
Abstract
Curcumin has shown protective potential on osteoarthritis. However, its effect on treatment of osteoarthritis remains elusive so far. This study aimed to determine whether curcumin could ameliorate osteoarthritis in vivo and the underline mechanisms. The mice subjected to destabilization of the medial meniscus (DMM) surgery were administered curcumin. Cartilage integrity was evaluated by immunohistological staining. Expression levels of inflammatory cytokines from mice arthrodial cartilage were detected. THP-1 cells were primed by lipopolysaccharide (LPS)/ATP to induce inflammation, followed by the addition of curcumin. The expression of proinflammatory cytokines was also detected. Moreover, the expression of pro-caspase-1, cleaved caspase-1, and NLRP3 inflammasome was examined. Administration of curcumin significantly reduced osteoarthritis disease progression in DMM model of osteoarthritis. Curcumin suppressed mRNA expression of proinflammatory mediators in arthrodial cartilage of mice subjected to surgery. In LPS- and ATP-induced THP-1 macrophage cells, curcumin significantly suppressed the expression of interleukin 1 beta (IL-1β) and tumor necrosis factor alpha (TNF-α) at both RNA and protein levels. Compared to vehicle-treated controls, curcumin also showed remarkably increased pro-caspase-1 and decreased cleaved caspase-1. This study provides the first evidence that curcumin exerts protection on osteoarthritis by inhibition to the release of inflammasome NLRP3, leading to the downregulation of inflammatory cytokines.
Introduction
O
Although the disease cause remains unclear, inflammation has significant relevance in the pathogenesis of osteoarthritis (Scanzello and Goldring 2012), and the conventional treatments for osteoarthritis include the use of acetaminophen or nonsteroidal anti-inflammatory drugs (Zhang and others 2010). Nevertheless, with long-term use in chronic disease treatment, these drugs sometimes cause serious gastrointestinal and cardiovascular adverse events (Ameye and Chee 2006). Given the limitation of current drugs for osteoarthritis, there is great need to deepen understanding of the inflammatory pathways in the progression of osteoarthritis, which could enable development of targeted disease-modifying osteoarthritic drugs.
Curcumin is the best-characterized chemopreventive agent extracted from the roots of Curcuma longa (Turmeric) (Henrotin and others 2010). It has long been used as an anti-inflammatory treatment in traditional Chinese and Ayurvedic medicine (Goel and others 2008) and some studies have suggested that curcumin exhibits antioxidant and anti-inflammatory properties (Jackson and others 2006; Shakibaei and others 2007).
Moreover, several extensive clinical trials on the effects of curcumin in osteoarthritis patients revealed that curcumin could reduce pain and improve the functionality of the patients (Chin 2016). Due to the effects of osteoarthritis on the articular cartilages such as the molecular destruction of the extracellular cartilage matrix, and the severe alterations of chondrocytes phenotypes (Roach and others 2007), the mechanistic studies on the antiosteoarthritic properties of curcumin were performed on primary chondrocytes isolated from human cartilage (Shakibaei and others 2007, 2011; Csaki and others 2009; Comblain and others 2015).
Curcumin reversed apoptosis of chondrocytes induced by interleukin 1 beta (IL-1β) by increasing the expression of antiapoptotic protein, such as Bcl-2, Bcl-xL, and TNF receptor-associated factor 1, and suppressing expression of caspase-3 (Csaki and others 2009). It has also been shown that curcumin inhibited the expression of matrix metalloproteinase (MMP)-3, MMP-9, cyclooxygenase 2 (COX-2), and prostaglandin E-2 in human primary chondrocytes induced by IL-1β or tumor necrosis factor alpha (TNF-α) (Shakibaei and others 2007, 2011).
Unconventional secretion of proinflammatory cytokine IL-1β has been reported to be associated with inflammasomes, which are large cytosolic multiprotein complexes that lead to the activation of caspase-1-mediated inflammatory responses (de Zoete and others 2014). NLRP3 is by far the most thoroughly studied inflammasome. NLRP3 is expressed at low levels in myeloid cells and the transcription can be induced by Toll-like receptor agonists, such as LPS, and by inflammatory cytokines, such as TNF-α (Bauernfeind and others 2009).
