
Abstract
Eosinophils constitute an important component of helminth immunity and are not only associated with various allergies but are also linked to autoinflammatory disorders, including the skin disease psoriasis. Here we demonstrate the functional relationship between eosinophils and plasmacytoid dendritic cells (pDCs) as related to skin diseases. We previously showed that pDCs colocalize with neutrophil extracellular traps (NETs) in psoriatic skin. Here we demonstrate that eosinophils are found in psoriatic skin near neutrophils and NETs, suggesting that pDC responses can be regulated by eosinophils. Eosinophils inhibited pDC function in vitro through a mechanism that did not involve cell contact but depended on soluble factors. In pDCs stimulated by specific NET components, eosinophil-conditioned media attenuated the production of interferon α (IFNα) but did not affect the maturation of pDCs as evidenced by the unaltered expression of the costimulatory molecules CD80 and CD86. As pDCs and IFNα play a key role in autoimmune skin inflammation, these data suggest that eosinophils may influence autoinflammatory responses through their impact on the production of IFNα by pDCs.
Introduction
E
Despite the protective role of eosinophils in host immunity, such as toxicity against parasitic helminths, the damaging effects of these cells in allergic diseases are widely acknowledged. However, the role of eosinophils in autoinflammatory responses, including the skin disorder psoriasis, remains less clear. The eosinophil levels in blood increase during helminthic infections and allergy, and eosinophils accumulate in the affected tissues (Rothenberg and Hogan 2006). Likewise, blood eosinophilia, significantly elevated serum levels of the eosinophil marker ECP, and an increase in eosinophils in duodenal stroma has long been reported in psoriasis and other autoinflammatory diseases (Rothenberg and others 2001; Mansur and others 2008). Gut-infiltrating eosinophils were detected by ECP-specific Abs or EG2 Abs that recognize both ECP and EDN (Michaelsson and others 1996; Nakajima and others 1999). However, it is unclear if psoriatic skin is infiltrated with eosinophils.
In contrast to allergic dermatitis (eg, eczema) eosinophils are rarely detected in sections of psoriatic skin by a routine histology staining, but a subset of patients with psoriasis has lesions that resemble eczema (Lundin and others 1990; Guttman-Yassky and others 2011). ECP- and EG2-positive eosinophils were reported in human psoriatic skin lesions (Lundin and others 1990; Michaelsson and others 1996). ECP immunoreactivity was detected both intracellularly and extracellularly in skin biopsies, suggesting that at least some eosinophils are degranulated in psoriatic skin. Notably, ECP-positive cells were observed in association with groups of granulocytes, suggesting there is an interaction between these cells in psoriatic skin. Significantly increased ECP levels were also found in suction—blister fluid from lesional psoriatic skin compared to uninvolved skin from the same patients and control individuals (Lundin and others 1990). Additional support for the possible involvement of eosinophils in psoriasis comes from a mouse model of psoriasis-like pathology that showed increased expression of the eosinophil-specific chemoattractant eotaxin in skin lesions (Zenz and others 2005). Taken together, these data suggest the presence of eosinophils in involved skin and that there is possible cross talk between eosinophils and other immune cells that infiltrate lesional psoriatic skin, such as neutrophils and plasmacytoid dendritic cells (pDCs) that closely associate with the neutrophils (Skrzeczynska-Moncznik and others 2012).
We and others have recently demonstrated that the affected skin of psoriatic patients contains neutrophil-derived deposits of extracellular DNA known as neutrophil extracellular traps (NETs) (Villanueva and others 2011; Skrzeczynska-Moncznik and others 2012, 2013). NETs were first reported to clear microorganisms through catching and killing entrapped microbes with antimicrobial agents decorating the NETs (Brinkmann and others 2004; Urban and others 2009). However, recent advances have implicated NETs in various pathologies, including autoinflammatory diseases, systemic lupus erythematosus and psoriasis (Majewski and others 2016). NET constituents, including self-DNA, histones, and myeloperoxidase, are potential autoantigens (Kessenbrock and others 2009; Garcia-Romo and others 2011; Lande and others 2011).
In psoriatic skin, NETs are likely to provide stimulatory signals for pDCs, which are key IFNα- and IFNβ-producing immune cells (Liu 2005). The pDCs express TLR9, and the recognition of DNA by this receptor triggers the production of copious amount of IFNα or IFNβ, which is also referred to as type 1 interferon (IFNI). Whereas genes encoding IFNα come in variety of subtypes, including IFNA1, IFNA2, IFNA6, and IFNA8, IFNβ is encoded by a single IFNB gene (Ivashkiv and Donlin 2014). In addition to IFNI, pDCs were recently shown to express high levels of type III interferon, which includes 3 members; IFNλ1, also known as IL29, and IFNλ2 and IFNλ3 (Yin and others 2012).
IFNI plays a key pathogenic role in autoinflammatory diseases (Nestle and others 2005; Cichy and Zabel 2011; Garcia-Romo and others 2011; Lande and others 2011). Intracellular TLR9 is known to efficiently recognize microbial DNA but can also sense self-DNA. Although self-DNA is typically not immunogenic, when coupled with factors such as antimicrobial peptide-LL37 (Lande and others 2007) or human neutrophil elastase (HNE) together with secretory leukocyte protease inhibitor (SLPI), (Majchrzak-Gorecka and others 2016), strong activation of pDCs can result. In the lesional skin of psoriasis patients, SLPI together with HNE and DNA is a component of NETs, and these SLPI- and HNE-decorated DNA structures are found adjacent to skin-recruited pDCs (Skrzeczynska-Moncznik and others 2012).
In this study, we show that the ability of pDCs to produce IFNα in response to HNE- and SLPI-competent DNA deposits is significantly downregulated by soluble factors derived from eosinophils. We also demonstrate that eosinophils can be found in close proximity to NETs in psoriatic skin. Together, these data suggest a regulatory role of eosinophils in the production of IFNα by pDCs, which has potential relevance in psoriasis and other autoinflammatory diseases.
