Th1 and Th2 cytokines in plasma in the first 1−2 h after exercise (Suzuki and others 2002). By contrast, there is less research published concerning changes in Th cytokine secretion (including Th17 cytokines) during recovery from prolonged heavy exercise. While these changes are transient, it may have implications related to potential immune suppression after prolonged heavy-intensity exercise (Nieman 2000).
Exercise-induced glucocorticoid release occurs during prolonged exercise (Pedersen and others 1998). An increase in glucocorticoid concentration after exercise may influence Th cytokine secretion differently from exercise of moderate duration and intensity because of the large effect of changes in cortisol on immune function (Pedersen and others 1998; Walsh and others 2011b). Th17 cells play a key role in mucosal host defense of the upper respiratory and gastrointestinal tracts against a myriad of airborne antigens (Yao and others 1995; Liang and others 2006; Korn and others 2009). Functioning as part of the “first line of defense” in the mucosa, Th17 cytokines have been linked to increased polymeric immunoglobulin receptor expression, saliva SIgA, and human β-defensin 2 release (Brandtzaeg and others 1999; Johansen and others 1999; Kao and others 2004; Jaffar and others 2009). These results demonstrate that Th17 cytokine pathways are influential in the innate immune response in the mucosa. However, there is little evidence available about changes in Th17 cytokines after exercise.
The aim of the present study was to examine the effect of prolonged heavy exercise on plasma cortisol concentration and Th1, Th2, and Th17 cytokine release. We hypothesized that the exercise-induced stress would increase plasma cortisol concentration after exercise, resulting in the release of IL-17 and a shift from secretion of Th1 to Th2 cytokines.
Materials and Methods
Subjects
Ten well-trained male cyclists volunteered to participate in the present study [mean±SEM: age 24.2±1.7 years; height 1.82±0.02 m; body mass 73.8±2.0 kg; peak oxygen uptake 65.9±2.3 mL/(kg•min)]. Subjects were excluded if they exhibited any cardiovascular, metabolic, neurological, immunological, or autoimmune disorders. The cyclists provided written informed consent for participation in the study, which was approved by the Bond University Human Research Ethics Committee.
Experimental design
The cyclists visited the laboratory on 3 occasions; for a screening/familiarization session, a maximal exercise test, and a constant load test with blood sampling before and after exercise. Peak oxygen consumption (
The cyclists refrained from exercise for 48 h before the maximal exercise test and experimental trial and avoided consumption of alcohol or caffeine in the previous 24 h, during the exercise, and postexercise sample period. Pre-exercise blood samples were collected from the cyclists after an overnight fast. The cyclists then consumed a standardized breakfast (2,000 kJ) of cereal, milk, and juice and commenced the exercise trial at 0800 hrs. The cyclists performed exercise for 2 h at 90% VT2 (Monark Ergomedic 828 E). Cardiac rate and rhythm were monitored continuously throughout the exercise, and cardiac rate was recorded each minute. Pulmonary gas exchange was sampled for 4 min at 20-min intervals to ensure that the cyclists were exercising at the work rate associated with 90% VT2 and that no drift in oxygen uptake occurred. The cyclists were allowed to drink water ad libitum during exercise. The cyclists were provided with a standardized meal (4,500 kJ) of sandwiches, and a milkshake at 2.5 h postexercise.
Blood collection
Venous blood samples were collected pre-exercise, immediately postexercise, and 2, 4, 6, 8, and 24 h postexercise. Blood was collected into tubes containing K3EDTA (Becton Dickinson). One tube was immediately centrifuged at 1,000×g for 15 min. Plasma was stored in aliquots at −80°C. The remaining blood sample was used for the cytokine stimulation assay.
Plasma cortisol analysis
Plasma cortisol was analyzed by ultra-high-performance liquid chromatography tandem mass spectrometry (UHPLC MS/MS) described elsewhere (McWhinney and others 2010). Briefly, plasma samples were treated with 1 M HCl at room temperature to displace cortisol and cortisone from binding proteins. To each sample, the multianalyte internal standard (d4-dexamethasone, d2-11-deoxycortisol, and d2-cortisone) and H2O were added before loading onto activated Oasis HLB 1 cc (30 mg) solid-phase extraction cartridges (Waters). Samples were eluted with 100% ethyl acetate, dried at 50°C, and reconstituted in 45% methanol with 2 mM ammonium acetate and 0.1% formic acid. The flow rate of the UHPLC analysis was 0.4 mL/min, sample injection volume 20 μL, and column temperature 50°C. The tandem mass spectrometer was operated in positive mode electron spray ionization. The total analytical run time on the UHPLC MS/MS was 3 min.
