
Abstract
Interleukin-37 (IL-37) is emerging as an important inhibitor of immune response. This study was set up to investigate the expression of IL-37 in Vogt-Koyanagi-Harada (VKH) disease and to explore its possible regulatory role during inflammation. Twenty-four untreated active VKH patients, 10 VKH patients receiving corticosteroids and cyclosporin A (CsA), and 35 healthy controls were included in this study. IL-37 expression in lipopolysaccharides (LPS)-stimulated peripheral blood mononuclear cells (PBMCs) from these 3 groups was assayed by real-time polymerase chain reaction (RT-PCR) and flow cytometry. Cytokines in the supernatants of stimulated PBMCs and CD4+ T cells were assayed by enzyme-linked immunosorbent assay (ELISA). Mitogen-activated protein kinase (MAPK) and nuclear factor kappa B (NF-κB) activation were measured by flow cytometry. VKH patients showed a decreased IL-37 and IL-27 expression and increased IL-1β, IL-6, and tumor necrosis factor-alpha (TNF-α) levels in PBMC culture supernatants. IL-37 significantly inhibited the production of IL-1β, IL-6, and TNF-α, but induced IL-27 expression. VKH patients treated with corticosteroids combined with CsA showed a regression of the intraocular inflammation, and treatment was associated with an enhanced IL-37 production. IL-37 did not affect the production of IL-17, interferon-gamma (IFN-γ), or IL-10 from CD4+ T cells. The present study suggests that a decreased IL-37 expression in VKH patients is associated with a reduced control of the inflammatory response. Treatment of VKH patients with corticosteroids and CsA is associated with an increased expression of IL-37, which suggests that corticosteroids and CsA may partly exert their immunosuppressive effect by upregulating IL-37 production.
Introduction
V
Recent studies revealed that interleukin-37 (IL-37) has a strong anti-inflammatory effect during an innate immune response as well as in the development of inflammation (Nold and others 2010). IL-37 (IL-1F7), one of the members of the IL-1 cytokine family, has 5 splice variants (IL-1F7a–e) (Boraschi and others 2011). It is expressed in human peripheral blood mononuclear cells (PBMCs) and various tissues at low levels and can be induced by inflammatory stimulation, such as Toll-like receptor agonists. Studies on the animal model of colitis revealed that IL-37 could attenuate the clinical signs and ameliorate the histological indices of colitis by decreasing the production of inflammatory cytokines (McNamee and others 2011). These results indicated that IL-37 may participate in feedback mechanisms of an inflammatory response. How IL-37 exactly exerts this role is not yet clear, and whether it might also play a role in the pathogenesis of VKH disease has not yet been addressed. In the present study, we investigated the role of IL-37 in VKH disease, its possible mechanisms of action, and the effects of an immunosuppressive treatment on IL-37 expression.
Materials and Methods
Study subjects
During the study period, blood samples were collected from 34 VKH patients (19 men and 15 women, average age=35 years). Twenty-four were not receiving any immunosuppressive drugs at least 1 month before visiting us, and 10 had been treated with corticosteroids and cyclosporin A (CsA) for at least 2 months (Table 1). Thirty-five healthy volunteers (23 men and 12 women, average age=37 years) served as normal controls between December 2011 and August 2014. The diagnosis of VKH disease was made according to the diagnostic criteria revised for VKH disease by an international committee on nomenclature (Read and others 2001). All untreated patients included in this study typically showed cells and flare in the anterior chamber, mutton-fat keratic precipitates, sunset glow fundus and iris nodules, and at least one extraocular presentation, including tinnitus (11 patients), dysacusis (9 patients), vitiligo (7 patients), poliosis (12 patients), alopecia (14 patients), and headache (13 patients). In the treated patient group, the intraocular symptoms were controlled and only showed sunset glow fundus following corticosteroids and CsA administration.
CsA, cyclosporin A.
All procedures followed the tenets of the Declaration of Helsinki and were approved by the Clinical Research Ethics Committee of Chongqing Medical University with informed consent.
Cell isolation and culture
PBMCs were isolated from heparinized blood by Ficoll–Hypaque density gradient centrifugation. PBMCs were cultured in a Roswell Park Memorial Institute 1640 (RPMI 1640) medium containing 10% fetal bovine serum and 1% penicillin/streptomycin and seeded into 24-well plates at a concentration of 1×106 cells/mL. To analyze the effect of IL-37 on cytokine production, PBMCs from normal controls were plated at 1×106/mL and stimulated with 100 ng/mL lipopolysaccharides (LPS) (Sigma-Aldrich) in the presence of different concentrations of recombinant IL-37 (rIL-37) (0, 50, 100, and 200 ng/mL; R&D Systems) for 72 h, after which the cells and culture supernatants were collected to analyze the mRNA level of cytokines by real-time polymerase chain reaction (RT-PCR) and the protein level by enzyme-linked immunosorbent assay (ELISA). A stock solution of 100 μg/mL rIL-37 was made in sterile phosphate buffer solution (PBS) and appropriate volumes of this solution were added to the medium with PBMCs to reach the required final concentration of rIL-37. To analyze the effect of IL-37 on cytokine production by PBMCs from VKH patients and normal controls, PBMCs were plated at 1×106/mL and stimulated with 100 ng/mL LPS in the presence or absence of 100 ng/mL rIL-37 for 72 h, and cytokines in the culture supernatants were analyzed for cytokines by ELISA. Control groups were stimulated with the same volume of PBS alone.