However, there are few reports about targeting inflammasomes for the treatment of osteoarthritis. Clavijo-Cornejo and others (2016) showed an increase in the protein expression of NLRP3 was related to the chronic inflammatory state present in patients with knee osteoarthritis. Jin and others (2011) demonstrated that the NLRP3 inflammasome mediated the pathological effect of hydroxyapatite crystals by activating caspase-1 and the secretion of IL-1β and IL-18, providing a mechanism for the critical role for the inflammasome in the pathogenesis of osteoarthritis.
The aim of this study was to investigate whether curcumin has effects on inflammasome activation in the amelioration of osteoarthritis using mouse surgical destabilization of the medial meniscus (DMM) model of osteoarthritis and THP-1 monocyte–macrophage cell line to decipher the potential of curcumin as targeted disease-modifying osteoarthritic drugs.
Materials and Methods
Animals
Male C57BL/6 mice (20–25 g) were purchased from Sino-British Sippr/BK Lab Animal Ltd. (Shanghai, China). The rats were housed in an isolated room with controlled temperature and a 12-h light/12-h dark schedule. Mouse food and water were available ad libitum. All experimental procedures were ethically approved by the Animal Use and Care Committee of the Fifth Hospital of Harbin and were conducted in accordance with the National Institute for Health “Guide for the Care and Use of Laboratory Animals.”
Cell culture and reagents
Curcumin, lipopolysaccharide (LPS), and ATP were purchased from Sigma-Aldrich (St. Louis, MO). Enzyme-linked immunosorbent assay (ELISA) kits for IL-1β and TNF-α were purchased from Thermo Fisher Scientific (Waltham, MA). The following monoclonal antibodies were obtained from Cell Signaling Technology (Danvers, MA): anti-caspase-1, anti-cleaved caspase-1, anti-ASC, and anti-beta actin antibodies. Anti-NLRP3 antibody was purchased from Abcam (Shanghai, China); Cell-counting kit-8 assay (CCK-8) was obtained from DOJINDO molecular technologies (Kumamoto, Japan). Human mononuclear macrophage cell line THP-1 was purchased from Shanghai Institute of Cell Biology (Shanghai, China) and maintained in RPMI 1640 medium from Hyclone (Logan, UT), supplemented with 100 U/mL of penicillin, 100 mg/mL of streptomycin, and 10% fetal calf serum under a humidified 5% (v/v) CO2 atmosphere at 37°C.
Surgical mouse osteoarthritis model and histological evaluation
The mouse surgical osteoarthritis model was modified based on the methods described by Glasson and others (2007) one decade ago (Glasson and others 2007). Specifically, the medial meniscotibial ligament (MMTL) is transected to generate DMM. Control joints included no surgery and sham surgery in which the ligament was visualized, but not transected. At 4–8 weeks postoperation, mice were euthanized and the knee joints were fixed in 4% paraformaldehyde for 24 h for immunohistological staining with Safranin-O and Fast green, Masson's Trichome, and hematoxylin and eosin. Stained slides were scored using a semiquantitative grading scale 6 as described by Glasson and others, 2007. A total of 40 male C57BL/6 mice were randomly divided into no surgery group, sham group, osteoarthritis (OA) group, OA plus DMSO group, and OA plus curcumin group (daily intraperitoneally injected curcumin at 50 μM or vehicle only after surgery).
