
Abstract
Type III interferons (IFN-λ1/λ2/λ3, also known as IL-29/28A/28B, and IFN-λ4) are a recently discovered interferon group. In this study, we first identified the Chinese goose IFN-λ (goIFN-λ). The full-length sequence of goIFN-λ was found to be 823 bp. There was only one open reading frame that contained 570 bp, and, encoded 189 amino acids. The predicted goIFN-λ protein showed 78%, 67%, and 40% amino acid identity with duIFN-λ, chIFN-λ, and hIFN-λ3, respectively. The tissue distribution of goIFN-λ existed as a parallel distribution with goIFNLR1 as its functional receptor, which was mainly expressed in epithelium-rich tissues, such as lung, gizzard, proventriculus, skin and pancreas, and immune tissues, such as harderian gland and thymus. Furthermore, the immunological characteristics studies of goIFN-λ showed that there was a significant increase in the mRNA at the transcriptional level of goIFN-λ after the peripheral blood mononuclear cells were stimulated with ploy (I:C) and ODN2006, and infected with Gosling plague virus (GPV). In vivo, the mRNA transcriptional level of goIFN-λ increased nearly 20 times in the lung tissue and nearly 40 times in the pancreatic tissue after being artificially infected with H9N2 AIV. It is suggested that goIFN-λ might play a pivotal role in the mucosal immune protection and antiviral defense.
Introduction
F
Different from type I interferon receptor, that has a ubiquitous expression, type III interferon cognate receptor subunit IFNLR1 has a much restricted expression and is primarily distributed in certain cells of epithelial origin (Kotenko and others 2003; Sheppard and others 2003; Doyle and others 2006; Lasfar and others 2006). Several investigations have shown that, IFN-λ also has limited responses, which closely depend on the expression of its cognate receptor IFNLR1 (Zhou and others 2007). In human and mouse cell lines, IFN-λ response is observed primarily in epithelial specificity, such as keratinocytes, intestine, airway epithelial cells, and pDCs (Lasfar and others 2006; Megjugorac and others 2009; Pott and others 2011; Ioannidis and others 2012; Yin and others 2012), while no response is seen in fibroblasts, splenocytes, and endothelial cells (Lasfar and others 2006; Sommereyns and others 2008).
Compared to human type III IFNs, the Type III IFNs of other vertebrates or nonmammalian species were gradually identified. However, there was not much known about the immunological regulation and antiviral function of IFN-λs. Chicken IFN-λ (chIFN-λ) and duck IFN-λ (duIFN-λ) were identified in years 2008 and 2014, respectively (Karpala and others 2008; Yao and others 2014). The protein structure of chIFN-λ has five exonic regions, which consist of 186 amino acids (aa), and has 36% identity with huIFN-λ2 (Karpala and others 2008). It is shown that chIFN-λ might play a role in defending mucosal surfaces against viral intruders (Reuter and others 2014). The duIFN-λ cDNA encodes a 185 amino acid protein, which has 63% and 38% identity with chIFN-λ and huIFN-λ3, respectively (Yao and others 2014). In our previous research, the receptor of goose Type III IFN (goIFNLR1) had been identified, and it indeed has a cell type-specific expression profile, primarily expressed in epithelia-related cells (Zhou and others 2015).
In this article, we performed cloning and sequencing of the Chinese goose IFN-λ (goIFN-λ) cDNA for the first time. Furthermore, the tissue distribution and the immunological characteristics of goIFN-λ were studied. Identification of goIFN-λ will shed light on the antiviral immune response of aquatic birds and facilitate further study on its antiviral defense research.
Materials and Methods
Animals and reagents
One-day-old and 2-week-old goslings and adult (beyond 1 year) Chinese geese (Anser cygnoides) were purchased from the farm of Sichuan Agricultural University (Yaan, Sichuan Province). The animal studies were approved by the Institutional Animal Care and Use Committee of Sichuan Agricultural University and followed the National Institutes of Health guidelines for the performance of animal experiments. To detect tissue distribution of goIFN-λ, 20 tissues including small intestine (SI), cecum (Ce), rectum (Re), lung (Lu), skin (SK), proventriculus (Pr), spleen (SP), cecal tonsil (CT), bursa of Fabricius (BF), harderian gland (HG), liver (Li), kidney (K), heart (H), brain (B), gizzard (Gi), thymus (T), muscle (M), trachea (Tr), pancreas (P) and blood were collected.