CD4+T cells were separated from PBMCs from healthy volunteers by magnetic microbeads (both purity >90%; MiltenyiBiotec). CD4+T cells were stimulated with anti-CD3/CD28 (2 μg/mL; eBioscience) in the presence or absence of 100 ng/mL rIL-37 for 72 h to detect the IL-17, IFN-γ, and IL-10 levels in the supernatants.
To analyze the effect of corticosteroids and CsA on IL-37 expression in vivo, 10 VKH patients without any treatment, 10 VKH patients treated with corticosteroids [0.3–0.5 mg/(kg·day−1)] combined with CsA [2–5 mg/(kg·day−1)], and 10 normal controls were included in this study. PBMCs were isolated from these subjects to measure the level of IL-37 by RT-PCR. To analyze the effect of corticosteroids and CsA on IL-37 expression in vitro, PBMCs from 7 healthy volunteers were stimulated with 100 ng/mL LPS in the presence or absence of 500 ng/mL dexamethasone (Sigma-Aldrich) or 100 ng/mL CsA (Novartis) for 72 h.
Real-time quantitative RT-PCR
Total RNA was extracted from isolated PBMCs of VKH patients, normal controls, and rIL-37-stimulated PBMCs using the RNAeasy Mini Kit (Qiagen) according to the manufacturer's instructions. The synthesis of cDNA was performed using the Superscript III Reverse Transcriptase system (Invitrogen). Real-time analysis of IL-37, IL-1β, IL-6, tumor necrosis factor-alpha (TNF-α), IL-27EBI3, IL-27p28, IL-10, and β-actin mRNA was performed using SYBR Premix Ex Taq™ II (TaKaRa) on the ABI 7500 Real-Time PCR System (Applied Biosystems). The primers for the cytokines were as follows:
IL-1β forward 5′-GCAATGAGGATGACTTGTTCTTTG-3′ and reverse 5′-CAGAGGTCCAGGTCCTGGAA-3′, IL-6 forward 5′-GGTACATCCTCGACGGCATCT-3′ and reverse 5′- AGTGCCTCTTTGCTGCTTTCAC-3′, TNF-α forward 5′-ACCTCTCTCTAATCAGCCCTCT-3′ and reverse 5′- GTTGAGGGTGTCTGAAGGAGG-3′, IL-27EBI3 forward 5′-CCGAGCCAGGTACTACGTCC-3′ and reverse 5′- CCAGTCACTCAGTTCCCCGT-3′, IL-27p28 forward 5′-GCGGAATCTCACCTGCCA-3′ and reverse 5′-GGAAACATCAGGGAGCTGCTC-3′, and IL-10 forward 5′-ATGCCCCAAGCTGAGAACCAAGACCCA-3′ and reverse 5′-TCTCAAGGGGCTGGGTCAGCTATCCCA-3′. The primers for IL-37 and β-actin were purchased from Qiagen. Data were normalized to the expression of β-actin using the 2−ΔΔCT method, as described previously (Livak and Schmittgen 2001).
ELISA
The concentration of IL-1β, IL-6, TNF-α, IL-27, and IL-10 in the culture supernatants of PBMCs from VKH patients and normal controls and the concentration of IL-17, IFN-γ, and IL-10 of CD4+T cells from healthy volunteers were measured by human Duoset ELISA kits (R&D Systems) according to the manufacturer's instructions.
Flow cytometry
To analyze the expression of IL-37, PBMCs were fixed with BD Cytofix™ Fixation Buffer (BD Biosciences) at 37°C for 10 min and permeabilized in BD Phosflow™ Perm Buffer III (BD Biosciences) at 4°C for 30 min. Then, the cells were incubated with anti-IL-37 antibody (Abcam) for 30 min at 22°C. The secondary antibody used was PE goat anti-mouse Ig (4A Biotech), which was incubated with the cells for 30 min at 22°C according to the manufacturer's instructions.
Cells were washed and analyzed on a FACScan flow cytometer (BD Biosciences) in terms of mean fluorescence intensity (MFI). Results were expressed as increments relative to the isotypic control (IC) using the formula [MFI of sample−MFI of IC]/MFI of IC. The data were processed using FlowJo software (Reestar, Inc.).
