
Abstract
Human type I Interferons (IFN-β, IFN-ɛ, IFN-κ, IFN-ω, and 12 subtypes of IFN-α) are a family of pleiotropic cytokines with antiviral, antiproliferative, and immunomodulatory activities. They signal through the same cell surface receptors, IFNAR1 and IFNAR2, yet evoking markedly differential potency. One differentiating factor of IFN-β from other type I interferons is the presence of a consensus sequence (NG) for deamidation. Comparing almost completely deamidated IFN-β-1a with untreated IFN-β-1a, this present study reports the increased activities in 3 in-vitro bioassays testing the antiviral, antiproliferative, and immunomodulatory properties, respectively, of the molecule. Deamidated IFN-β-1a has the potential to improve current therapies in multiple sclerosis, and its ability to potentiate the MHC-Class I expression suggests a clinical benefit in diseases where the downmodulation of the MHC-class I expression plays a role (eg, in immuno-oncology combination therapies or antiviral agents). The present study on IFN-β deamidation adds a new prospective on deamidation as part of a posttranslational modification code that allows the modulation of the biological properties of proteins. Moreover, it underlines the unique IFN-β-1a properties that differentiate this molecule from other members of the type I interferon family.
Introduction
N
IFN-β is part of type I interferon family, which includes IFN-ɛ, IFN-κ, IFN-ω, and 12 subtypes of IFN-α. All members of this family signal through the same cell-surface receptors, IFNAR1 and IFNAR2, albeit evoking differential potencies (Lavoie and others 2011; Thomas and others 2011). The different potencies of type I interferons are modulated by their respective receptor recognition chemistry that lead to different receptor-ligand stabilities of the ternary complex (Kalie and others 2008). A key role is played by the functional plasticity of the IFNAR (Thomas and others 2011). Among the type I interferons, IFN-β seems to play a unique physiological role. Type I IFNs are secreted in abundance in response to viral infection, acting early during the immune response to potentiate antiviral responses and to prime and maintain adaptive immunity. However, unlike other Type I IFNs, IFN-β is constitutively secreted at low amounts by many tissues of the body as a means of maintaining homeostasis and priming cells to maintain a rapid and robust innate and adaptive immune response to subsequent challenge (Gough and others 2012).
IFN-β is a 166-amino acid glycoprotein. The primary sequence contains a deamidation consensus sequence (Asn-Gly) at Asn25 in a structurally flexible loop region. When found in such loops, these deamidation sites are known to be more susceptible to deamidation (Brennan and Clarke 1993; Robinson and Robinson 2001a). Residues susceptible to deamidation have been suggested to act as molecular timers of biological events in which deamidation is seriously disruptive to biological systems, unless they are being used for compensating beneficial biological purposes. Rates of deamidation depend on the primary sequence, the 3-dimensional structure of the protein and solution properties such as pH, temperature, buffer ions, and ionic (Robinson 2002). Elevated temperature and pH, found in-vivo conditions, facilitate deamidation (Yuksel and Gracy 1986).
IFN-β-1b, a recombinant protein expressed in Escherichia coli, as opposed to recombinant IFN-β-1a (expressed in CHO), is not glycosylated and carries some modifications in the primary structure with respect to IFN-β-1a. Consequently, these can be considered as different molecules in terms of structure, activity, and stability (Runkel and others 1998; Antonetti and others 2002; Sega and others 2004).
In the present study, the production of a fully deamidated IFN-β-1a variant and the characterization of its physicochemical and in vitro biological functions in comparison to IFN-β-1a that had not been subjected to deamidation treatment will be reported. To our knowledge, this is the first study reporting the increased immunomodulatory activity of deamidated IFN-β-1a. Due to the identical sequence of recombinant IFN-β-1a and human endogenous IFN-β, highly similar structural and functional properties can be assumed. Consequently, the increased activities, as a result of the deamidation, may be extrapolated also to the endogenous IFN-β.
Materials and Methods
Drug substance used for the characterization study
Highly purified IFN-β-1a drug substance was supplied by Merck Serono S.A. (Switzerland). The production method used a recombinant expression of the protein in a CHO-K1-derived cell line using a commercial serum-free culture medium. Crude harvests were processed through a series of chromatographic steps, which included affinity, ion exchange, reversed-phase, and size-exclusion chromatography. The final drug substance was formulated and stored in sodium acetate with a pH of 3.8. This buffer composition was selected on the basis of formulation development results.