Materials and Methods
Materials
Recombinant SLPI was purchased from R&D Systems or was produced as previously described (Zabieglo and others 2015). HNE and cathepsin G (CatG) were isolated using fresh whole human blood from healthy donors as the starting material as previously described (Baugh and Travis 1976). Human genomic DNA was isolated from peripheral blood neutrophils of healthy donors as previously described (Skrzeczynska-Moncznik and others 2012). LL37 was obtained from ProImmune. CpG (ODN2216) and DNaseI were purchased from Invivogen and Invitrogen, respectively.
Patients
All of the human studies were performed in compliance with ethical protocols KBET/44/B/2011 and KBET/87/B/2014 approved by the Jagiellonian University Institutional Bioethics Committee. Declaration of Helsinki protocols were followed. All participants provided their written informed consent to participate in these studies as recommended by the ethics board. In total, 6 psoriasis patients (age 27.7 ± 6.6 years; M:F 5:1) and 63 healthy individuals (age 32 ± 7.7 years; M:F 56:7) were enrolled in these studies. The severity of the psoriatic skin lesions was assessed according to the Psoriasis Area Severity Index score (minimum, 0 points; maximum, 72 points) and ranged from 13.6 to 28 (mean ± standard deviation 18.2 ± 5.3).
Isolation of pDCs, granulocytes, and eosinophils
Human blood was collected, and peripheral blood mononuclear cells (PBMC) and the granulocyte-enriched fractions were harvested using Pancoll (PAN Biotech) gradient separation as described by the manufacturer. Purified pDCs were prepared from PBMC by negative selection (Miltenyi Biotech) according to the manufacturer's recommendations. The purity of the pDCs was ∼95% as determined by CD123 and CD303 immunoreactivity and flow cytometry analysis. The eosinophils were purified from the granulocyte-enriched fraction. This fraction was mixed with a 1% solution of polyvinyl alcohol (Sigma-Aldrich) in phosphate-buffered saline (PBS) and was incubated for 20 min at room temperature. The granulocytes were harvested from the upper phase and were subjected to hypotonic lysis to remove the contaminating red blood cells. The eosinophils were then purified by negative selection (Miltenyi Biotech) according to the manufacturer's recommendations. The purity of the isolated cells was examined by flow cytometry based on forward scatter and side scatter parameters and low CD16 immunoreactivity. The eosinophil preparations were routinely ∼95% pure.
pDC treatment
A total of 5 × 104 purified pDCs were seeded into round-bottomed 96-well plates in 50–100 μL RPMI. HNE, SLPI, and DNA; CatG, SLPI, and DNA; or LL37 and DNA were incubated for 15 min at room temperature, and the indicated mix was added to the pDC cultures at the following final concentrations: HNE, CatG, and SLPI, 1 μM; LL37, 10 μM, human neutrophil-derived DNA, 2 μg/mL. Alternatively, CpG at a final concentration 0.15 μM was added to the pDCs. The pDCs were then cultured at 37°C with 5% CO2 for 24 h in RPMI supplemented with 10% fetal bovine serum (FBS) and human recombinant IL-3 (50 ng/mL; Peprotech), which improves pDC survival in vitro. Where indicated, the pDCs were cultured for 24 h in the presence of eosinophil-conditioned media that constituted 1/4 of the final culture volume. To produce eosinophil-conditioned media, 5 × 104 purified eosinophils were cultured for 24 h in RPMI. The cells were then removed by centrifugation at 500 g for 5 min, and the supernatants were stored at −20°C until they were used. For the coculture experiments, the pDCs were plated with the indicated cell type at a 1:1 ratio or at the indicated eosinophil ratio, and the cells were grown in physical contact in the same well for 24 h. Alternatively, the pDCs and eosinophils were grown separated by a membrane. In this coculture system, the pDCs were plated at a density of 3 × 105 cells per well on the bottom well in the presence or absence of the indicated factors, whereas the eosinophils were plated at a similar density on semipermeable transwell inserts (6.5-mm diameter, 0.4 μm pore size; Falcon Transwell-Clear supports; BD Bioscience) and were cultured for 24 h. The conditioned media were then collected and centrifuged at 500 g for 5 min to remove the cells and were then tested by enzyme-linked immunosorbent assay (ELISA).
Real-time polymerase chain reaction
Cells were lysed with Fenozol Plus (AABiotechnology) after 3–24 h of culture. Total RNA was isolated with the Total RNA Zol-out kit (AABiotechnology) according to the manufacturer's instruction and converted to cDNA using MMLV reverse transcriptase (Lucigen) with oligo-dT primers (Genomed) and random hexamers (Invitrogen). Real-time polymerase chain reaction (PCR) was performed on the CFX96™ Real Time System (BioRad) using SYBR Green I containing universal PCR master mix (AABiotechnology) and the following primers specific for human IFNs and GAPDH (Genomed); IFNA1 (5′-CTTCAACCTCTTTACCACAAAAGATTC-3′ 5′-TGCTGGTAGAGTTCGGTGCA-3′), IFNA2 (5′-CTTGAAGGACAGACATGACTTTGGA-3′ 5′-GGATGGTTTCAGCCTTTTGGA-3′), IFNA6 (5′-TCCATGAGGTGATTCAGCAGAC-3′ 5′-GCTGCTGGTAAAGTTCAGTATAGAGT-3′), IFNA8 (5′-CCTTCTAGATGAATTCTACATCGAACT-3′ 5′-ACTCTATCACCCCCACTTCCTG-3′), IFNB (5′-TTGACATCCCTGAGGAGATTAAGC-3′ 5′-TTAGCCAGGAGGTTCTCAACAATAG-3′), and GAPDH 5′-GAGTCAACGGATTTGGTCGTATTG-3′, 5′-ATGTAGTTGAGGTCAATGAAGGGG-3′). The relative gene expression normalized to GAPDH was calculated using the 2−ΔΔCT method (Livak and Schmittgen 2001).