Statistical analysis
Data are shown as mean±standard deviation (SD). The one-way analysis of variance (ANOVA), independent samples t-test, paired sample t-test, Mann–Whitney test, Wilcoxon test, Kruskal–Wallis H test, and Bonferroni correction were applied using SPSS13.0 software (SPSS Inc.). A p-value below 0.05 was considered statistically significant. A description of which test was used for the various experiments performed in this study is shown in the Supplementary Table S1 (Supplementary Data are available online at
Results
Decreased expression of IL-37 in PBMCs from active VKH patients
To investigate the role of IL-37 in VKH, we first compared IL-37 mRNA levels in PBMCs from 20 untreated active VKH patients and 20 normal controls by real-time quantitative PCR. The results showed that the expression of IL-37 mRNA in VKH patients was significantly lower than that observed in normal controls (Fig. 1A).
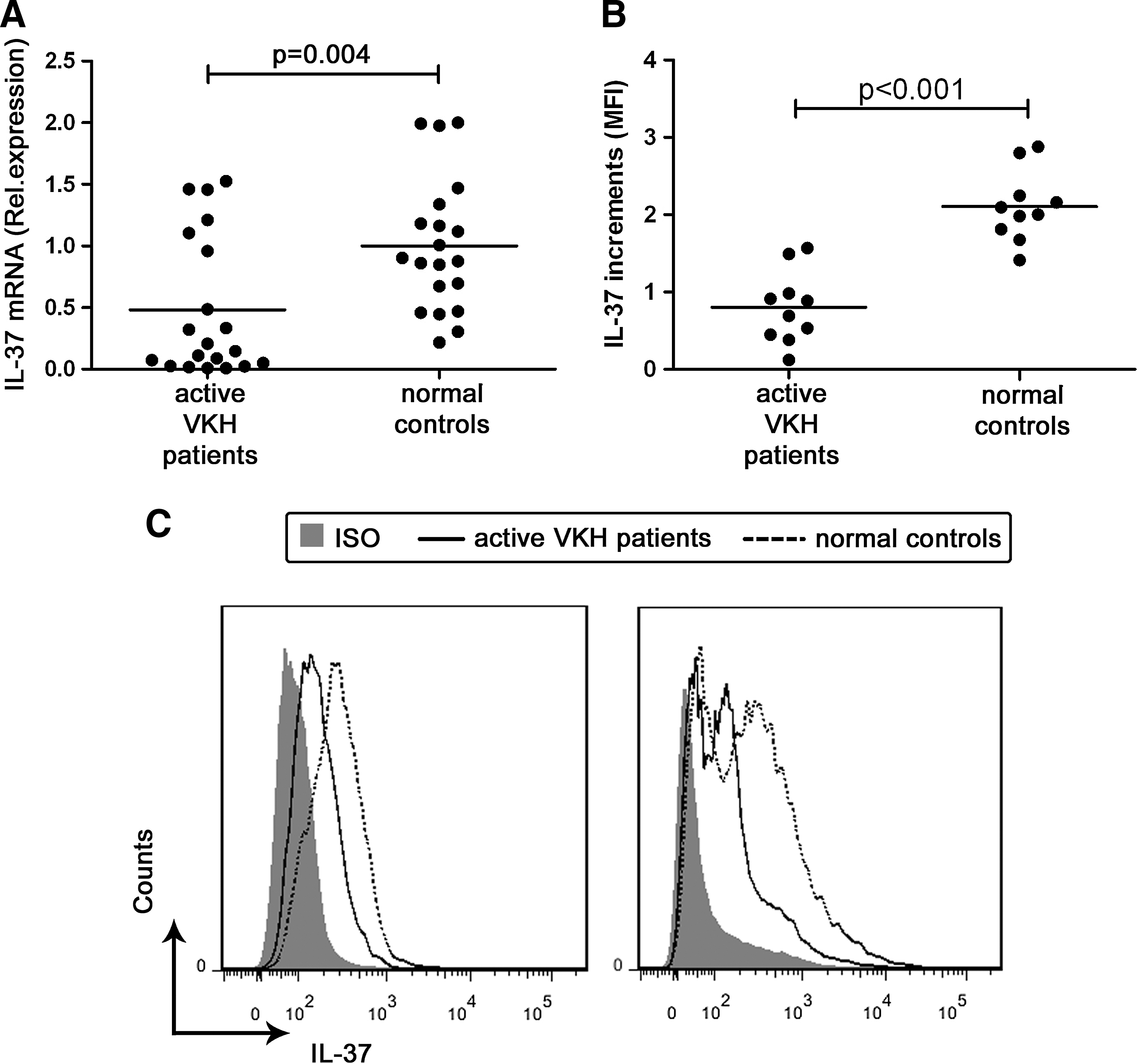
VKH patients have a lower IL-37 level than normal controls.
The protein level of IL-37 in PBMCs from 10 untreated VKH patients with active uveitis and 10 normal controls was determined by flow cytometry. The results were in line with the obtained mRNA data. The PBMC IL-37 protein expression was significantly decreased in active VKH patients as compared with normal controls (Fig. 1B, C).