Preparation of artificially deamidated IFN-β-1a
Deamidation was carried out by incubating IFN-beta-1a drug substance into 1.2 M ammonium bicarbonate pH 9.2, at 23°C for 20 h (final ammonium bicarbonate concentration ∼200 mM and IFN-beta-1a concentration ∼0.3 mg/mL). After treatment, the deamidated sample was buffer exchanged by ultrafiltration to 50 mM sodium acetate pH 3.8 and 20 mM sodium phosphate pH 7.2.
Peptide mapping (deamidated and oxidized forms)
The samples were mixed with the digestion/reduction buffer to obtain the correct pH value and to reduce the protein using dithiothreitol contained in the buffer. Endoproteinase Lys-C was added to the sample for digestion. The sample was incubated at 5°C± 3°C for 6 h. After incubation, a small volume of water:TFA 5% is added to adjust the pH to a value of 3–3.5. The chromatographic separation for the analysis of the desamido forms uses water:TFA (0.2%)/ water:ACN:TFA (30%:70%:0.2%) gradient on a C18 BEH ultraperformance liquid chromatography (UPLC) column 50 × 2.1 mm 1.7 μm at a temperature of +40°C. The flow rate was set at 0.6 mL/min and detection was performed by UV adsorption at 214 nm wavelength, sample injection volume 30 μL. The chromatographic separation for the analysis of oxidized forms uses a water:MSA (2.5 mM)/water:ACN:
MSA (30%:70%:2.5 mM) gradient on a C18 BEH UPLC column 50 × 2.1 mm 1.7 μm at a temperature of +20°C. The gradient ranges from 6% of B1 held for 1 min to 30% over a 29-min time period and then to 32% over a 30-min time period. The wash was performed at 100% B1 for a 10-min time period. The flow rate was set at 0.6 mL/min and the detection was performed by UV at 214 nm wavelength, with a sample injection volume of 40 μL.
Electrospray ionization–mass spectrometry whole molecule analysis
Prior to analysis, samples were ultrafiltrated and subsequently analyzed by means of direct infusion. The electrospray ionization–mass spectrometry (ESI-MS) method, which was applied, is based on a technique described in the European Pharmacopoeia monograph (Nr. 1639E, Interferon beta 1a concentrated solution) with adaptations for the designated instrument. The analyses were performed on an Orbitrap Mass Spectrometer (Thermo Scientific) with the following instrumental settings: Scan ES (+) from 100 to 2000 m/z with a resolution set at 100,000; scan time 1 microscan 500 ms; capillary temperature: 200°C; capillary voltage 35 V, tube lens 145 V. The calibration of the Orbitrap Mass Spectrometer was set at 5 ppm (m/z) of mass error. However, for highly multicharged ions when referring to intact proteins such as IFN-β-1a, the obtained average molecular weight accuracy is around 100 ppm. After the acquisition, the m/z spectra were semiautomatically deconvoluted by using the XTract software package (Thermo Scientific). The expected molecular weights were calculated on the basis of the amino acid sequence, the correct disulfide bridge, and the contribution of the attached oligosaccharide structure. The identities of the detected glycan moieties have been verified in previous more detailed studies involving HILIC-UPLC glycan profiling of fluorescently labeled glycan standards, sequential exoglycosidase digestions, and fragmentation analysis (MS/MS).
Size exclusion chromatography
The monomeric purity of the samples was analyzed by size exclusion HPLC analysis on a Waters HPLC system (Waters) by using a TSK G2000 SWXL column (Toso Bioscience), elution buffer sodium acetate pH 3.8 containing 50 mM sodium chloride. The flow rate during elution was set to 0.5 mL/min and the eluent was monitored by UV absorption at a wavelength of 215 nm.