Western blot analysis
Purified eosinophils were cultured alone or together with a similar number of pDCs in RPMI for indicated time. The cells were lysed with 0.2% Triton X100, and the conditioned media were collected and centrifuged as described above. The samples were then resolved using 16% sodium dodecyl sulfate–polyacrylamide gel electrophoresis under reducing conditions. ECP was visualized by enhanced chemiluminescence after incubation with rabbit anti-human ECP antibodies (Abcam) and goat anti-rabbit immunoglobulin G (IgG) antibodies conjugated to horseradish peroxidase (Sigma-Aldrich).
Flow cytometry
The pDCs were stained with the following directly conjugated monoclonal antibodies: PE-anti-CD80 (clone 2D10.4, IgG1) and PE-anti-CD86 (clone IT2.2), which were both from eBioscience; or mouse isotype controls PE-IgG1 (clone MOPC-21) and PE-IgG2b (clone MPC11), which were both from BioLegend. The pDC viability was monitored by 7AAD and AnnexinV (BD Bioscience) staining. Flow cytometry was performed on an LSRII (BD Biosciences), and the data were analyzed using the DIVA and FCS Express software.
Enzyme-linked immunosorbent assay
Expression of IFN proteins was measured by ELISA; IFNα, (IFNα2 subtype; eBioscience) and IFNλ (IFNλ1 subtype; R&D Systems) according to the manufacturer's recommendations.
Immunohistochemistry
Frozen 6-μm sections were prepared from the skin biopsies. The sections were fixed in methanol and stained with May-Grunwald and Giemsa. Alternatively, the sections were fixed in acetone, blocked with goat IgG, and were stained with rabbit anti-human ECP (Abcam), rabbit anti-human HNE (Athens Research and Technology), or rabbit IgG (Sigma-Aldrich) as a negative control followed by allophycocyanine-goat anti-rabbit IgG F(ab)2 (Jackson ImmunoResearch). The sections were counterstained with Hoechst 33258 (Invitrogen). The isolated eosinophils were seeded on coverslips, fixed with 3.7% formaldehyde for 10 min, and were blocked overnight with 5% FBS, 1% bovine serum albumin, 0.05% Tween, 2 mM ethylenediaminetetraaceticacid (EDTA) in PBS at 4°C. The cells were then stained for ECP in the presence of 0.1% saponin as described above. The images were captured with a fully motorized fluorescence microscope (NIKON; Eclipse) and were analyzed by NIS elements software (Nikon).
DNA degradation
Human neutrophil DNA (4 μg) was incubated with 10 μL of eosinophil-conditioned media for 24 h at 37°C. As a positive control, the DNA was incubated with DNAseI (100–200 U/mL) for 15 min. To stop the enzymatic activity, 5 mM EDTA was added. To analyze DNA degradation, the samples were run on a 1% agarose gel in the presence of ethidium bromide.
Results
We initially tested if the skin of individuals suffering from psoriasis is infiltrated with eosinophils. Immunohistochemical examination of affected skin revealed the presence of scattered cells with the appearance of eosinophils by the May-Grunwald and Giemsa staining (Fig. 1A), and ECP-positive cells (Fig. 1B, D) were present in the skin of psoriasis patients. We also noted ECP-positive cells in close proximity to NETs (Fig. 1B, C) and NETs decorated with SLPI (Fig. 1D, E). Eosinophils that are found in the same skin region as NETs may play a regulatory role in the NET-mediated stimulation of IFNI expression in pDCs. To test whether eosinophils affect the ability of pDCs to secrete IFNI, we stimulated pDCs purified from the blood of healthy donors with a mixture of HNE, neutrophil DNA, and SLPI (HNE/DNA/SLPI) in the presence or absence of eosinophils isolated from autologous donors. We previously reported that SLPI- and HNE-decorated DNA deposits are potent stimulators of IFNα production in pDCs (Skrzeczynska-Moncznik and others 2012, 2013). However, in contrast to IFNα, IFN type III such as IFNλ1was not produced in response to the HNE/DNA/SLPI complexes (data not shown), suggesting that this stimulus primarily induces IFNI or more specifically IFNα in pDCs. As shown in Fig. 2, eosinophils on their own did not produce detectable amounts of IFNα, and the addition of eosinophils to pDCs did not significantly alter IFNα levels in the conditioned media. However, when pDCs were activated with the HNE/DNA/SLPI complexes to produce IFNα, the presence of a comparable number of eosinophils in the cocultures significantly inhibited production of IFNα in pDCs. Eosinophil treatment resulted in an up to 10-fold reduction in IFNα production (Fig. 2A) with an efficiency proportional to the eosinophil/pDC ratio (Fig. 2B) without significantly altering pDC viability as determined by 7ADD and AnnexinV staining and flow cytometry (Fig. 2C, D). Moreover, the observed effects were specific for eosinophils since a similar number of granulocytes or PBMCs did not significantly block the production of IFNα in pDCs (Fig. 2A). Collectively, these data suggest that eosinophils selectively suppressed the production of IFNα in the HNE/DNA/SLPI-activated pDCs.