Viruses
Low pathogenic avian influenza (LPAI) virus H9 N2 was provided by Shanghai Veterinary Research Institute, Chinese Academy of Agricultural Sciences. Duck tembusu virus (DTMUV) and gosling plague virus (GPV) were provided by our laboratory. In this assay, the genomic copies of H9 N2 AIV was 7.12 × 1012.64 copies/100 μL; GPV titer determined as median egg infectious dose (EID50) on goose embryo was 10A−6.6 EID50/200 μL and the infectious dose of DTMUV was 6.3 × 106 TCID50/100 μL.
Gene cloning of goose IFN-λ
Based on the conserved domains from the duck IFN-λ sequence (GenBank accession number: NM_001310792.1) and the chicken IFN-λ sequence (GenBank accession number: EF587763.1), degenerate primer goIFNL-F1 and goIFNL-R1 (Table 1) were designed to amplify the partial cDNA fragment of goIFN-λ from the goose lung tissue template with the polymerase chain reaction (PCR). The PCR volume is 0.2 μL cDNA, 0.2 μL of each primer, 5 μL 2 × Taq Mix polymerase (Tiangen Biotech (Beijing) CO., LTD), and ddH2O for complement in total 10 μL volume. Then, using the conventional 5′ and 3′ rapid amplification of cDNA ends, (RACE) PCR was performed to obtain full-length cDNA sequences of goIFN-λ as described before (all the primers listed in Table 1) (Zhou and others 2015). Finally, the PCR products were cloned into pMD-19T vector (TaKaRa Co., LTD) for sequence analyses. The primer goIFNL-F2 and goIFNL-R2 were used for amplifying the full goIFN-λ open reading frame (ORF) by using PrimeSTAR DNA polymerase (TaKaRa Co., LTD) to further confirm the sequence.
Quantitative real-time PCR analysis
Total RNA was extracted with RNA plus reagent (TaKaRa Co., LTD) according to the manufacturer's instructions and reverse transcribed into cDNA with 5× All-in-one RT Master Mix (Applied Biological Materials) according to manufacturer's protocol. qPCR was performed as described previously by using Bio Rad CFX 96 Real-Time qPCR instrument with goose GAPDH as the internal control (primers used are listed in Table 1). All reactions contained 0.4 μL cDNA from certain templates, 5 μL 2× EvaGreen Mix (Applied Biological Materials), 4 μL ddH2O, and 0.3 μL of each primer to a final volume of 10 μL. The relative gene expression was calculated using the mean values obtained with the arithmetic formula ΔΔCt.
Isolation and culture of goose peripheral blood mononuclear cells
Peripheral blood mononuclear cells (PBMC) were isolated using the Goose Lymphocyte Separation Medium kit (GLSMK) (TBD sciences) from adult goose. Twenty milliliters of blood was collected from the jugular veins of each of the three adult geese into sterile 50 mL centrifuge tubes, which contained heparin sodium (25 IU/mL). The blood was diluted 1:1 in sample diluent (TBD sciences), and 1:1 slow added the diluted sample above on the GLSM (ban mixed), centrifuged for 30 min at 800 g. After centrifugation, second ring of ivory PBMC was collected into a new 50 mL tube. Cells were washed thrice with an equipped cleaning fluid and centrifuged for 5 min at 250 g. Finally, the sediment of PBMC was resuspended in 1 mL RPMI 1640 medium (Gibco). Cells were counted with the microscope and treated with a complement culture medium with 10% fetal bovine serum (FBS), 100 μ/mL penicillin, and 100 μg/mL streptomycin at 37°C with 5% CO2. Cells were seeded at the density of 8 × 105 cells/mL.
Immunological characteristics research in vitro
Four kinds of TLR agonists were selected in this assay to stimulate the PBMCs. Poly (I:C) (30 μg/mL) is the agonist of TLR3, ODN2006 (25 μg/mL) is the agonist of TLR9, LPS (25 μg/mL) is the agonist of TLR4, and R848 is the agonist of TLR7/8 (5 μg/mL). Three kinds of poultry-related viruses, H9N2 AIV (goose origin), DTMUV, and GPV, mentioned above were also used to treat the cell models. Each of them received 50 μL. Cells with 50 μL of PBS were chosen as the negative controls. Six hours later, 3 identical wells of each group were collected for qRT-PCR detection of the mRNA expression level of gene goIFN-λ and goIFNLR1 according to the procedure described above.