Protein content determination by reversed-phase ultraperformance liquid chromatography
RP-HPLC analysis was performed on an Acquity UPLC BEH300 C4 1.7 μm 2.1 × 50 mm column (Waters) connected to an Acquity UPLC system (Waters) equipped with a TUV detector (Waters). A gradient of 40%–70% acetonitrile was used at a flow rate of 0.45 mL/min. All experiments were performed at 25°C. Quantification was performed by the use of a calibration curve based on a reference house standard.
Higher-order structure analysis by circular dichroism
Analysis of the secondary and tertiary structure of the samples was performed by circular dichroism (CD) analysis performed in the Far-UV and Near-UW region, respectively. The analyses were performed on a Jasco J-810 Spectropolarimeter equipped with a Peltier temperature controller PTC-423S (Jasco). In the far-UV region (200–250 nm) a 1 mm path length QS cuvette and in the near-UV region (250–300 nm) a 1 cm path length QS cuvette were used. Spectra were normalized for protein concentration by dividing the CD signal by the UV absorption of the respective sample measured at 215 nm, and blank spectra of the respective buffer matrix of the sample (50 mM sodium acetate pH 3.8 or 20 mM sodium phosphate pH 7.2) were subtracted. The following measurement parameters were used: data pitch 0.1 nm, bandwidth 1 nm, response time 4 s, scan speed 20 nm/min, and accumulations 5.
Isothermal chemical denaturation
All denaturation experiments were carried out by using the Unchained Labs HUNK Automated Chemical Denaturation System with fluorescence detector (Unchained Labs). The protein concentrations for the deamidated and the untreated IFN-β-1a were ∼19 M (in 50 mM sodium acetate pH 3.8 and 20 mM sodium phosphate pH 7.2 mM, respectively). The protein concentrations for the deamidated and the untreated IFN-β-1a were 19 μM in all experiments. Using urea as a denaturant, 32-point denaturation curves were recorded. All 4 samples exhibited a single unfolding transition and, therefore, the experimental data were fitted to a 2-state unfolding model. The excitation wavelength was set at 280 nm and the unfolding transition was monitored by the fluorescence signal set at 350 nm. The conformational stability of a sample was expressed as its Gibbs energy, ΔG°, which is determined from the denaturant concentration at the transition midpoint (C1/2, 50% unfolded) and the change in ΔG as a function of the denaturant concentration (m) in accordance to the equation ΔG° = m * C1/2.
Differential scanning calorimetry
Thermal denaturation experiments were performed using a Nano differential scanning calorimetry (DSC) Model 602001 (TA Instruments). The samples were concentrated in their respective buffers (50 mM sodium acetate pH 3.8, and 20 mM sodium phosphate pH 7.2 mM, respectively) to a concentration of ∼5.2 mg/mL. Before temperature scanning, the samples were equilibrated for 10 min at +10°C. The scanning rate was set at 1°C/min and the applied temperature range was at 20°C–90°C. The obtained results were baseline corrected by first defining the baseline range (range set between 50°C and 80°C) and then, creating a polynomial baseline to fit the thermal transitions.
Antiproliferative activity (WISH cells) assay
This assay is based on a method described by Grossberg and others (1986). It measures the ability of IFN-β-1a to inhibit the growth of the WISH cells. In summary, the WISH cells were plated in a 96-well microtiter plate, which contained IFN-β-1a in a range of concentrations and then incubated for 72 h at a temperature of 37°C. At the end of the incubation period, the cells were stained with MTT and the optical density (OD at 595 nm) of each well was measured by using a multiwell scanning spectrophotometer.
A549/encephalomyocarditis virus antiviral activity
This assay is based on a method reported by Grossberg and others (1986) and is described in the European Pharmacopoeia 5.0. Chapter 5.6: Assay of Interferons. Briefly, this assay measures the antiviral activity of IFN-β-1a by its protection exerted on the A549 cells (human lung carcinoma cells) against the cytopathic effect of the encephalomyocarditis virus (EMCV). In summary, the A549 cells were plated in a 96-well microtiter plate, which contained IFN-β-1a in a range of concentrations and then incubated for at least 6 h at 37°C. At the end of the incubation time, the medium was removed. The EMCV suspension was added to each well and a 1-h reaction time period is given at 37°C. After ∼30 h of incubation at 37°C, the cells were stained with Naphthol Blue Black (NBB) and the OD set at 620 nm of each well was measured by using a multiwell scanning spectrophotometer.