IL-37 inhibits production of proinflammatory cytokines by PBMCs both in VKH patients and normal controls
To investigate the effect of IL-37 on the proinflammatory cytokine production and whether PBMCs isolated from VKH patients respond differently to IL-37, we performed a series of experiments. The PBMCs were stimulated with LPS in the presence of different concentrations of rIL-37 (0, 50, 100, and 200 ng/mL) for 72 h. The production of several proinflammatory cytokines was determined by RT-PCR and ELISA. The results showed that rIL-37 could inhibit the mRNA level of IL-1β, IL-6, and TNF-α from PBMCs at 100 and 200 ng/mL (Supplementary Fig. S1A–C). The ELISA results were consistent with the RT-PCR results (Supplementary Fig. S1D–F).
To further explore the effect of IL-37 on the proinflammatory cytokine in PBMCs from VKH patients, the PBMCs from VKH patients and normal controls were stimulated with LPS in the presence or absence of 100 ng/mL rIL-37. The concentration of IL-1β, IL-6, and TNF-α in PBMC culture supernatants from untreated VKH patients was significantly higher than that observed in normal controls (Fig. 2). IL-37 inhibited the LPS-induced production of these cytokines both in VKH patients and normal controls. There was no difference in cytokine production when comparing VKH patients with normal controls in their response to IL-37. LPS stimulation was necessary, since earlier experiments from our group showed that the level of these proinflammatory cytokines in culture supernatants obtained from unstimulated PBMCs was very low and these controls were therefore not included in the current study.
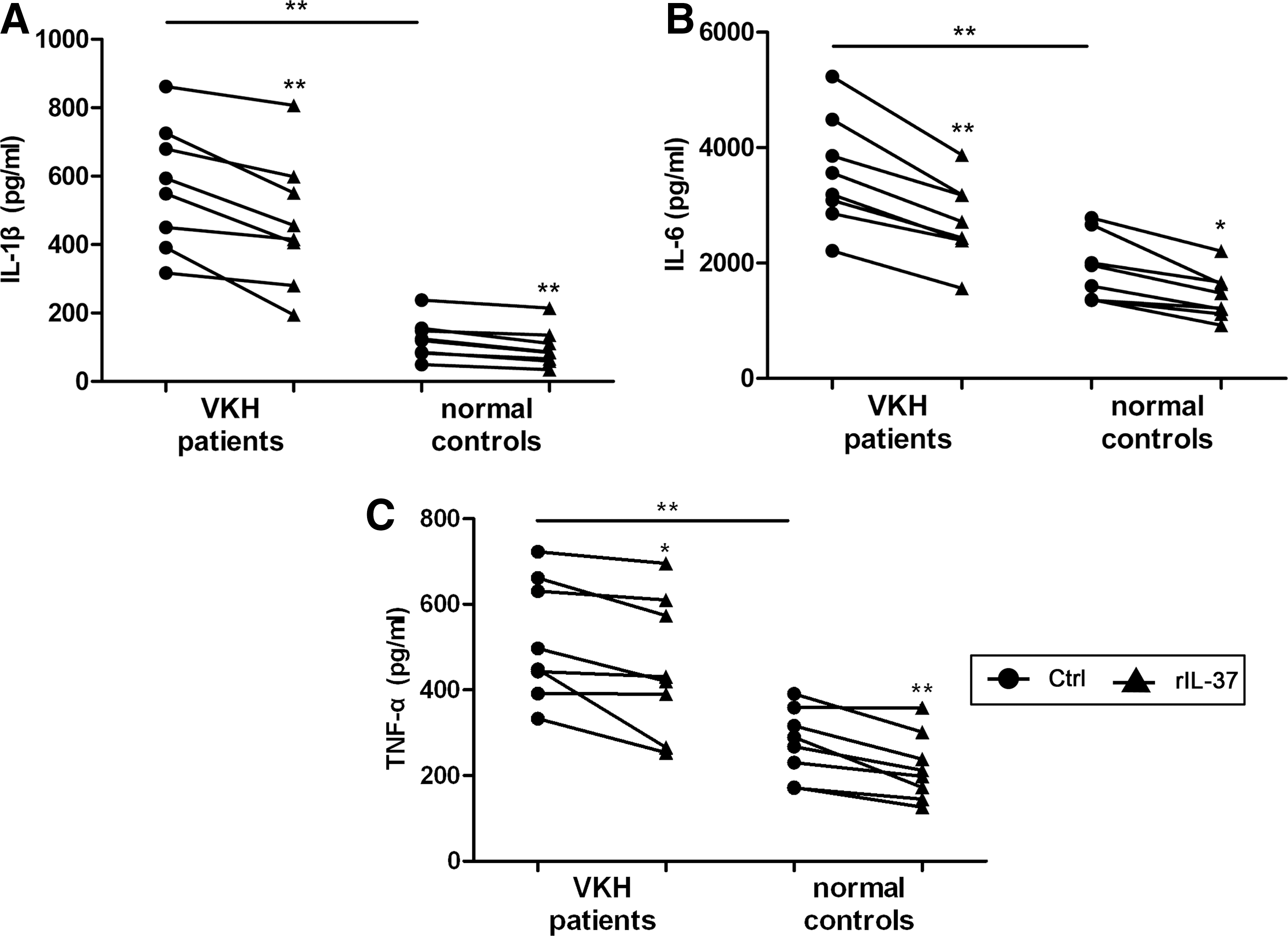
IL-37 inhibits the production of proinflammatory cytokines by PBMCs both in VKH patients and normal controls. PBMCs from active VKH patients without treatment (n=8) and normal controls (n=8) were stimulated with LPS or LPS plus rIL-37 for 72 h. IL-1β, IL-6, and TNF-α expression in the supernatants was measured by ELISA.