The effects of viruses on goIFN-λ mRNA level in vivo
Goslings (3 days old) were selected and divided into four groups, randomly. Three individuals of each group received artificial infection with H9 N2 AIV (p.o. 250 μL and i.n. 250 μL), DTMUV (i.h. 500 μL), and GPV (p.o. 250 μL and i.m. 250 μL). Group receiving treatment with 0.9% NaCl (i.m. 500 μL) served as the control group. Five days later, 6 tissues CE, LU, P, SI, TR, and blood were obtained from each individual for the detection of goIFN-λ, and goIFNLR1 mRNA expression level was determined by qRT-PCR method as described above.
Sequence bioinformatics analysis
The potential ORF amino acid sequence of goIFN-λ was predicted using the ORF Finder (
Statistical analysis
The mRNA transcriptional level assays underwent statistical analysis. All data analyses were performed using GraphPad Prism 5 and presented as mean ± standard deviation (SD). A P-value less than 0.05 (*) was considered to indicate significant differences between groups and a P-value less than 0.01 (**) was considered highly significant differences.
Results
Identification and sequence analysis of goIFN-λ cDNA
Accumulating data have shown that IFN-λ was mainly produced in epithelia-related cells; with this in mind, goIFN-λ was first cloned by PCR amplification from goose lung tissue cDNA (GenBank accession number was KX377964). The full-length sequence of goIFN-λ is 823 bp. There is only one ORF that contained 570 bp, encoded 189 aa, and with a 21 aa putative signal peptide situated in the N-terminal (Fig. 1). The goIFN-λ protein has an amino acid identity of 78% with duIFN-λ and 67% with chIFN-λ, and the amino acid identity decreased to 40% with huIFN-λ3. The phylogenetic tree was constructed based on the amino acid sequences that further showed goIFN-λ clusters in avian IFN-λ and closet structural similarity to duIFN-λ and chIFN-λ (Fig. 2).

Nucleotides and deduced amino acid sequence of the goIFN-λ ORF. The numbers of nucleotides and amino acids (left) and predicted signal peptide of goIFN-λ (underlined). * represents the termination codon. ORF, open reading frame.
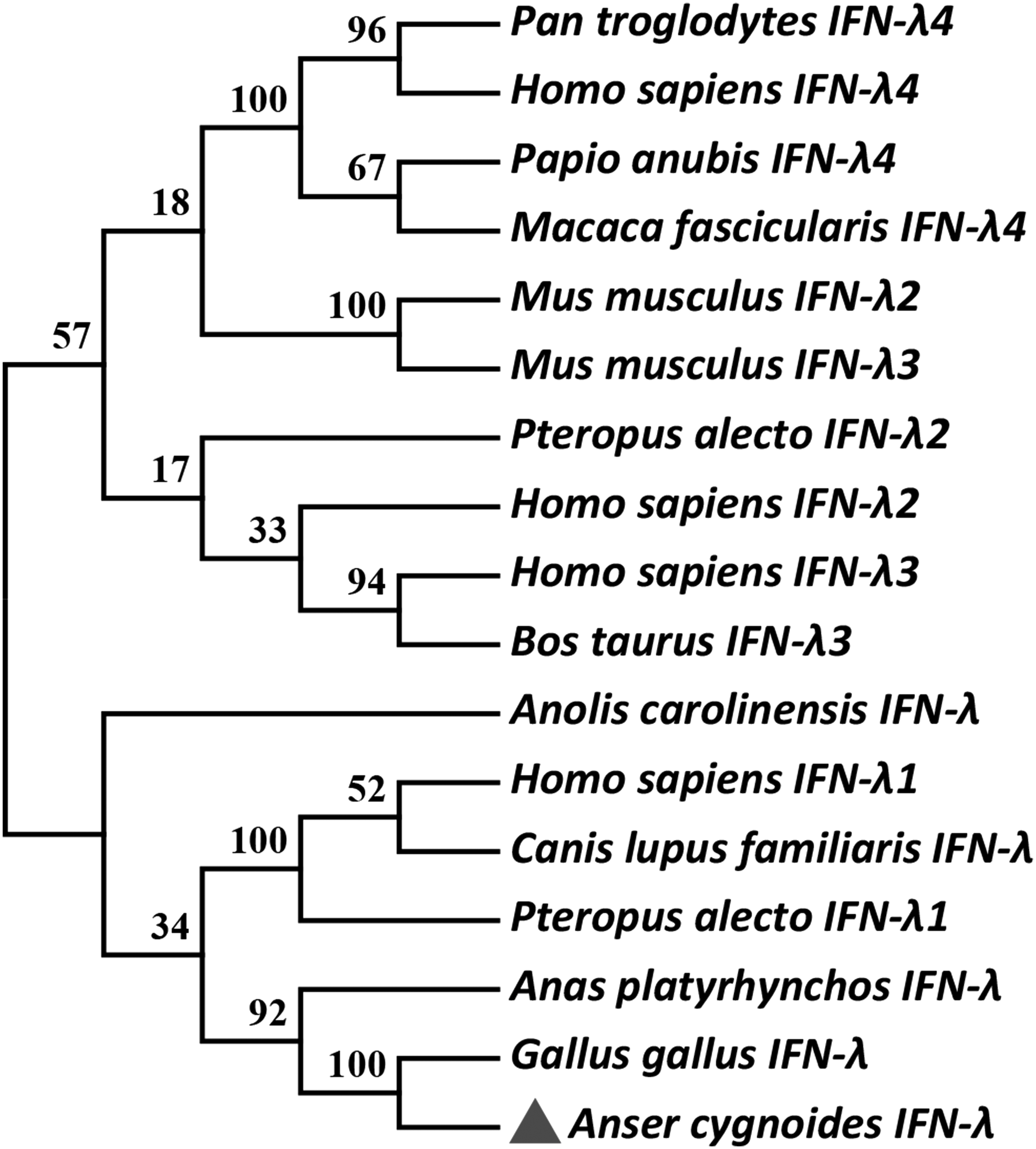