Immunomodulatory activity (MHC expression)
This assay is based on a method described by Grossberg and others (1986). The assay principle is based on the ability of IFN-β-1a to upregulate the MHC class I expression on the A549 cells in a dose-related manner. The A549 cells are incubated with different concentrations of IFN-β-1a for 48 h at 37°C. Then, the cells are harvested by using trypsin/EDTA and the expression of the MHC class I is evaluated by FACS analysis using a FITC-conjugated anti-hMHC I antibody and expressed in terms of mean fluorescence intensity.
In vitro bioassay data normalization and assessment
The results of the individual samples are expressed as a biological response in relation to a standard sample run in parallel in each analytical session and set to be equal to 100%. The biological activity was derived from dose–response curves that were constructed on a mass basis using the protein content (mass per volume) determined for the individual fractions by reversed-phase ultraperformance liquid chromatography (RP-UPLC). Moreover, to facilitate their comparison, the relative response of each sample was normalized against the relative response of their corresponding start material. One-way analysis of variance (ANOVA) has been performed for each bioassay to compare the mean potencies of the deamidated versus the untreated sample.
Results
IFN-β-1a artificial deamidation and physicochemical characterization
CHO-expressed IFN-β-1a (drug substance buffered at pH 3.8) was placed under deamidation-inducing conditions at alkaline pH as described in the Materials and Methods section. One part of the deamidated material was rebuffered to the original drug substance pH of 3.8, whereas the other part was rebuffered to physiological pH (7.2). As a control, another aliquot of the untreated starting material was rebuffered to pH 7.2. The aim of this experimental design was to compare the effects of the deamidation at 2 pH levels (ie, 3.8 and 7.2 respectively, with the related untreated control samples, thus deamidation-treated versus untreated, respectively at pH 3.8 and 7.2). The resulting samples and controls were first analyzed by a panel of physicochemical methods to assess the effect of the deamidation treatment on the integrity of the protein.
Peptide mapping was performed to assess the level of deamidation and potential other posttranslational modifications. Size exclusion chromatography was performed to assess the monomeric purity of the protein. CD and intrinsic fluorescence spectroscopy were performed to assess a potential impact on the secondary and tertiary structure of the molecule. Furthermore, structural stability of the deamidated and the untreated IFN-β-1a drug substance was compared by DSC and isothermal chemical denaturation (ICD), which probe the unfolding as a response to thermal and chemical stress, respectively. Table 1 summarizes the results obtained by the physicochemical characterization.
The percentage level of desamido forms represents the sum of peptides in which Asn25 is converted into Asp, succinimide and isoaspartic acid compared with the corresponding unmodified peptide.
DSC, differential scanning calorimetry; ICD, isothermal chemical denaturation; IFN, Interferons.
These analyses reveal that the treatment resulted in an almost complete deamidation (>95% compared with ∼25% as baseline in the untreated control) without any significant impact on the aggregation. Peptide mapping further shows that the treatment did not increase the methionine oxidation level of the protein (data not shown). In addition, the glycosylation was analyzed by means of ESI-MS, which showed that the distribution of the glycoforms was not affected by the deamidation treatment and that the obtained glycoform profiles corresponded to the IFN-β isoforms described in the European Pharmacopoeia monograph (termed glycoforms A–E), in which the biantennary, disialylated core-fucosylated species is the main IFN-β glycoform (data not shown).
In terms of structural stability both the buffer pH as well as the deamidation had significant impact. Both the DSC and the ICD have revealed significant differences in terms of melting temperatures and the Gibbs energy ΔG, respectively, which means that both the deamidated and untreated molecules are more stable at pH 3.8 than at pH 7.2. When comparing the deamidated forms with the untreated forms at their respective pHs, both analyses showed that the deamidated variant is more stable than the untreated variant. Figure 1 depicts the differences in the unfolding behavior of the 2 variants under chemical stress conditions, which were applied during the ICD analysis. At both pHs, ΔG is higher for the deamidated samples. All measurements yielded high R2 values (goodness of fit) indicating that the model faithfully represents the experimental data (32 data points per curve) and that the obtained differences in terms of ΔG are significant and not attributable to instrument- or measurement-related variation (Table 1). It is worth noting that R2 values at pH 7.2 are slightly lower compared with the tight fit at pH 3.8, which is consistent with a higher degree of disorder observed at pH 7.2. Deamidation thus renders the molecule more stable against thermal (DSC) and chemical stimuli (ICD).