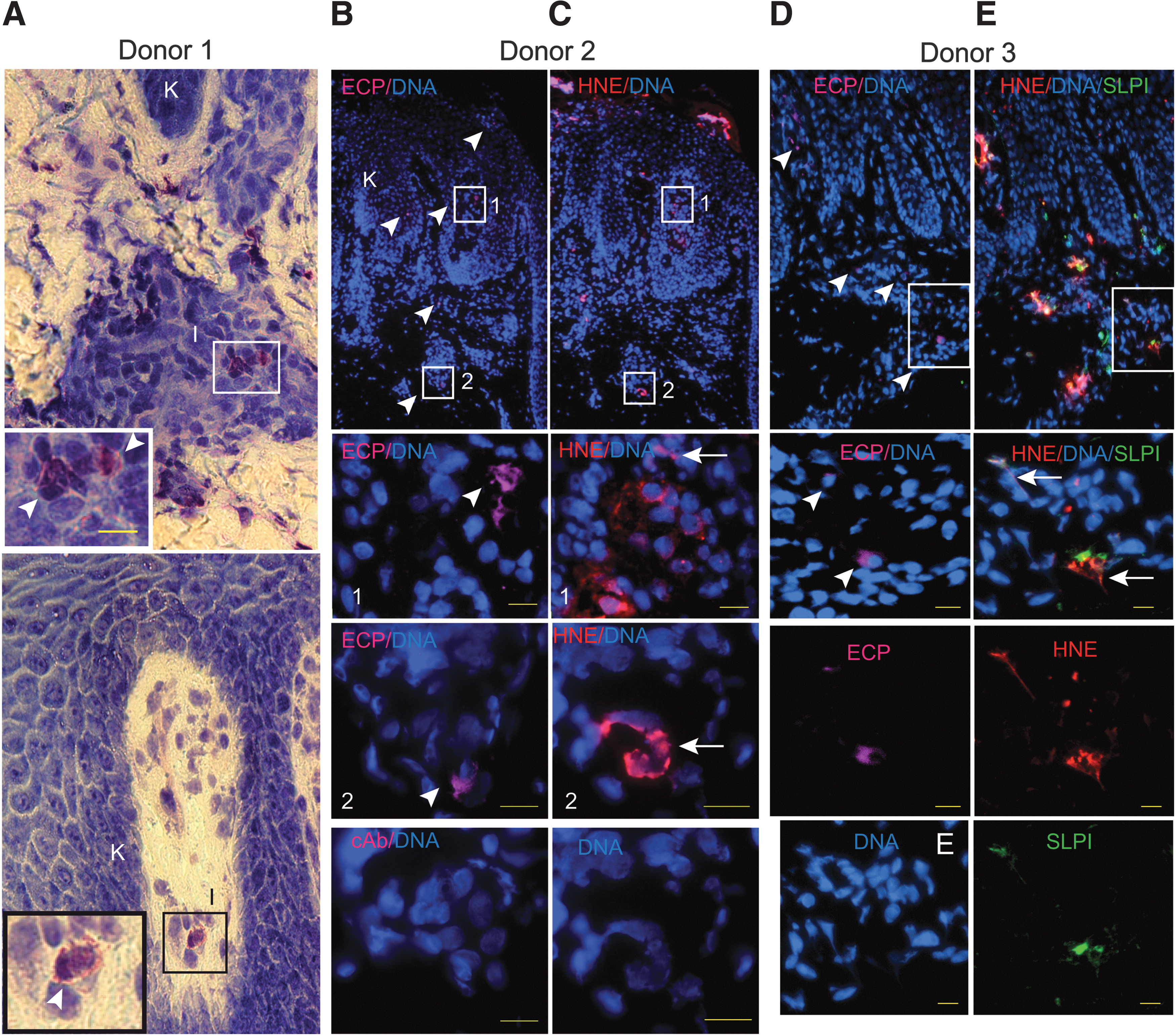
Eosinophils infiltrate lesional skin of psoriasis patients. Light micrographs of psoriatic skin stained with May-Grunwald and Giemsa, donor 1
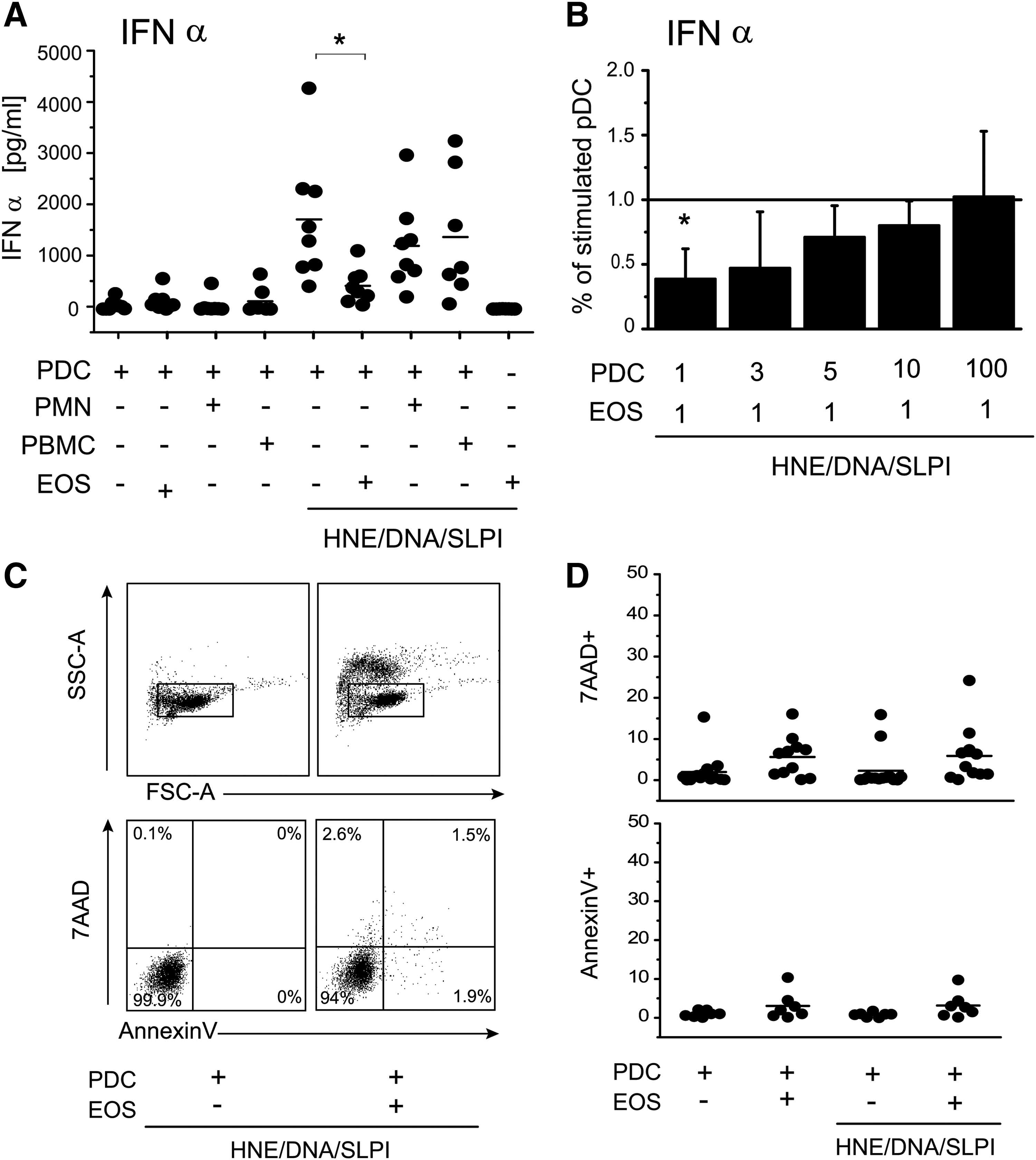
Activated pDCs produce less IFNα in response to coculture with eosinophils. The purified pDCs were cultured alone or together with the indicated, autologous cells at a 1:1 ratio
To determine whether the cultured eosinophils exhibit a degranulated phenotype, we analyzed the expression pattern of ECP in the cultured cells by immunohistochemistry and the levels of ECP in conditioned media by Western blot analysis. As shown in Fig. 3A, in contrast to the freshly isolated eosinophils that exhibited strong intracellular localization of ECP, eosinophils cultured for 24 h were found to stain for ECP mainly at the cell surface, suggesting degranulation and release of ECP from the cells. These findings were corroborated by the presence of ECP in the conditioned media of the cultured eosinophils or the eosinophils incubated with pDCs (Fig. 3B). Together, these data suggest that degranulated eosinophils are responsible for attenuating IFNα release in pDCs.