Phylogenetic tree based on amino acid sequence of goIFN-λ. Phylogenetic tree of IFN-λ proteins was generated with MEGA6.0 using the neighbor-joining method. Branches were validated by bootstrap analysis from 1,000 repetitions and are represented by numbers at the branch nodes. The GenBank IDs for IFN-λ amino acid sequences used for this study are as follows: Pan troglodytes IFN-λ4 (AFY99109.1); Homo sapiens IFN-λ4 (NP 001263183.2); Papio anubis IFN-λ4 (AGK38234.1); Macaca fascicularis IFN-λ4 (AGK38235.1); Mus musculus IFN-λ2 (AAX58714.1); Mus musculus IFN-λ3 (AAX58715.1); Pteropus alecto IFN-λ2 (AEF33949.1); Homo sapiens IFN-λ2 (AAI13584.1); Homo sapiens IFN-λ3 (AAI30315.1); Bos taurus IFN-λ3 (NP_001268830.1); Anolis carolinensis IFN-λ (AHY86484.1); Homo sapiens IFN-λ1 (AAI26184.1); Canis lupus familiaris IFN-λ (AGN92945.1); Pteropus alecto IFN-λ1 (AEF33950.1); Anas platyrhynchos IFN-λ (AII23257.1); Gallus gallus IFN-λ (ABU82742.1); and Anser cygnoides IFN-λ (KX377964). Triangle represents target sequence goIFN-λ.
Amino acid sequence analysis of goIFN-λ
Secondary structure prediction showed that the mature goIFN-λ protein has six α-helices (Helix A-Helix F, in magenta shadow of Fig. 3). It belongs to typical class II cytokines. The multiple amino acid sequence alignment of goIFN-λ with other IFN-λ sequences reported for mammalian species (Human, Mouse) and poultry species (Chicken, Duck) is illustrated in Fig. 3. It is shown that mature goIFN-λ protein contains four conserved cysteine residues (in green shadow); cysteine residues of hIFN-λ3 and hIFN-λ2 were an intramolecular disulfide bond site within the C-terminus. While some conserved residues (in gray shadow) were significant for IFNLR1 interaction and antiviral activity of hIFN-λ3, they were k36 hIFN-λ3, R53hIFN-λ3, L54 hIFN-λ3, Q100 hIFN-λ3, and F158 hIFN-λ3, respectively (Fig. 3).

Multiple alignment analysis of goIFN-λ amino acid sequence. The name of each species (left) and the site of amino acids (right). Predicted signal peptide of these genes (underlined), conserved cysteine residues (green shadow), and conserved residues (gray shadow) predicted as the binding site between IFN-λ and IFNLR1, and α-Helices (magenta). Amino acids conserved among all species are indicated as follows: “*”means identity, “:” means highly conserved, and “.” means weakly conserved.