Isothermal chemical denaturation (ICD) curves of artificially deamidated IFN-β-1a compared with the untreated control. The figure shows duplicate analyses of untreated (blue and green profiles) and deamidated (red and pink profiles). IFN, Interferons.
The Far and Near-UV CD spectra of the samples are shown individually in Fig. 2 and in Fig. 3. The structural analysis in the Near-UV region (Fig. 2) shows that both the deamidation treatment (Panel A) and the pH (Panel B) impact on the tertiary structure. At pH 7.2, the deamidation alters the spectrum, mainly seen in the wavelength region between 270 and 295 nm, whereas at pH 3.8, the same region was hardly affected while the difference becomes visible in the region between255 and 265 nm. A shift in the pH affects the untreated control, mainly seen in the wavelength range between 250 and 260 nm, whereas for the deamidated molecule, intensity differences are visible in features over a wider range between 255 and 300 nm. In the Far-UV region (Fig. 3), the spectra of the deamidated sample and the untreated control overlap well at pH 3.8 and pH 7.2, respectively. However, a slight shift in the secondary structure, both for the deamidated sample and the untreated control, is visible as a pH effect.
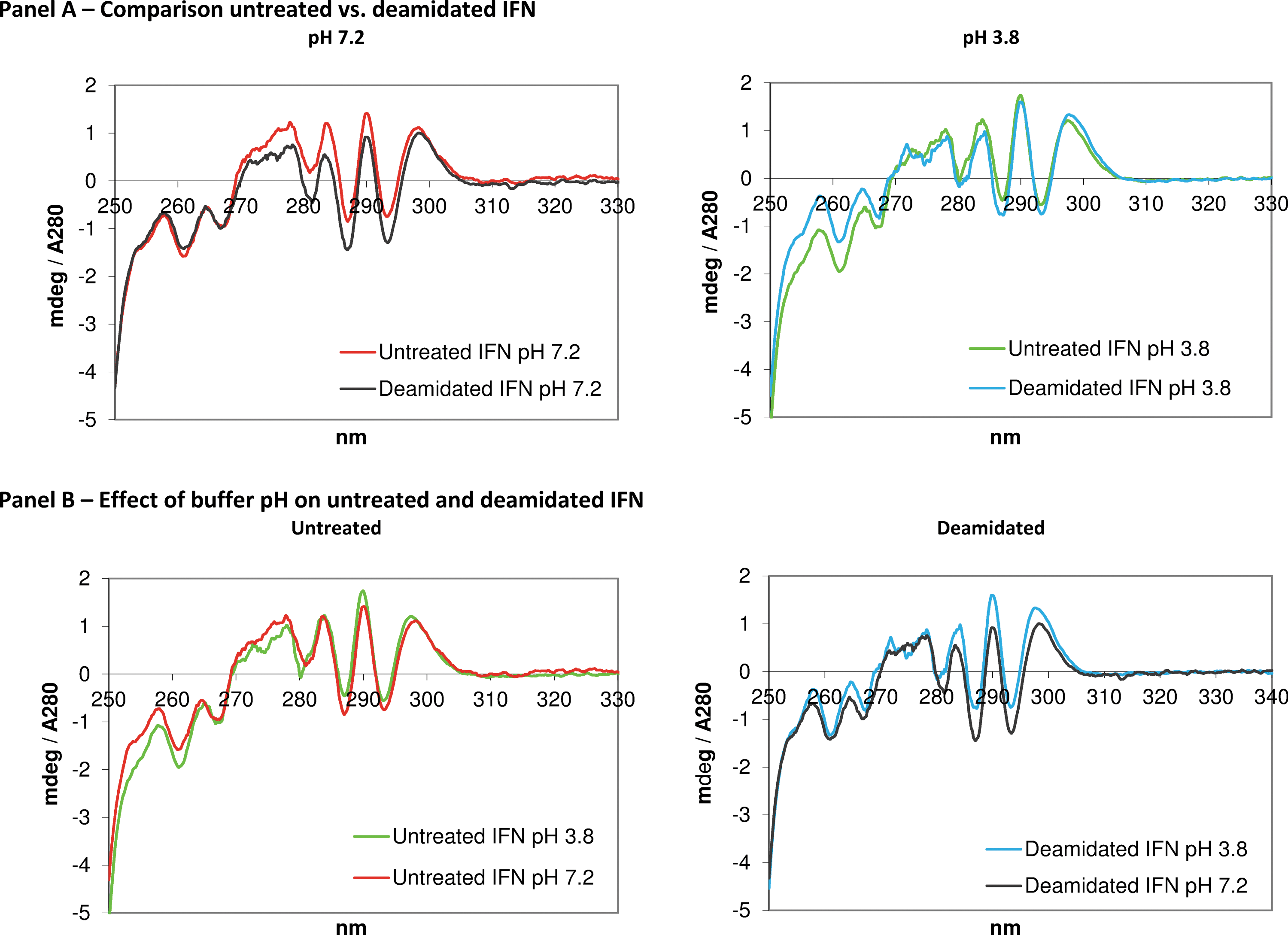
Near-UV CD spectra of deamidated and untreated IFN-β-1a.

Far-UV CD spectra of deamidated and untreated IFN-β-1a.
Biological characterization
The biological characterization of the deamidated IFN-β in comparison with the starting material is reported in Fig. 4. In this study, it is interesting to note that in all tests that were performed, the deamidated material showed a higher activity compared with the