Cultured eosinophils undergo degranulation as evidenced by the release of ECP. Purified eosinophils were incubated in RPMI for the indicated times and were then stained for ECP (red) and DNA (blue)
To determine whether physical pDC–eosinophil contact was required for inhibition of IFNα production, we cultured pDCs and eosinophils in separate chambers in transwell plates. In this coculture system, pDCs and eosinophils were separated by a 0.4 μm semipermeable membrane allowing only for a cell–cell contact via soluble factors. Blocking the direct interaction between the HNE/DNA/SLPI-stimulated pDCs and eosinophils resulted in a similar level of IFNα production as in the nonseparated cells (Fig. 4A), which is indicative of contact-independent inhibition. Moreover, a statistically significant decrease (up to 5-fold reduction) in IFNα production was observed in pDCs stimulated with the HNE/DNA/SLPI complex in the presence of eosinophil-conditioned media (Fig. 4B). Together, these findings suggest that eosinophils control the extent of IFNα expression in the activated pDCs through soluble factors.
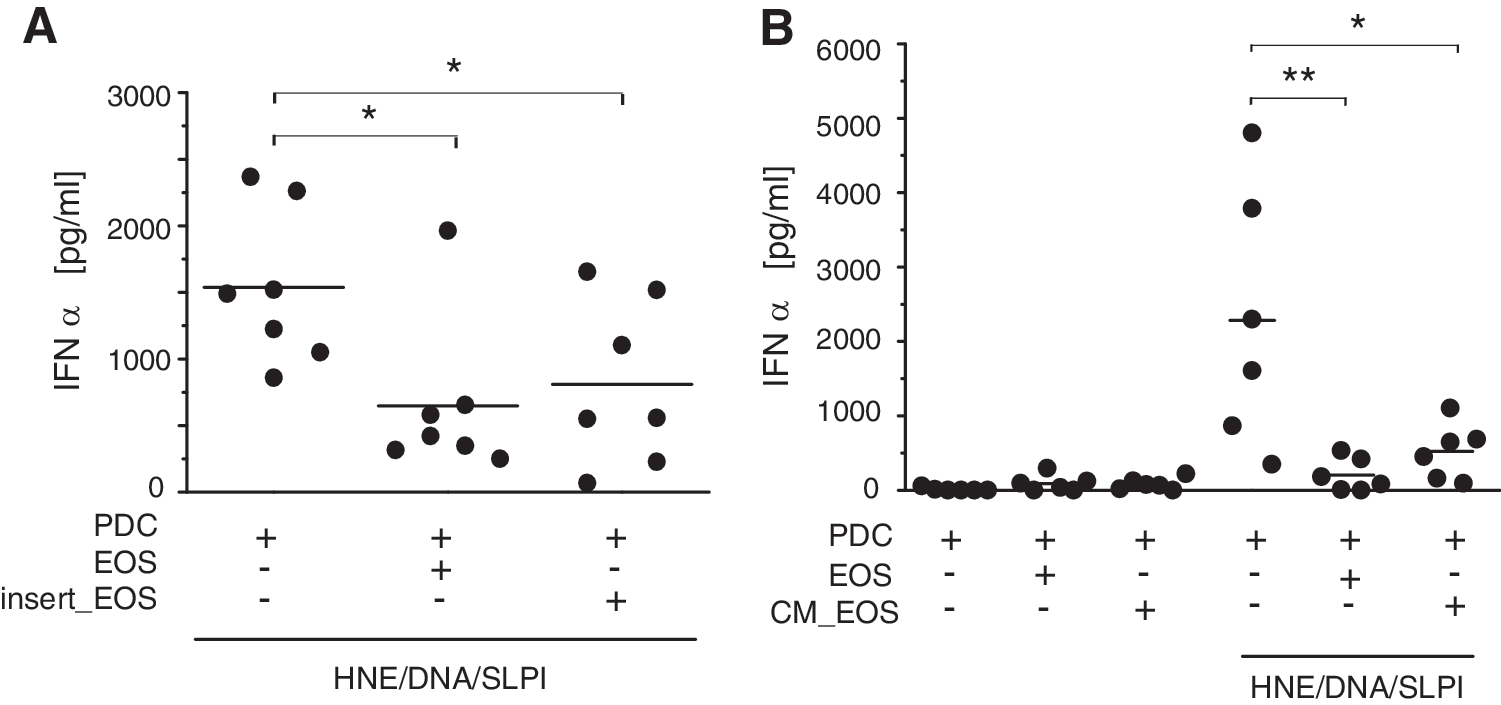
Eosinophils attenuate the pDC response through soluble factors. The purified pDCs were stimulated for 24 h with the HNE/DNA/SLPI complexes in the absence or presence of eosinophils (EOS) at a 1:1 ratio. Where indicated, the pDCs and eosinophils were physically separated with a semipermeable membrane (insert_EOS)
To explore whether eosinophils drive changes in IFNα transcription, and whether expression of other IFNIs is affected by eosinophils, qPCR analysis was first used to measure mRNA levels for IFNA subtypes and for IFNB in the pDCs stimulated with the HNE/DNA/SLPI mixture for 3, 4, 6, 18, and 24 h. Among 4 tested genes encoding IFNα, IFNA1, IFNA2, IFNA6, and INFA8, the highest induction (∼120-fold) was noted for IFNA1 and IFNA2 and the lowest for IFNA6 (∼25-fold) in response to the HNE/DNA/SLPI complexes at 4 or 6 h, whereas IFNB expression was induced on average 90-fold compared with untreated pDCs (Fig. 5A B and data not shown). Therefore, in subsequent experiments, we determined an impact of the eosinophil-conditioned media on mRNA levels for IFNA2 and IFNB in the HNE/DNA/SLPI-activated pDCs at 6 h. Of note, IFNA2 gene product IFNα2 was the IFNα subtype primarily detected by ELISA (Figs. 2 and 4). pDCs treated with the eosinophil-conditioned media for 6 h displayed significantly reduced IFNA2 expression (Fig. 5C), in addition to showing a decrease in IFNα protein levels at 24 h (Fig. 4). In contrast, IFNB transcript levels were not reduced by eosinophils (Fig. 5D). We conclude that a response of the HNE/DNA/SLPI-activated pDCs to eosinophil-derived factors was essentially limited to IFNα. Moreover, eosinophil-dependent inhibition of both gene and protein IFNα expression suggests that eosinophils suppress production of IFNα, at least partially, at the mRNA level.