Tissue distribution of goIFN-λ
Some of the currently available data show that IFN-λ has a much more restricted cellular distribution, which depends on the distribution of its cognate receptor (IFNLR1) (epithelial cell specificity). To test whether goIFN-λ also has this tissue specificity, quantitative analysis was performed for goIFN-λ mRNA detection. It is shown that goIFN-λ had a relatively high expression in the epithelia-related tissues (Fig. 4). In a 2-week-old gosling, a high mRNA expression level was found in the epithelia-related tissues of gizzard (GI), pancreas (P), and small intestine (SI), as well as the immune-related tissue (harderian gland, HG), while a bare expression in substantive organs, heart (H), brain (B), and muscle (M). In adult goose, the highest level of goIFN-λ was detected in the pancreas (P), while goIFN-λ was barely expressed in the muscle (M).

The tissues IFN-λ transcription profile of IFN-λ in gosling (2 W) and adult goose (Adult). Bar columns P (pancreas), LU (lung), SK (skin), GI (gizzard), PR (proventriculus), CU (rectum), CE (cecum), TR (trachea), LI (liver), K (kidney), and SI (small intestine) represent epithelia-related tissues (dark grey); bar columns T (thymus), BF (bursa of fabricius), HG (harderian gland), SP (spleen), Blood, and CT (cecum tonsil) represent immune tissues (black); and bar columns B (brain), H (heart), and M (muscle) represent substantive organs (light grey).
The effects of TLR ligands and viruses on the mRNA transcriptional level of goIFN-λ and goIFNLR1 in vitro
IFN-λs can be produced by virus infection and TLR ligands and subsequently induce the same Jak-STAT signal transduction pathway as type I IFNs. In this study, four kinds of agonists of different TLRs and three poultry-related virus were selected to treat goose PBMCs and after that, the mRNA transcriptional level of goIFN-λ and goIFNLR1 was detected. As shown in Fig. 5, the mRNA transcriptional level of goIFN-λ showed a significant increase after stimulating with ploy (I:C) and ODN2006, and treating with GPV in the PBMCs. While no changes of the goIFN-λ-mRNA transcriptional level were shown in R848-, LPS-, DTMUV-, and H9N2 AIV-treated cells. In addition, goIFNRL1 transcriptional level showed no significant change, neither by TLR agonists nor by poultry-related virus treatment.
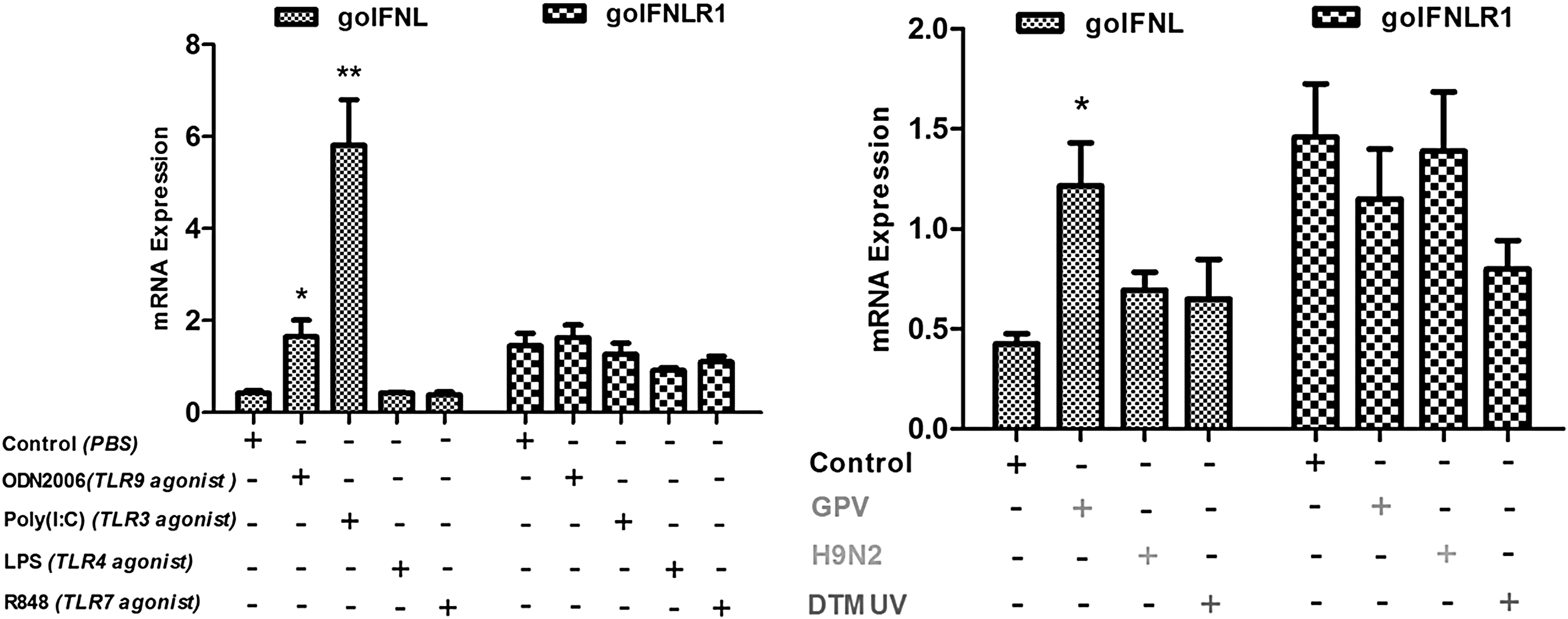