untreated control. The in-vitro bioactivity ratios of the deamidated versus control for the antiproliferative, antiviral, and immunomodulatory assays were found to be 2.1-fold, 1.5-fold, and 2-fold, respectively. One-way ANOVA confirmed that for each bioassay there is a statistically significant difference between the mean potency from the deamidated sample compared with the untreated control at the 95.0% confidence level. The obtained P values were: antiproliferative assay 0.0277, antiviral assay 0.0353, and immunomodulatory assay 0.0011.
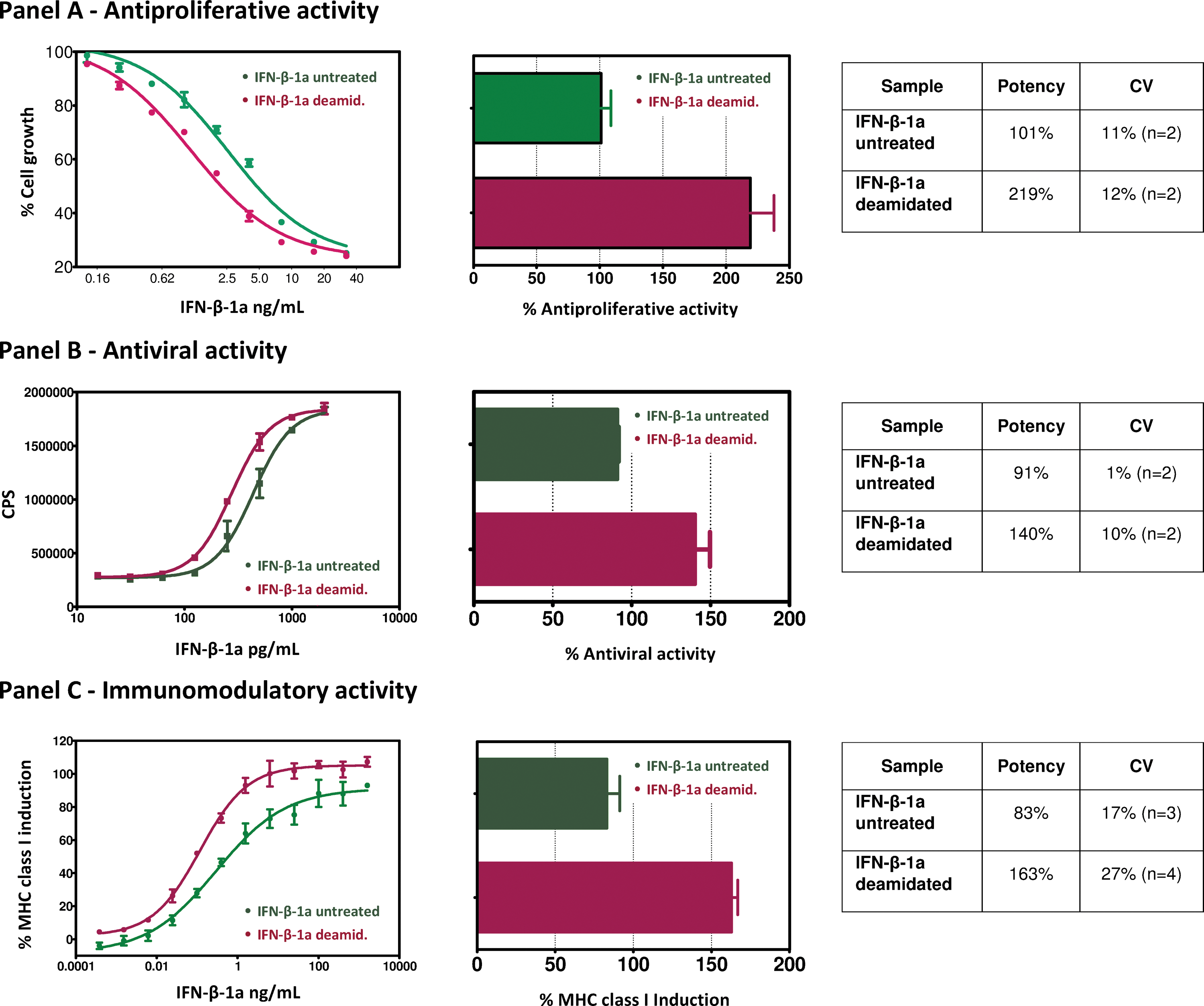
In vitro antiproliferative
Discussion
Asparagine deamidation is one of the degradation routes of proteins that convert asparagine into aspartic/isoaspartic acid. This conversion alters the net charge of the protein due to the fact that the neutral amide side chain of the asparagine is converted into the acidic carboxyl group of the aspartic acid. This can have varying effects on the physiochemical and functional stability of a protein and can affect the shelf-life of a product that is not formulated or stored appropriately (Chelius and others 2005; Wakankar and Borchardt 2006). Asparagine deamidation is considered a major pathway for the chemical degradation of therapeutic proteins. Moreover, deamidation is also generally considered a molecular clock of biological events such as protein turnover, development, and aging (Robinson 2002). Depending on the localization of the residue, deamidation may be seriously disruptive to biological systems unless it exerts a beneficial biological effect (Robinson and Robinson 2001b).