IFNA2 but not IFNB mRNA levels are decreased by the eosinophil-conditioned media in the activated pDCs. The purified pDCs were stimulated with the HNE/DNA/SLPI complexes for indicated times
HNE/DNA/SLPI complexes stimulate pDCs largely in a TLR9-dependent manner, which implicates DNA as a major activating component in the complexes (Skrzeczynska-Moncznik and others 2012). To determine whether eosinophils attenuate the ability of the HNE/DNA/SLPI-stimulated pDCs to produce IFNα through releasing DNA-degrading factor(s), we incubated neutrophil-derived DNA with the eosinophil supernatants. As demonstrated in Fig. 6A, the conditioned media from the eosinophils alone or the cocultures of eosinophils with pDCs did not significantly degrade the DNA, suggesting that neither eosinophils nor eosinophils that were in contact with pDCs secreted factors with DNAse-like activity. Moreover, the levels of IFNα produced in the pDCs in response to stimulation with CpG, the well-known short synthetic single-stranded DNA TLR9 agonist, did not differ in the presence of eosinophils (Fig. 6B). Together, these data indicate that the inhibitory effect of eosinophils on the production of IFNα in pDCs is not likely to be DNA dependent.
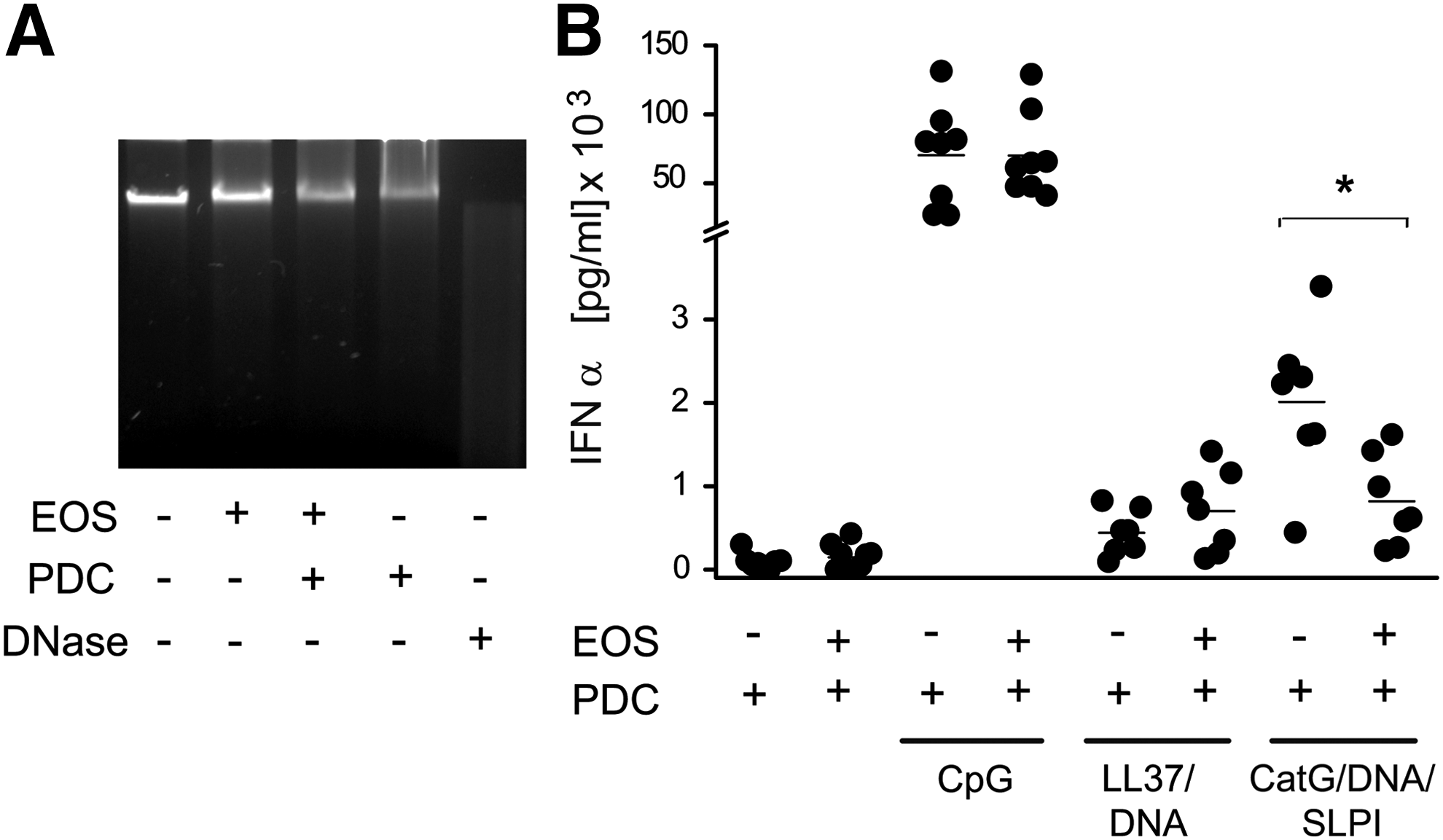
Eosinophils do not release DNAse-like activity and do not inhibit IFNα production in the CpG- or LL37/DNA-stimulated pDCs. The neutrophil-derived DNA was treated with conditioned media from the indicated cells for 24 h or with DNAse (positive control). The samples were then run on agarose gels and were visualized by ethidium bromide staining. The data are from 1 experiment and are representative of 3 experiments
To determine whether inhibition of IFNα synthesis by eosinophils is specific for pDCs stimulated with the HNE/DNA/SLPI complexes, we substituted these complexes with complexes containing human neutrophil DNA and LL37 (LL37/DNA). Alternatively, human neutrophil DNA together with SLPI and CatG (CatG/DNA/SLPI) instead of HNE was also used as a stimulating agent. Similarly to HNE, neutrophil protease-CatG together with DNA and SLPI markedly stimulates pDCs to secrete IFNα (Skrzeczynska-Moncznik and others 2013). As shown in Fig. 6B, production of IFNα by pDCs treated with the CatG/DNA/SLPI complexes was significantly inhibited by eosinophils, in contrast to the LL37/DNA-stimulated pDCs that were not affected by eosinophils. These data suggest that eosinophils exhibit inhibitory potential toward pDCs in the stimulus-dependent manner.