Flow cytometry
Peripheral blood mononuclear cells were isolated using Ficoll-Paque PLUS (GE Healthcare Life Sciences) and then counted (Countess Automated Cell Counter; Life Technologies). Cells were cultured at a concentration of 1×106 cells/mL and stimulated with 1 μg/mL of the mitogen phytohemagglutinin (PHA) for 72 h. After incubation, the supernatant was removed and stored at −80°C for subsequent analysis. Th1, Th2, and Th17 cytokine concentrations were quantified using a commercially available cytometric bead array kit (BD Pharmingen) to measure the concentrations of IL-2, IL-4, IL-6, IL-10, TNF, INF-γ, and IL-17A from supernatant of stimulated samples. In the absence of stimulation with PHA, the concentrations of these cytokines were not detected.
Statistics
Before statistical analysis, all data were tested to determine if they were normally distributed. Data that were normally distributed (Fig. 1) were analyzed using one-way ANOVA with repeated measures to determine any change over time, with a Bonferroni correction applied post hoc when significance was evident. These data are presented as mean±SEM. Data that were not normally distributed (Table 3) were analyzed using the nonparametric Friedman's ANOVA and Wilcoxon signed-rank test. These data are presented as median±interquartile range. The Bonferroni correction was used for all multiple comparisons, with significance set at P<0.008 (P<0.05 divided by the number of time points). All statistical analyses were performed using SPSS software version 19.0 (SPSS Inc.). Data were further analyzed for clinical significance using magnitude-based inferences (Hopkins and others 2009). All qualitative analyses based on effect size were conducted using a modified statistical spreadsheet (available at
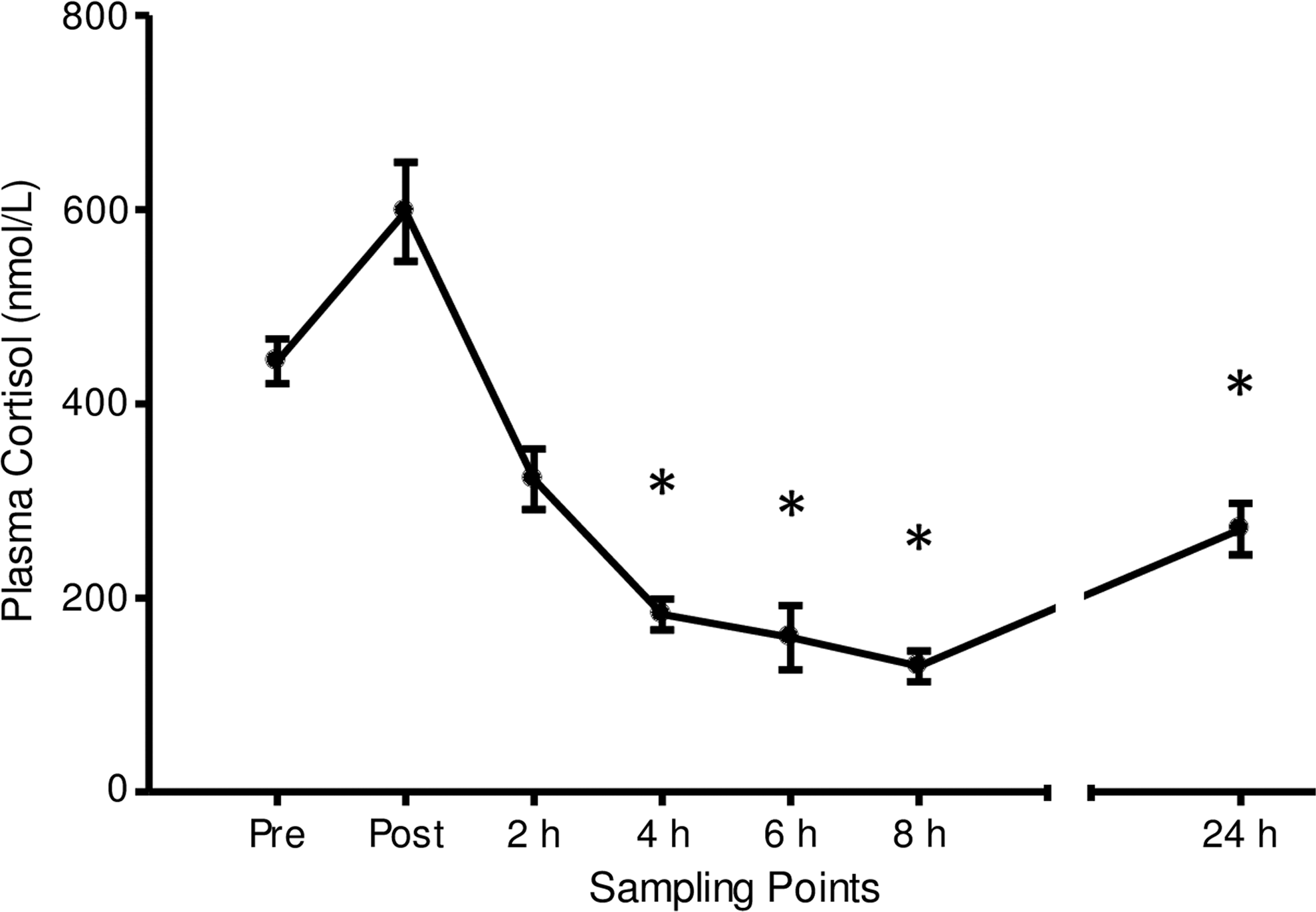
Plasma cortisol concentration before and after prolonged heavy-intensity cycling. Data represent mean±SEM. *Significantly change compared with “Pre-exercise” (P<0.05).