Cytokine analysis by real-time quantitative polymerase chain reaction
Total RNA was extracted from mice arthrodial cartilage and reverse transcribed to cDNA for quantitative polymerase chain reaction (PCR), which was performed with the BioRad CFX96 Touch™ Real-Time PCR Detection System (BioRad, CA) using iQTM SYBR1 Green Supermix (BioRad). The program for amplification was 1 cycle of 95°C for 2 min followed by 40 cycles of 95°C for 10 s, 60°C for 30 s, and 95°C for 10 s. The primer sequences used in this study were as follows: IL-1b, 5′-CTTCAGGCAGGCAGTAT CACTC-3′ (forward) and 5′-TGCAGTTGTCTAATGG GAACGT-3′ (reverse); IFN-γ, 5′-GAT ATCTCGAGGAACTGGCAAAA-3′ (forward) and 5′-C TTCAAAGAGTCTGAGGTAGAAAG AGATAAT-3′ (reverse); IL-17A, 5′-TCGAGAAGAT GCTGGTGGGT-3′ (forward) and 5′-CTCT GTTTAGGCTGCCTGGC-3′ (reverse); IL-18, 5′-G CCTCAAACCTTCCAAATCA-3′ (forward) and 5′-TGGATCCATTTCCTCAAAGG-3′ (reverse); TNF-a, 5′-CGAGTGACAAGCCTGTAGCCC-3′ (forward) and 5′-GTCTTTGAGAT CCATGCCGTTG-3′ (reverse); VCAM1, 5′-GGAGACACTGTCATTATCTCCTG-3′ (forward) and 5′ -TCCTTTCATGTTGGCTTTTCT TGC-3′ (reverse); and beta-actin, 5′-GTATGCCTCGGTC GTACCA-30′ (forward) and 5′-CTTCTG CATCCTGTCAGCAA-3′ (reverse).
Cytokine analysis by enzyme-linked immunosorbent assay
THP-1 cells in each group were treated with lysis buffer to extract total protein. The homogenate was centrifuged at 12,000 g at 4°C for 15 min. The amount of total extracted protein was determined by the Bradford protein assay kit (Pierce, Rochford, IL). The amount of IL-1β and TNF-α in the homogenate was quantified by the ELISA kit from Thermo Fisher Scientific (Waltham, MA).
Cell proliferation and apoptosis assay
The effect of curcumin on the proliferation of THP-1was determined by the cell-counting kit-8 assay (CCK-8) according to the recommendations of the manufacturer. Briefly, 5 × 103 cells per well were cultured for 24 h in a 96-well plate and then treated with dimethyl sulfoxide (DMSO) alone or 10 μM curcumin for the indicated time. The absorbance of each well was read on a spectrophotometer (Thermo Fisher Scientific) at 450 nm. Apoptosis was detected with an Annexin V-FITC & PI Apoptosis kit, according to the recommendations of the manufacturer (Biouniquer).
Western blot analysis
To investigate the mechanisms of the inhibition of curcumin on the inflammation, THP-1 cells were treated as described in Effects of Curcumin on proliferation or apoptosis of THP-1 cells section. Equal amount (20 μg per lane) of total proteins were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto PVDF membranes. The membranes were blocked by 5% nonfat milk in TBST buffer for 1 h at room temperature and incubated with primary antibodies against caspase-1, cleaved caspase-1, and anti-beta actin antibodies overnight at 4°C. Membranes were washed thrice with TBST, and were incubated with secondary antibodies for 1 h. The density of each band was measured by Amersham Imager 600 (GE Healthcare, Little Chalfont, Buckinghamshire, United Kingdom). In addition, anti-ASC antibody (Abcam, Cambridge, MA) was used to immunoprecipitate the homogenized cell lysate, and antibodies against ASC, NLRP3 inflammasome, and pro-caspase-1 were used for Western blotting as described above.
Statistical analyses
Data are expressed as mean and standard deviation (SD). Statistical analyses were performed with GraphPad Prism 6.02 software and statistical significance was set at P-value <0.05. t-test was used to analyze the histologic scores in DMM model between the curcumin-treated group and vehicle control. The expression levels of mRNA in mice model and THP-1 cells were assessed by analysis of variance (ANOVA).