IL-37 induces immunosuppressive IL-27 production by PBMCs from VKH patients and normal controls
To investigate the effect of IL-37 on the immunosuppressive cytokines IL-27 and IL-10, we first stimulated PBMCs with LPS in the presence of different concentrations of rIL-37 for 72 h. The results showed that rIL-37 induced the mRNA level of IL-27EBI3 and IL-27p28, 2 subunits of the IL-27 cytokine, in PBMCs and the protein level of IL-27 in cultured supernatants at 100 and 200 ng/mL (Supplementary Fig. S2A, B, and D). No detectable effect on the expression of IL-10 was observed (Supplementary Fig. S2C, D).
The studies on the PBMCs from VKH patients and normal controls showed that the concentration of IL-27 in the culture supernatants was significantly decreased in VKH patients, and the addition of 100 ng/mL IL-37 increased the production by PBMCs both in VKH patients and normal controls (Fig. 3A). No detectable difference was observed in the production of IL-10 by PBMCs, stimulated with or without IL-37, either from VKH patients or normal controls (Fig. 3B).

IL-37 enhances the production of IL-27 by PBMCs both in VKH patients and normal controls. PBMCs from active VKH patients without treatment and normal controls were stimulated with LPS or LPS plus rIL-37 for 72 h. IL-27 and IL-10 expression in the supernatants was measured by ELISA.
IL-37 has no influence on the effector cytokine production by CD4+ T cells
Because of the important role of IL-17, IFN-γ, and IL-10 in the pathogenesis of uveitis (Chi and others 2007; Egwuagu and others 1999; Imai and others 2001; Sun and others 2010), experiments were performed to examine the role of IL-37 on the production of these cytokines by CD4+ T cells. The concentration of cytokines in the supernatants of these cells obtained from healthy volunteers stimulated with anti-CD3/CD28 or anti-CD3/CD28 plus rIL-37 was determined by ELISA. IL-37 did not influence the production of IL-17, IFN-γ, or IL-10 by stimulated CD4+ T cells (Fig. 4). Controls using unstimulated CD4+ T cells showed that the levels of IL-17, IFN-γ, and IL-10 in the culture supernatants were below the detection limit of the assay (data not shown).
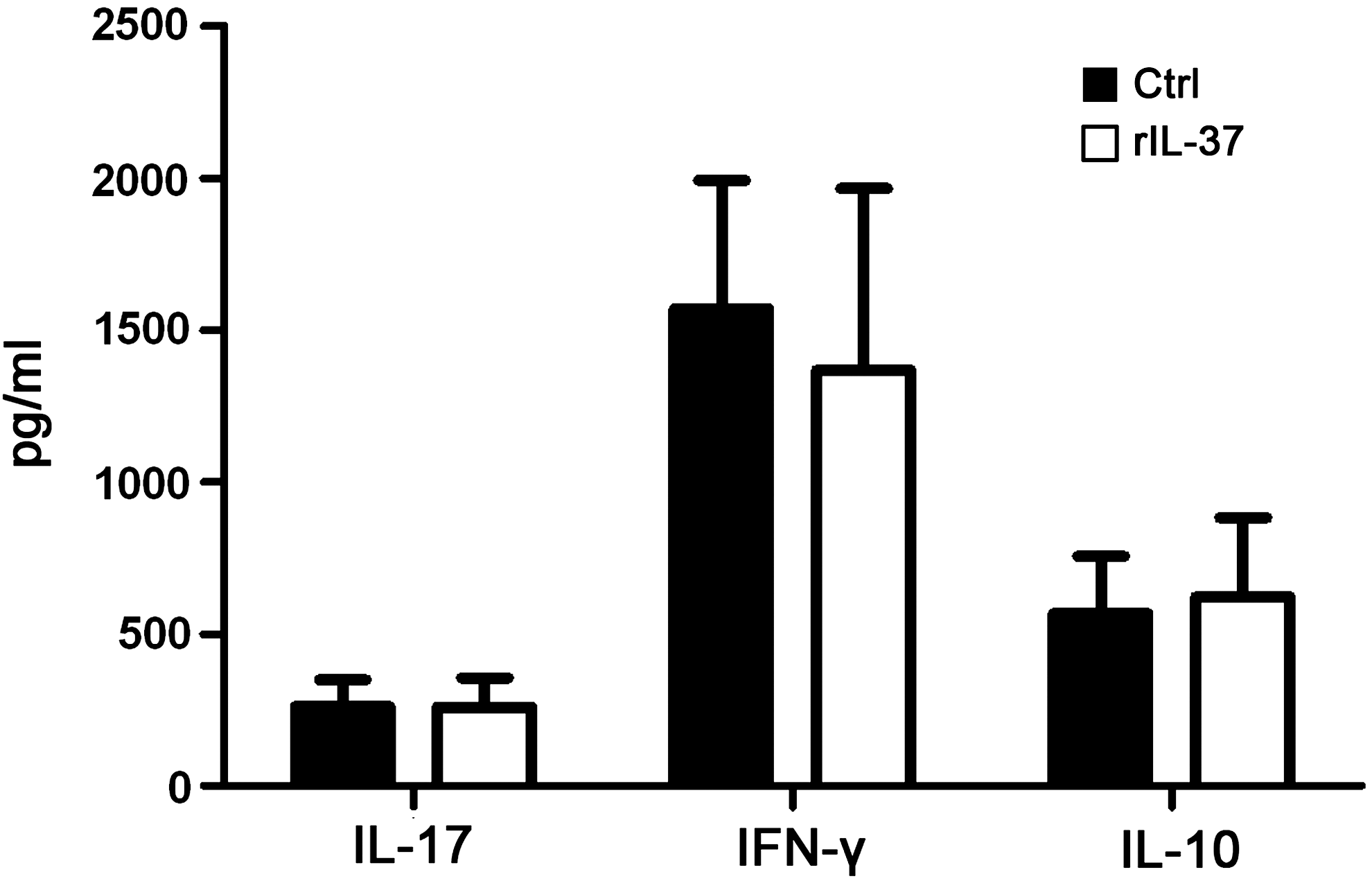