The present study reports the generation, physicochemical, and biological characterization of a deamidated IFN-ß-1a. Almost complete deamidation was found after treatment versus ∼25% of the untreated sample. The conditions used to achieve deamidation resulted in negligible amounts of protein-related impurities, such as oxidation, aggregation, fragmentation, and glycosylation, which were not affected by the deamidation treatment.
Our results clearly demonstrated that the deamidation treatment positively affects the biological properties of IFN-ß-1a. In particular, the deamidated IFN-β1a, compared with the untreated control, exhibited a 2.1-fold, 1.5-fold, and 2-fold increase, respectively, in antiproliferative, antiviral, and immunomodulatory activities.
It may be speculated that the enhanced biological properties result from the increased stability of the deamidated molecule as evidenced by the chemical and thermal unfolding experiments, which revealed a higher resistance against denaturation for the deamidated variant. In the IFN-β1a 3D-structure, the LeuAsn25Gly residues are located in the AB1 loop as described by Karpusas and others (1997). This is a nonstructured and exposed region, which may facilitate the deamidation reaction in comparison to corresponding motifs located in more structured regions. The deamidation rate of a given Asn residue can significantly vary depending on the environment of the motif as demonstrated by Robinson and Robinson (2004). For example, the authors observed that for the same motif (LNxG) deamidation coefficients, a measure of the deamidation rate (in which lower coefficients represent higher deamidation rates) of 0.04 (mouse interleukin-1β, motif in a nonstructured region) and 125 (human interleukin-2, motif in a structured region), respectively.