The pDCs in lesional skin of psoriatic patients display an activated phenotype that manifests not only in the production of IFNα but also in the upregulated surface expression of costimulatory molecules, including CD86 and CD80 (Nestle and others 2005). Therefore, we next determined whether the effect of eosinophils on the HNE/DNA/SLPI-stimulated pDCs is restricted to the inhibition of IFNα synthesis or is accompanied by a change in pDC maturation. As shown in Fig. 7, the pDCs cultured in the presence of eosinophil-conditioned media were not significantly altered in their maturation level as assessed by both the percentage of cells expressing CD80 and CD86 (Fig. 7A) and by the mean fluorescence intensity (Fig. 7B). Collectively, these data suggest that eosinophil-derived soluble factor(s) affect pDC function primarily through inhibition of IFNα release.
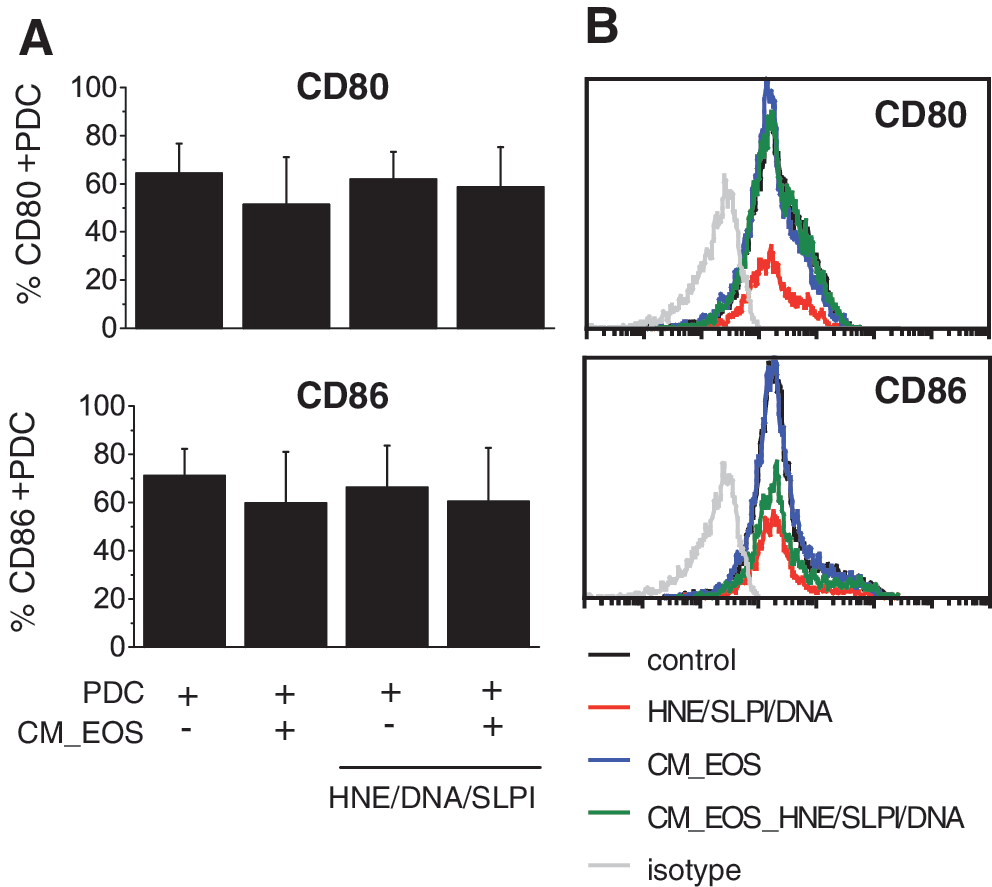
Eosinophils do not alter costimulatory molecule expression on pDCs. The purified pDCs were cultured alone or were stimulated with the HNE/DNA/SLPI complexes in the absence or presence of eosinophil-conditioned media (CM_EOS). After 24 h, the cells were characterized for CD80 and CD86 expression by flow cytometry analysis. The cumulative frequency data
Discussion
In this study, we identify a previously uncharacterized eosinophil activity associated with the regulation of pDC function in autoinflammatory responses. PDCs and their hallmark product, IFNα, were reported to play a crucial role in autoimmunity (Nestle and others 2005; Cichy and Zabel 2011; Garcia-Romo and others 2011; Lande and others 2011). Thus, eosinophils may serve to limit the deleterious effects of this cytokine during inflammatory responses through the downregulation of IFNα levels. In contrast to neutrophils, intact eosinophils were not found in high numbers in psoriatic skin, but they were present near NETs, suggesting that they are well positioned to regulate NET-mediated signaling. Neutrophils accumulate in the skin of psoriatic patients at sites that are also infiltrated by pDCs, and the components of NETs mobilize pDC responses (Majewski and others 2016). Together, these data suggest that eosinophils may control IFNα production in the pDCs that infiltrate psoriatic skin.