Results
Physical characteristics and exercise values
Physical characteristics of the cyclists are presented in Table 1. Mean heart rate for the 2 h exercise bout was 161±3 beats/min, and mean exercise intensity associated with 90% VT2 occurred at 75.1±0.9% of
Data are mean±standard error of the mean.
Plasma cortisol
Figure 1 displays plasma cortisol concentration after prolonged heavy-intensity exercise. There was no statistically significant change immediately or 2 h postexercise. Plasma cortisol concentration significantly decreased 4 h postexercise (P<0.001), when compared with the resting pre-exercise concentration, and remained significantly decreased at 6 h postexercise (P<0.01), 8 h postexercise (P<0.001), and 24 h postexercise (P<0.01).
Magnitude-based inferences revealed that compared with pre-exercise, plasma cortisol concentration was likely higher immediately postexercise. The chances that the true difference in cortisol was higher/trivial/lower were 94/5/1%. Cortisol was very likely lower at 2 h postexercise (1/2/97%). Furthermore, compared with pre-exercise, cortisol was almost certainly lower at 4 h (0/0/100%), 6 h (0/0/100%), and 8 h (0/0/100%). At 24 h postexercise, cortisol was likely lower (4/12/84%) than pre-exercise values (Table 2).
Data presented as percentage difference compared with the pre-exercise concentration. CL, confidence level.
Th cell cytokines
There were no statistically significant changes in PHA-stimulated production of the Th 1 cytokines IL-2, TNF, and IFN-γ; Th 2 cytokines IL-4 and IL-10; and Th 17 cytokine IL-17A after exercise (Table 3). A significant time effect was observed for IL-6; however, post hoc analysis revealed no significant change at individual time points after exercise compared with the pre-exercise IL-6 concentration.
Th1 cytokines: IL-2, TNF, IFN-γ; Th2 cytokines: IL-4, IL-6, IL-10; Th17 cytokine: IL-17A. Data represent median (1st and 3rd quartile).
Table 4 displays changes in Th1 cytokine concentrations. Compared with pre-exercise values, the concentration of IL-2 was likely higher than pre-exercise at 4 h (85/15/0%) and 8 h (90/10/0%) postexercise. The concentration of TNF was also likely higher than pre-exercise values at 4 h postexercise (82/17/1%). Changes in IFN-γ concentration were unclear at every time point postexercise.
Data presented as percentage difference compared with the pre-exercise concentration.
Compared with pre-exercise, the concentration of IL-4 was likely higher immediately postexercise (79/18/3%), while the difference in IL-4 during the remaining recovery period was unclear (Table 5). Compared with pre-exercise, the IL-6 concentration was possibly higher at 4 h (68/29/3%), possibly lower at 6 h (4/42/54%), and likely higher at 8 h postexercise (94/6/0%). Compared with pre-exercise, IL-10 concentration was likely higher at 4 h postexercise (77/23/0%).
Data presented as percentage difference compared with the pre-exercise concentration.
IL-17A concentration compared with pre-exercise was possibly higher than pre-exercise at 8 h postexercise (73/25/2%) (Table 6).
Data presented as percentage difference compared with the pre-exercise concentration.