The crystal structure reported by Karpusas and others (1997) has revealed that the AB1 portion of the loop, in which the deamidation sequence is located, is stabilized by a hydrogen bond between the side chain Nɛ of Arg147 and the backbone carbonyl of Leu24, in both murine and in human IFN-β. Therefore, the Asn25 is in close proximity to the Thr144 and a positively charged Arg147 residue. Accordingly, the deamidation reaction to the Asp25, (which generates a negative charge), may further be facilitated by the resulting electrostatic interactions with the spatially close and positively charged Arg147, as well as by the possibility to form additional hydrogen bonds with the surrounding residues. Such interactions promote intrachain interactions potentially resulting in increased stability against chemical and thermal stimuli.
An alternative explanation could be the alteration of the local structure due to deamidation, which may be transmitted and amplified to a distant region, as suggested for the opposite effect of alkylation of free cysteine reported to increase a non-native state conformation without altering the binding with IFNAR2, but leading to inactive IFN-β form through a yet unknown allosteric modification that has a deleterious effect on a distant region responsible for recognition to IFNAR-1 (Bobst and others 2008; Kaltashov and others 2010). In the case of deamidation, the suggested alteration of the local structure may lead to an allosteric modification that in this case results in the observed increasing activity likely due to an optimized and differential interaction of the ternary complex with IFNAR1.
The overall architectures of the receptor binding of IFNα2 and IFNω has been shown to be nearly identical (Thomas and others 2011). It is likely that all subtype I IFNs adopt this ternary complex architecture and different IFNs can induce differential potencies as a result of distinct receptor-binding chemistries and stabilities of ternary complexes. Sequence alignment of type I IFNs and residues reported to be relevant for the formation of the ternary complex (Thomas and others 2011) indicate that the Asn25 is part of the binding regions with IFNAR2. However, IFN-ω and IFN-α in this position replace the Asparagine with an Arginine residue and within the surrounding environment, whereas the Arg147 residue is conserved, and the Thr144 is replaced by glutamic acid. Consequently, this specific microenvironment created by the deamidated Asn25 interacting with the Arg147 and the Thr144 is a unique feature of IFN-β, thus differentiating it from other type I interferons. Moreover, other type I interferons (IFN-α, IFN-ω) do not contain the deamidation consensus sequence able to modulate the IFN biological activity, which is a further differentiation of IFN-β within this family of proteins, all signaling through the same cell-surface receptors, IFNAR1 and IFNAR2. Deamidation may thus be part of a regulation mechanism for the bioactivity of IFN-β. For example, considering that deamidation increases with temperature, it may be speculated that a higher deamidation rate of IFN-β, as induced under physiological conditions of fever, will contribute to the defense mechanisms of the host by increasing the bioactivity of the endogenous IFN-β.
In conclusion, the present study reveals that asparagine deamidation enhances the structural stability of the IFN-β-1a molecule. Furthermore, the in vitro results from our studies demonstrate that the deamidated IFN-β-1a has improved biological properties in comparison to the current IFN-β-1a preparation. This study adds a new prospective on deamidation as part of a posttranslational modification code that allows the modulation of the biological properties of proteins. The increased activity of deamidated IFN-β further differentiates IFN-β from other type I interferon family members. Moreover, modulation of the biological properties of IFN-β-1a may improve current therapeutic options or extend the clinical therapeutic potential of IFN-β-1a. The enhanced biological properties of IFN-β-1a may, therefore, improve current therapies in MS or translate into a clinical benefit in diseases where the downmodulation of the MHC-class I expression plays a role (eg, in immuno-oncology combination therapies or antiviral agents). However, to pursue such routes, further studies will be required to investigate a potential immunogenicity of the deamidated variant of IFN-β-1a.
Footnotes
Acknowledgments
The authors would like to thank the members of the Bioanalytical Development and Protein Chemistry Departments of Merck Serono's Analytical Development Biotech Products organization for their efforts and dedication regarding this study. The contribution of the ICD data by Unchained Labs (Rick Brown, Rick Gordon, and Patrick Ross) is greatly acknowledged.
Author Disclosure Statement
No competing financial interests exist.