IL-37 has no influence on the production of IL-17, IFN-γ, and IL-10 by CD4+ T cells. CD4+ T cells were isolated from healthy volunteers (n=6) and stimulated with anti-CD3/CD28 in the presence or absence of rIL-37 for 72 h. The concentration of these 3 cytokines in the supernatants was assayed by ELISA. Data are expressed as mean±SD. SD, standard deviation.
IL-37 expression is increased in VKH patients treated with corticosteroids and CsA
Since corticosteroids and CsA are common drugs used to effectively treat VKH disease, we performed a preliminary experiment to investigate the effect of these 2 drugs on the expression of IL-37. The IL-37 mRNA level was measured in PBMCs from 10 VKH patients who had not yet been treated before visiting our uveitis clinic, 10 VKH patients who were on a CsA combined with prednisone treatment regimen, and 10 normal controls. The results showed that the IL-37 mRNA level in untreated patients was significantly lower than treated patients and normal controls. No difference could be detected in the IL-37 mRNA level between treated patients and normal controls (Fig. 5A).
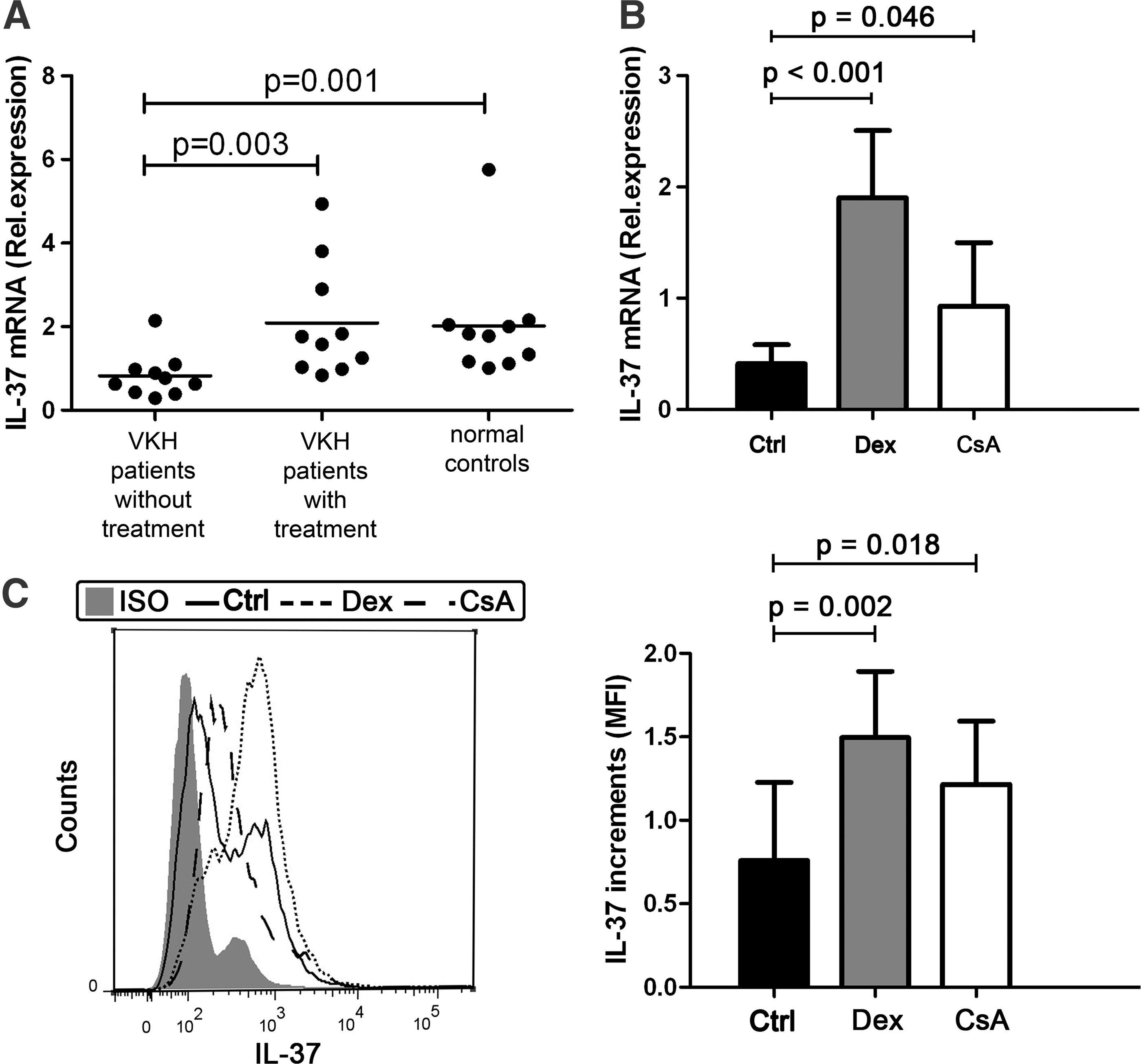
The effect of corticosteroids and CsA on the production of IL-37.
To clarify which of the 2 drugs upregulated the production of IL-37, PBMCs from 7 healthy volunteers were stimulated with LPS in the presence or absence of dexamethasone or CsA for 72 h. The results showed that dexamethasone significantly enhances the mRNA expression of IL-37 in PBMCs and that CsA has a similar but weaker effect (Fig. 5B). We also measured the IL-37 protein level in the dexamethasone or CsA-treated PBMCs by flow cytometry and found results that were consistent with our mRNA expression data.
Discussion
The present study shows a decreased IL-37 expression in PBMCs from VKH patients and an increased production of IL-1β, IL-6, and TNF-α in LPS-stimulated PBMCs as compared with healthy controls. In vitro experiments showed that IL-37 significantly inhibited the production of these 3 proinflammatory cytokines and promoted the IL-27 release by PBMCs. We further showed that treatment with corticosteroids and CsA was associated with an increased IL-37 expression both in vivo and in vitro.