Discussion
This is the first study to analyze changes in Th1, Th2, and Th17 cytokine production during recovery from exercise. It is established that intensity and duration influence the cortisol response to exercise. Previously, cycle exercise of comparable intensity (∼75%
Plasma cortisol concentration decreased below pre-exercise levels from 4 to 24 h postexercise. While the initial decrease (which occurred early in the afternoon) may be explained by diurnal variation (Dhabhar and others 1994), plasma cortisol concentration remained below resting values at 24 h postexercise. This low plasma cortisol concentration may be clinically important when considering the repetitive nature of endurance training day after day. This result may indicate that the cyclists were well rested after the exercise bout, and would have been able to respond positively to an additional bout of exercise on the second morning.
Using traditional data analysis, there were no statistically significant changes in Th cell cytokine secretion at any time point during the postexercise period in the present study (Table 3). However, analysis using magnitude-based inferences suggested a multitude of changes after exercise. Compared with pre-exercise, IL-2 (Th1) production was likely higher at 4 h and again at 8 h postexercise (Table 4), while TNF (Th1) production was likely higher at 4 h postexercise (Table 4). IL-4 (Th2) production was likely higher immediately postexercise (Table 5), while IL-6 (Th2) production was possibly higher at 4 h, possibly lower at 6 h, and likely higher at 8 h postexercise (Table 5). IL-10 (Th2) production was likely higher at 4 h postexercise (Table 5), while IL-17A (Th17) production was possibly higher at 8 h postexercise (Table 6).
Previously, intracellular cytokines IL-2 (Th1), IFN-γ (Th1), and IL-4 (Th2) produced by CD4+ T cells were used to define Th cell types and their functional changes. Using this method, a Th1-to-Th2 shift was described after exercise and hormone infusion (Steensberg and others 2001; Elenkov 2004, 2008; Lancaster and others 2004). This shift occurs when IL-2- and/or IFN-γ-producing Th cells (Th1) decrease in cell concentration, while IL-4-producing Th cells (Th2) remain unchanged. In the present study, no change in IL-2 and IFN-γ occurred immediately postexercise (Table 4), while IL-4 concentration increased at the same time point (Table 5). Hence, a shift toward Th2 was observed immediately postexercise.
Although single intracellular cytokine analysis of Th cells provides a concise explanation of the Th response to exercise, it neglects to consider the magnitude of cytokines involved in the Th cell network. Cytokines are not solely secreted from Th cells, but include cytotoxic T cells, natural killer cells, dendritic cells, macrophages, and even skeletal muscle (Zaldivar and others 2006; Pedersen and Febbraio 2008). In the present study, cytokine secretion both increased and decreased occurred immediately after exercise (IL-4), and up to 8 hours postexercise (IL-2, IL-6, and IL-17A). This presents an intricate pattern of changes to consider. In particular, at 4 h postexercise, the Th1 cytokines IL-2 and TNF, and the Th2 cytokines IL-6 and IL-10 were all increased. This concomitant increase and balancing of Th1 and Th2 cytokines has been observed previously by measuring intracellular cytokine production from unstimulated peripheral blood mononuclear cells after 30 min of moderate-intensity cycling (Zaldivar and others 2006). In contrast, the present study examined the functional ability of peripheral blood mononuclear cells to respond to a stimulus (PHA). Interestingly, the release of multiple cytokines was elevated at 4 h postexercise. These responses demonstrate that exercise-induced stress can prime peripheral blood mononuclear cells to respond to stimulation (Dhabhar 2009).
Recently, analysis by magnitude-based inference has received greater use in exercise science and clinical settings. As previously stated, using traditional data analysis, there were no statistically significant changes in Th cell cytokine secretion at any time point during the postexercise period in the present study (Table 3). However, analysis using magnitude-based inferences suggested various changes after exercise, which may prove clinically significant. This approach may provide insights into what are clinically important, yet statistically nonsignificant results in physiological and immunological processes. This study assists in highlighting the importance of considering nontraditional methods for data analysis when dealing with small sample sizes, with high between-subject variability.
The changes that occurred in Th cell cytokine release suggest a multifactorial, prolonged response to heavy endurance exercise. The fluctuations in cytokine production that occurred during recovery from exercise could have important implications for resistance to illness in athletes. While intracellular cytokine analysis after stimulation provides a concise explanation of the Th1/Th2 shift, the current study suggests a much more complex response to exercise.
Footnotes
Acknowledgments
We would like to thank James Keane and Sharni Hardcastle for their technical assistance. and Dr. Sue Hooper for her advice and support. This work was supported by Bond University and the Queensland Academy of Sport Centre of Excellence in Applied Sports Science Research.
Author Disclosure Statement
No competing financial interests exist.