Eosinophils appear to preferentially target the ability of the HNE/DNA/SLPI-activated pDCs to express IFNα, but not IFNβ and without promoting the maturation of these cells, which is required for pDC-mediated activation of T cells (Liu 2005). These findings suggest that eosinophils may selectively control IFNα-mediated responses and likely do not control the initiation of adaptive immune responses by differentiated pDCs. Moreover, in contrast to the HNE/DNA/SLPI- or the CatG/DNA/SLPI-activated pDCs, the production of IFNα in the LL37/DNA- or the CpG-stimulated pDCs was not inhibited by eosinophils, suggesting that eosinophils affect pDC activation in a stimulus-dependent manner. The potential pathophysiological relevance of eosinophils, DNA deposits decorated with SLPI and HNE or CatG, and pDCs, is supported by partially overlapping distribution of these cells and structures in psoriasis skin (Fig. 1; Skrzeczynska-Moncznik and others 2012, 2013).
While eosinophils have long been associated with allergic diseases, they have also been linked to autoinflammatory conditions. One of the most well-documented involvements of eosinophils in autoinflammatory disorders relates to inflammatory bowel diseases (IBD). Activated eosinophils accumulate in the gut, and the elevated levels of eosinophils correlate with severity of IBD, but mice depleted of eosinophils develop less severe disease (Jung and Rothenberg 2014). In addition to the gastrointestinal tract where eosinophils are frequently found under both homeostatic and inflammatory conditions, these cells also infiltrate other organs, including inflamed skin in allergic dermatitis and to a lesser extent in psoriasis. Interestingly, some individuals are affected by both psoriasis and allergic dermatitis and displayed a mixed psoriasis and eczema skin phenotype by histology (Quaranta and others 2014a, 2014b). Therefore, patients suffering from both disorders might be particularly prone to eosinophil-mediated regulation of inflammatory responses in skin due to a higher number of skin infiltrating eosinophils. Support for a potential skin eosinophil involvement in psoriasis comes from recent findings that human eosinophils constitutively express receptors for IL17 and IL23 and respond to IL17A, IL17F, and IL23 with the release of specific chemokines (Cheung and others 2008). As IL17 and IL23 are considered key drivers of skin pathology in psoriasis, these data suggest that eosinophils are likely to respond to these cytokines in psoriatic skin through the release of regulatory mediators.
Our data indicate that conditioned media from eosinophils were sufficient to inhibit pDC responses in vitro, suggesting that factors that were released from eosinophils were responsible for the inhibitory effect. We detected ECP in the conditioned media of cultured eosinophils, suggesting that eosinophils are likely to release pDC-regulating factors on activation. Under in vivo conditions, the potential of eosinophils to control the production of IFNα in pDCs might be facilitated by the stimulation of eosinophils by inflammatory mediators that are abundant in psoriatic skin.
Considering that eosinophils can produce an array of nucleases, such as the RNAses ECP and EDN, we examined whether these cells inhibit the synthesis of IFNα by targeting DNA in the HNE/DNA/SLPI stimulatory complex. Although the inhibitory factor(s) responsible for tailoring HNE/DNA/SLPI-stimulated pDC responses by eosinophils remains unknown, our data suggest that eosinophils are unlikely to act through the degradation of DNA as they did not release factors with DNAse-like activity. Moreover, the inhibitory factor(s) does not appear to specifically target DNA in the stimulatory HNE/DNA/SLPI complex because, in contrast to the HNE/DNA/SLPI-activated pDCs, the production of IFNα in the CpG-stimulated pDCs was not inhibited by eosinophils.
In summary, our findings indicate a potential role of eosinophils in shaping pDC–neutrophil interactions that translate to the downregulation of IFNα production by pDCs. The eosinophil-dependent IFNα control in pDCs may contribute to constraining the NET-mediated excessive inflammatory responses.
Footnotes
Acknowledgments
This work was supported, in part, by Polish National Science Center grant 2011/02/A/NZ5/00337 and a grant from the Foundation for Polish Science TEAM/2010-5 and was cofinanced by the European Union within European Regional Development Fund Award (to J.C.), Polish National Science Centre grant UMO-2011/01/B/NZ5/00354 (to J.S.-M.), and DS6. The Faculty of Biochemistry, Biophysics, and Biotechnology of the Jagiellonian University is a beneficiary of the structural funds from the European Union (grant No: POIG.02.01.00-12-064/08) and a partner of the Leading National Research Center (KNOW) supported by the Polish Ministry of Science and Higher Education.
Author Disclosure Statement
No competing financial interests exist.