The pathogenesis of VKH disease has been extensively studied during recent years and various cytokines were shown to be involved in its pathogenesis (Chi and others 2007; Commodaro and others 2010; Ooi and others 2006; Wang and others 2012). Earlier studies have already shown that IL-37 has a powerful inhibitory effect on the immune response and protects mice from LPS-induced shock (Nold and others 2010). To investigate whether IL-37 has a role in the pathogenesis of VKH, we first compared the expression of IL-37 in VKH patients with normal controls. The results showed a significantly decreased IL-37 mRNA expression in PBMCs obtained from VKH patients. Flow cytometry also showed a lower IL-37 protein expression in VKH PBMCs. These results indicated that a decreased IL-37 production may be correlated with VKH disease development. Our group recently reported that decreased levels of IL-37 were also found in patients with active Behcet's disease, an autoinflammatory disease (Ye and others 2014). Although VKH disease and Behcet's disease are 2 different types of uveitis, the decreased levels of IL-37 were found in both patient groups. Based on these findings, we conclude that the decreased levels of IL-37 may also be associated with other uveitis entities. The exact association between IL-37 expression and other uveitis entities, however, remains unknown and needs further study. Furthermore, it would be interesting to test the serum IL-37 protein levels in uveitis patients. We used an ELISA to measure IL-37 in serum, but the levels remained below the detection limit of the assay we used.
Other groups found that IL-37 expression was increased in immune-mediated inflammatory conditions, as evidenced by studies in PBMCs in systemic lupus erythematosus (SLE) (Song and others 2013) and synovial cells of patients with rheumatoid arthritis (Boraschi and others 2011). The reasons as to why there is a difference in IL-37 expression between VKH disease and SLE or rheumatoid arthritis are not clear, but may be due to differences in the immunological mechanisms and tissues involved between VKH disease and SLE or rheumatoid arthritis.