Results
Curcumin inhibits inflammation in surgical mouse osteoarthritis model
The DMM model had more significant osteoarthritis than the no surgery and sham controls. Specifically, at 4 weeks postoperatively, low levels of osteoarthritis were observed in the no surgery and sham surgery groups utilizing the maximum scores method (scores of 1.0 ± 0.1 and 1.0 ± 0.2 out of a maximal score of 6.0). For the blank and DMSO group, the scores of the DMM model at 4 weeks were 4.0 ± 1.5 and 4.2 ± 1.7, respectively (Fig. 1A). At 8 weeks, similar scores were observed for the no surgery and sham groups, indicating no infection during the surgical process, while the scores for the blank and DMSO groups were 3.9 ± 1.5 and 4.3 ± 1.8, respectively (Fig. 1B), representing a progression from mild-to-moderate osteoarthritis. From 4 to 8 weeks, curcumin showed significant inhibition in the osteoarthritis progression caused by DMM model, with scores of 2.0 ± 0.5 and 2.1 ± 0.5, respectively (Fig. 1).

Box and whisker plots showing maximum histologic scores at 4 weeks
Curcumin suppresses inflammatory cytokine expression in surgical mouse osteoarthritis model
To determine the influence of curcumin on the cytokine expression in mouse surgical DMM model of osteoarthritis, levels of IL-1β, IFN-γ, IL-17A, IL-18, TNF-α, and VCAM1 in cartilage tissue samples were measured by quantitative RT-PCR. As shown in Fig. 2A–F, compared with the no surgery and sham surgery groups, the mRNA expressions of all detected cytokines in the surgical osteoarthritis group were significantly upregulated. In contrast, the levels of all detected cytokines showed remarkable downregulation in the curcumin-administrated group.
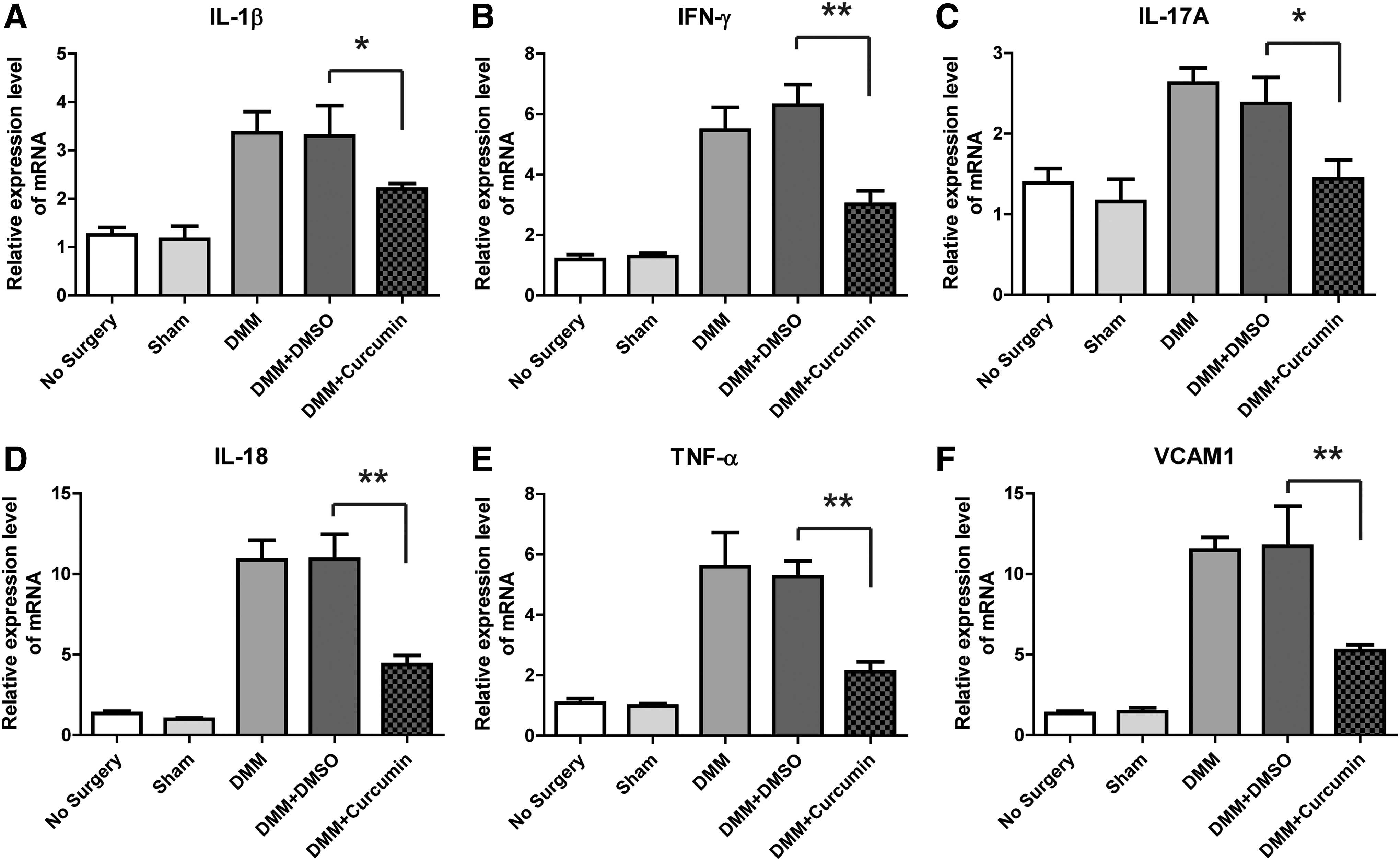
Curcumin suppressed the expression levels of the inflammation factors. Q-PCR showing the expression levels of IL-1β
Curcumin suppresses IL-1 β and TNF-α expression in THP-1 cells
To examine the effects of curcumin on the cytokine expression in THP-1 cells, levels of IL-1β and TNF-α in cultured cells were measured following induction of LPS and ATP. As shown in Fig. 3A, mRNA expressions of IL-1β and TNF-α were remarkably increased after LPS and ATP induction. Curcumin significantly inhibited the elevated expression of these cytokines triggered by LPS and ATP (P < 