The effects of TLR ligands and viruses on the mRNA transcriptional level of goIFN-λ and goIFNLR1 in goose PBMCs. GAPDH was the control gene to quantify and qualify the transcription level of goIFN-λ and goIFNLR1 in goose PBMCs stimulated with ODN2006, poly (I:C), LPS, and R848 and treated with GPV, H9N2 AIV, and DTMUV. Data are represented as the mean ± SEM (n = 4) and the error bars indicate standard deviation. Cells treated with PBS were blank control, the difference between mRNA cytokine production in ligand-treated cells, and media-treated cells were tested using the unpaired, two-tailed t-test (*P < 0.05 and **P < 0.01). “+” represents cells treated with certain immune stimulant, “-” represents cells untreated with this immune stimulant. PBMC, peripheral blood mononuclear cell.
The effects of viruses on the mRNA transcriptional level of goIFN-λ and goIFNLR1 in vivo
To explore the goIFN-λ and goIFNLR1 expression level in virus-infected geese, 3-day-old goslings were chosen as animal models and infected with H9N2, GPV, and DTMUV, respectively. At 5 dpi, expression of goIFN-λ and goIFNLR1 mRNA in selected six tissues was analyzed by qRT-PCR. In our work (Fig. 6), both goIFN-λ and goIFNLR1 mRNA were constitutively expressed in five epithelia-related tissues (cecum, lung, pancreas, small intestine, and trachea) and one immune-related tissue (blood) of an uninfected goose. Under H9N2 virus infection, the high increase of goIFN-λ was observed in the pancreas, approximately 40-fold higher than control; the upregulation of goIFN-λ was detected in the lungs (20-fold changes). No change of goIFN-λ was seen in cecum, small intestine, trachea, and blood of H9N2 AIV-infected birds. However, expression of goIFN-λ mRNA has no significant change both in GPV- and DTMUV-infected birds relative to control birds in other selected tissues. In addition, no significant change in the goIFNLR1 mRNA transcriptional level was detected in these six selected tissues from the H9 N2 AIV-, GPV-, and DTMUV-infected geese.
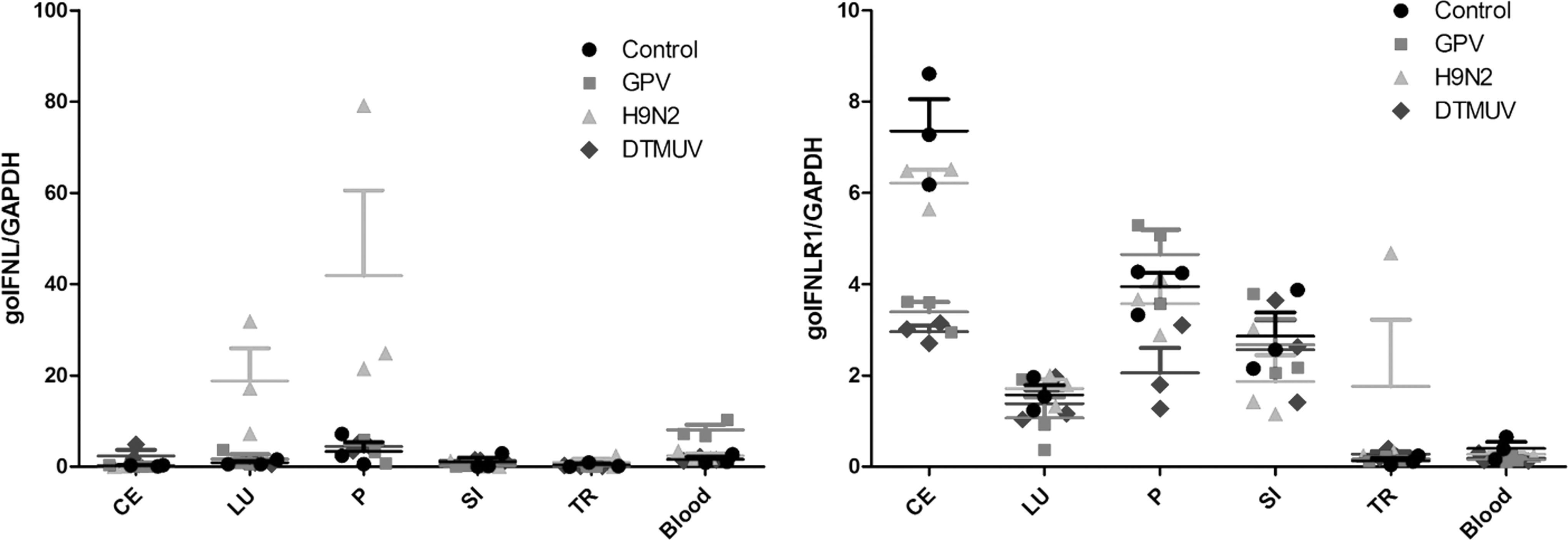
The effects of viruses on the mRNA transcriptional level of goIFN-λ and goIFNLR1 in vivo. GAPDH was the control gene to quantify and qualify the transcription level of goIFN-λ and goIFNLR1 in gosling tissues infected with GPV, H9N2 AIV, and TMUV, respectively. Six tissues were selected: cecum (CE), lung (LU), pancreas (P), small intestine (SI), trachea (TR), and blood. Negative control group (circle), GPV-infected group (square), H9N2 AIV-infected group (triangle), and TMUV-infected group (diamond). Data are represented as the mean ± SEM (n = 3) and error bars indicate standard deviation.