To further examine the potential anti-inflammatory properties of IL-37, we evaluated the influence of IL-37 on the inflammatory cytokine production both in VKH patients and normal controls. Our results showed a higher PBMC expression of IL-1β, IL-6, and TNF-α and a lower expression of IL-27, an important immunosuppressive cytokine (Batten and others 2006; Fitzgerald and others 2007; Stumhofer and others 2006), in VKH patients as compared with controls. These results are in agreement with earlier studies showing a decreased serum level of IL-27 in VKH patients (Wang and others 2012) and an increased expression of IL-1β, IL-6, and TNF-α in experimental models of uveitis (Horai and Caspi 2011; Ooi and others 2006). Furthermore, consistent with previous studies (McNamee and others 2011; Nold and others 2010), we found that IL-37 could significantly inhibit IL-1β, IL-6, and TNF-α production and upregulate the release of IL-27 by PBMCs both in VKH patients and normal controls. However, the expression of IL-10, another important immunosuppressive cytokine (Mosser and Zhang 2008; Sabat and others 2010; Ye and others 2007), was not affected by IL-37, either in VKH patients or in controls. These data collectively support the hypothesis that a decreased IL-37 expression, as observed in patients with VKH disease, contributes to the inflammatory events observed in this disease. The factors that control the IL-37 response are as yet unknown, and more study is needed to clarify why IL-37 levels are lowered in VKH disease.
IL-37 has been shown to inhibit the induced expression of the costimulatory molecule CD86 and class II antigen-presenting molecules on dendritic cells in mice (Nold and others 2010). Since dendritic cells (DCs) play a critical role in T cell differentiation (Lanzavecchia and Sallusto 2001; Sallusto and Lanzavecchia 2002), IL-37 may regulate the adaptive immune response. Because of the important role of the Th1 and Th17 response in the development of VKH disease (Chi and others 2007; Sugita and others 2006; Wang and others 2012) and in view of the fact that Foxp3+ Treg cells have been shown to play a role in experimental autoimmune uveitis (Sun and others 2010), we also examined the effect of IL-37 on the effector cytokines of Th1, Th17, and Foxp3+ Treg cells. We were not able to find any effect of IL-37 on the production of IL-17, IFN-γ, and IL-10 by purified CD4+ T cells. IL-37 may thus not influence the cytokine production of T cells directly, but may regulate the adaptive immune response by modulating DCs. This hypothesis is supported by recent experiments from our group showing that rIL-37-treated DCs remarkably inhibited Th17 and Th1 cell responses as compared with control DCs (Ye and others 2014).
Corticosteroids and CsA are extensively used for the treatment of immune-mediated inflammatory diseases and have been shown to be very effective in the resolution of intraocular inflammation in patients with VKH disease (Read and others 2006; Vitale and others 1996). Our preliminary results showed that the expression of IL-37 was decreased in untreated VKH patients. An increased expression of IL-37 was associated with the regression of the intraocular inflammation in patients receiving prednisone combined with CsA. Treatment with corticosteroids and CsA might be able to return the expression of IL-37 back to the levels observed in normal controls. To investigate whether these 2 drugs could regulate IL-37 production directly and whether there was a difference in their relative potency to control IL-37 production, we performed a set of in vitro experiments. Both RT-PCR and flow cytometry analysis showed that corticosteroids significantly enhanced the production of IL-37 by PBMCs and that CsA had a similar effect. Earlier studies showed that IL-37 expressed in NK, monocytes, and stimulated B cells (Boraschi and others 2011), which included in PBMCs, and further studies are needed to identify the subpopulation of cells involved. Our findings are in disagreement with results presented in SLE patients, where a decreased IL-37 mRNA expression in PBMCs was observed following prednisone treatment (Song and others 2013). This may be due to the different role of this cytokine in both diseases. As mentioned above, IL-37 production is high in SLE, whereas it is low in VKH. Further longitudinal experiments are needed to assess the exact response of IL-37 following treatment of VKH patients and how this correlates with the treatment response.
In conclusion, the present study indicates that a decreased IL-37 expression may be involved in the pathogenesis of VKH disease and provides evidence that this may be due to an IL-37-mediated control of pro- and anti-inflammatory cytokine production. Preliminary findings show that treatment of patients with corticosteroids and CsA is associated with an increased IL-37 expression, which suggests that it may contribute to the resolution of intraocular inflammation.
Footnotes
Acknowledgments
This work was supported by the Natural Science Foundation Major International (Regional) Joint Research Project (81320108009), the Key Project of Natural Science Foundation (81130019), Basic Research Program of Chongqing (cstc2013jcyjC10001), the Chongqing Key Laboratory of Ophthalmology (CSTC, 2008CA5003), the Key Project of Health Bureau of Chongqing (2012-1-003), National Key Clinical Specialties Construction Program of China, and the Fund for PAR-EU Scholars Program.
The authors thank all donors enrolled in the present study.
Author Disclosure Statement
None of the authors has a proprietary or financial interest in any product mentioned.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.