0.05). Moreover, we analyzed the cytokine expressions at protein level by ELISA. As shown in Fig. 3B, administration of curcumin to THP-1 cells significantly suppressed inflammatory cytokines at protein level (P < 0.01).
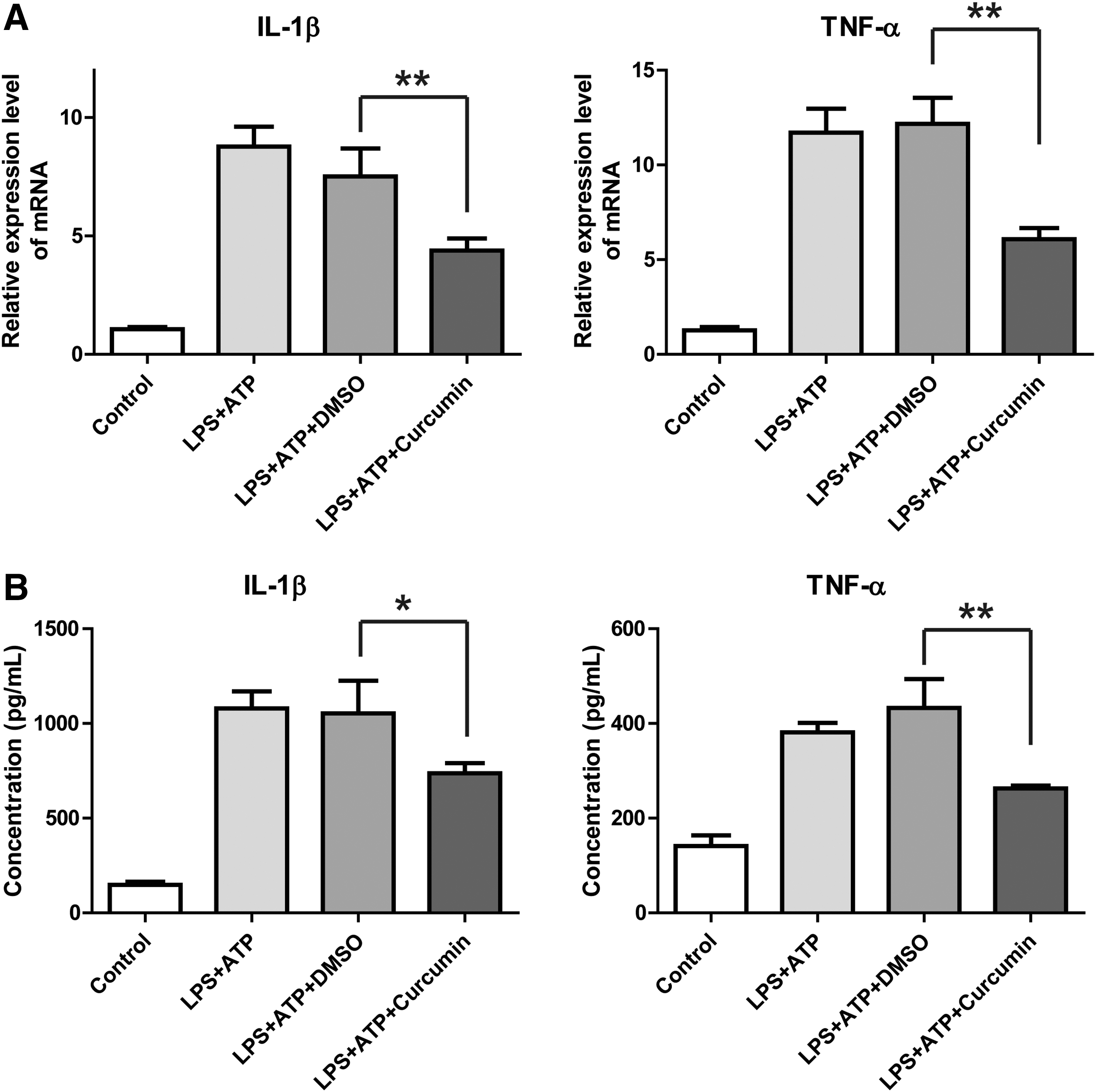
Curcumin suppressed IL-1β and TNF-α expression in THP-1 cells induced by LPS/ATP.
Effects of Curcumin on proliferation or apoptosis of THP-1 cells
The results of CCK8 experiment showed that the proliferation ability of THP-1 cells induced by LPS and ATP was not changed compared with the negative control group (0.2% DMSO) (Fig. 4A). After the cells were treated with LPS and ATP, cells were stained with PI and annexin V and then analyzed by FCM. The percentage of apoptotic cells did not exert a significant change compared to the DMSO group (Fig. 4B).
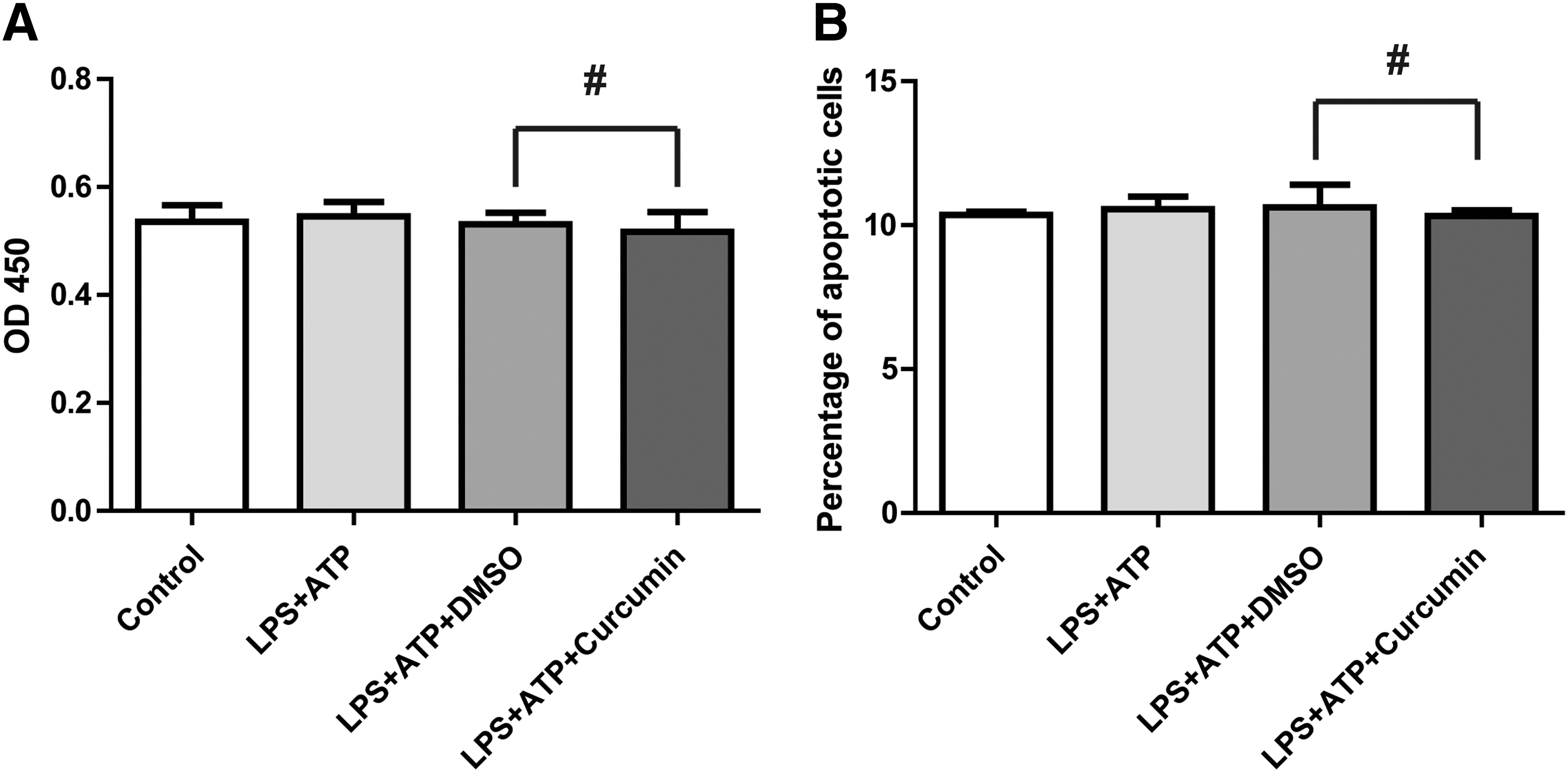
Curcumin did not affect proliferation or apoptosis on LPS-/ATP-induced THP-1 cells.
Curcumin suppresses inflammasome activation and caspase-1 cleavage
Since curcumin did not present inhibition on the THP-1 cell proliferation or cell apoptosis, there must be some other mechanisms for its suppression on inflammation. Given the significant inhibition of curcumin on IL-1β expression in the mouse osteoarthritis model and LPS-primed cells, we examined the ability of curcumin to inhibit the activation of caspase-1, which cleaves the precursor pro-IL-1β to mature active form of IL-1β.
As shown in Fig. 5A, the protein level of cleaved caspase-1 was significantly decreased with the administration of curcumin, nearly same as the protein level to the group without LPS/ATP induction. It has been reported that a caspase recruitment domain containing protein called ASC binds pro-caspase-1 to induce the autocleavage of pro-caspase-1 to produce the mature active form (Baroja-Mazo and others 2014), anti-ASC antibody was used to immunoprecipitate NLRP3 inflammsome and pro-caspase-1; the results showed that the protein level of pro-caspase-1 interacting with ASC and NLRP3 in curcumin group was significantly increased, in accordance with the decrease of activated caspase-1 (Fig. 5B). To be noted, as shown in Fig. 5B, the levels of ASC and NLRP3 were not significantly changed after curcumin treatment, indicating the specific targeting of curcumin on pro-caspase-1.