Discussion
This is the first report of the identification and biological characteristics of IFN-λ in the Chinese goose. The predicted goIFN-λ protein has an amino acid identity of 78%, 67%, and 40% with duIFN-λ, chIFN-λ, and hIFN-λ3, respectively. It is reported that the duIFN-λ gene is similar in size to chIFN-λ and hIFN-λ2/λ3, and the chIFN-λ gene is most similar to hIFN-λ2 (Karpala and others 2008; Yao and others 2014). In this study, the goIFN-λ gene is most similar to duIFN-λ and chIFN-λ. Unlike mammals, only one member of the IFN-λ family was predicted in the goose genome, which is also reported in chickens, ducks, and turkeys (Kaiser and others 2005; Karpala and others 2008; Dalloul and others 2010; Yao and others 2014). The phylogenetic tree analysis indicated that goIFN-λ belonged to poultry branch and was more similar to duIFN-λ and chIFN-λ compared with many other species. Interestingly, goIFN-λ perhaps belongs to interferon lambda-3-like.
The predicted goIFN-λ amino acid sequence analysis revealed that most likely, the signal peptidase cleavage site is located between 20 and 21 amino acid positions, which would yield a mature protein that has 168 aa and a signal peptide of 21 aa. The multiple amino acid sequence alignment shown by goIFN-λ has a signal peptide similar to other species' IFN-λ. Previous studies have demonstrated that hIFN-λ3 has two binding sites 1A and 1B, which react with its specific receptor IFNLR1 (Gad and others 2009; Zdanov 2010). Among those residues, R53 hIFN-λ3 and residue L54 hIFN-λ3 correspond to the binding site 1A, and the residue F158 hIFN-λ3 and residue K36 hIFN-λ3 correspond to the binding site 1B (Gad and others 2009). The residue Q100 hIFN-λ3 was in the vicinity site of interface with another receptor chain IL-10R2 (Gad and others 2009). In this work, all these available residues are mostly conserved between avian and other mammalian species. It is shown that residue F158 hIFN-λ3 plays a critical role of an antiviral activity in hIFN-λ3. However, when alanine substitutes this residue, the antiviral activity of hIFN-λ3 is reduced or abrogated (Gad and others 2009). Interestingly, residue F158 is highly conserved in mammalian and avian IFN-λ protein (Gad and others 2009). It has revealed that goIFN-λ perhaps shares similar biological and antiviral activities with hIFNL-λ.
In mammals and avian, type III IFNs and its functional receptor, especially the IFNLR1 chain, have a limited cellular distribution, such as gastrointestinal tract and respiratory tract mucosa-related immune organs (Sheppard and others 2003; Zhou and others 2007; Sommereyns and others 2008; Pulverer and others 2010; Dickensheets and others 2013). More recently, many systematic studies have found that IFN-λs has a prominent response in the stomach, intestine, lung, skin, and some epithelia-related tissues (Sommereyns and others 2008; Mordstein and others 2010; Pulverer and others 2010; Pott and others 2011). On the other hand, it has a very low expression level in the brain, heart, muscle, and central nervous system (Sommereyns and others 2008). Our previous research shows that type III IFN-goIFNLR1 of Chinese goose is mainly expressed in epithelium-rich tissues, such as small intestinal, lung, liver, and stomach (Zhou and others 2015). In this study, we found that goIFN-λ has a paralleled distribution with its functional receptor goIFNLR1. Both goIFN-λ and goIFNLR1 showed a relatively high expression in the pancreatic tissue. This was consistent with some previous studies (Lind and others 2013; Zhou and others 2015).
Similar to type I IFNs, IFN-λs can be induced in response to virus infection and TLR ligands (Ank and others 2006; Uzé and Monneron 2007). Previous research revealed that poly (I:C) is a synthetic dsRNA analogue and the agonist of TLR3 (Matsumoto and Seya 2008). ODN2006 as the B-type CpG-ODNs can effectively activate the B cells and is the agonist of TLR9 (Bauer and others 2001). Research indicated that no TLR9 exists in poultry species, but do have TLR21, which is functionally equivalent to the mammalian TLR9 (Keestra and others 2010). In this report, the mRNA transcriptional level of goIFN-λ showed a significant rise after the stimulation of PBMCs with ploy (I:C) and ODN2006. This finding showed that goIFN-λ can be induced by the agonist of TLR3 and TLR21.
Classical GPV causes a high morbidity and mortality rate among goslings and Muscovy ducklings < 4 weeks old with gross pathological changes, including catarrhal enteritis and fibrinous exudates within the enteric cavity. However, the host immune response against it is largely unknown. Our previous study reported that GPV could upregulate the mRNA level of IFN-α, IFN-γ, IL1β, and IL6 in goose PBMCs (Zhou and others 2016a). In this study, the mRNA transcriptional level of goIFN-λ showed a significant upregulation after the PBMCs were treated with GPV, which is consistent with the previous data.
Previous studies have shown that hIFN-λ plays an effective role in inhibiting low pathogenic avian influenza virus multiplication in lung epithelial cells, such as H5 N2, H5 N3, and H9 N2 (Sutejo and others 2015). Moreover, chIFN-λ can markedly inhibit the replication of highly pathogenic influenza A virus (Reuter and others 2014; Zhang and others 2015). In our research, after goslings were artificially infected with H9N2 AIV, the mRNA transcriptional level of goIFN-λ upregulated nearly 20-fold in the lungs and nearly 40-fold in the pancreas. It may be because H9N2 AIV is mainly located and replicated in the respiratory system, which belongs to the mucosal tissue, where IFNLR1 is primarily distributed. According to our previously published data, the H9N2 AIV antigen was most strongly distributed in the lungs (Zhou and others 2016b). It is suggested that goIFN-λ might play a pivotal role in anti-AIV defense of aquatic birds. GoIFN-λ showed a significant increase after the PBMCs were treated with GPV. It is suggested that goIFN-λ might play a role in anti-GPV defense of aquatic bird. Altogether, in this article, we have revealed that goIFN-λ is mainly expressed in epithelium-rich tissues. The immunological characteristics studies suggested that it might play a pivotal role in the mucosal immune protection and antiviral defense. All these finding provide a solid theoretical basis for further research on the function of goIFN-λ.
Footnotes
Acknowledgments
This work was funded by National Natural Science Foundation of China (31201891), The Ph.D. Programs Foundation of Ministry of Education of China (20125103120012), Sichuan Provincial Cultivation Program for Leaders of Disciplines in Science (2012JQ0040), Major Project of Education Department in Sichuan Province (12ZA107), Innovative Research Team Program in Education Department of Sichuan Province (No.12TD005, 2013TD0015), National Science and Technology Support Program (2015BAD12B05), National Special Fund for Agro-scientific Research in the Public Interest (201003012), and China Agricultural Research System (CARS-43-8).
Author Disclosure Statement
The authors declare no conflicts of interest.