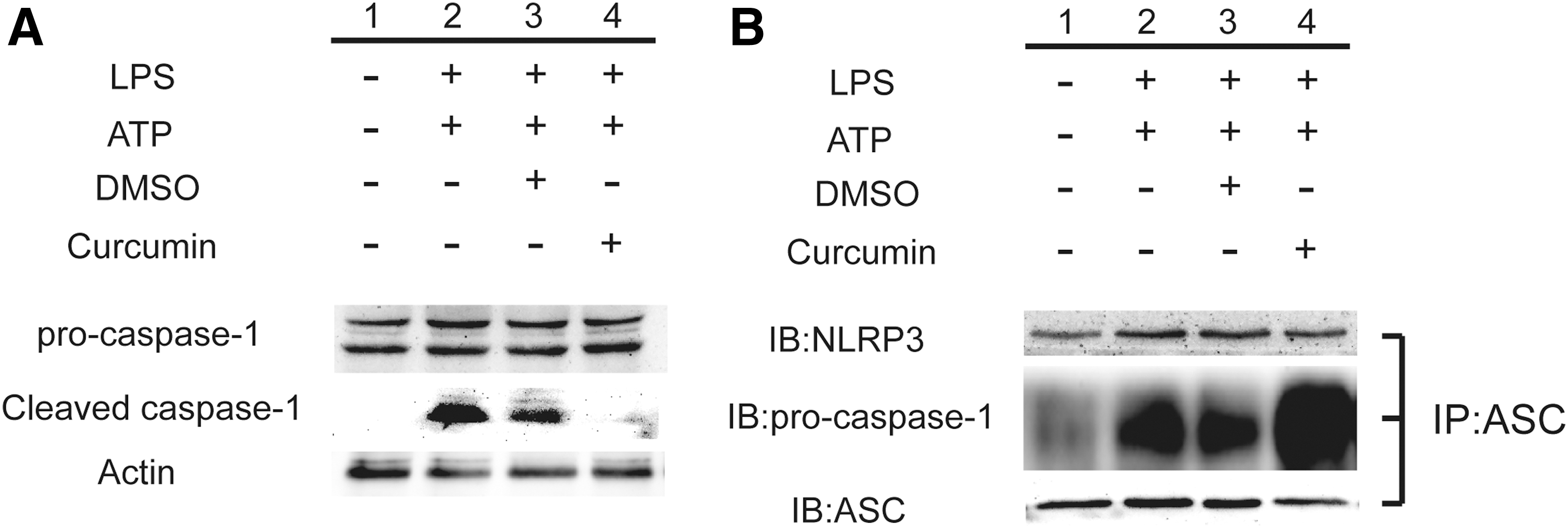
Curcumin inhibited the activation of inflammasome and prevented autocleavage of caspase-1.
Discussion
Osteoarthritis is a chronic inflammatory degenerative process that affects joints especially hips and knees in millions of people worldwide (Akuri and others 2017). As a promising natural agent, curcumin acts by either direct interaction with molecular targets or alteration of gene expression and signaling pathways (Shishodia 2013). In terms of osteoarthritis, curcumin is known for its potent anti-inflammatory and antioxidant properties, which could alleviate the osteoarthritis progression (Csaki and others 2009; Shakibaei and others 2011; Mobasheri and others 2012), and our studies confirmed this property.
Since the main effect of curcumin in osteoarthritis is related to the anti-inflammatory functions, the selection of an appropriate mouse model used for evaluation of curcumin efficacy is critical. Current available osteoarthritis animal models revealed over 20 different induction methods, including spontaneous, surgical, and chemical ways, in 10 different animals with varying strain and age (Little and Zaki 2012).
According to Zhang and others (2016), curcumin slows osteoarthritis progression and relieves osteoarthritis-associated pain symptoms in a mouse model of DMM. In addition, the transfection after surgical operation of DMM showed least influence in the osteoarthritis progression. This information supported the rational of utilization of DMM model in our study. In mouse surgical DMM model of osteoarthritis, curcumin exerted significant cartilage-protective and anti-inflammatory roles during the progression of osteoarthritis in the surgical group and suppressed the expression of several detected proinflammatory mediators, including IL-1β, IFN-γ, IL-17A, IL-18, TNF-α, and VCAM1. Curcumin has been reported to have anti-inflammatory properties on monocytes and macrophages (Abe and others 1999), as well as antiproliferative effects on T cells (Kim and others 2013).
Therefore, we used human monocyte–macrophage cell line THP-1 to further detect the effects of curcumin on the inflammation mediated by LPS/ATP. Our data showed that curcumin did not present significant effects on the proliferation and apoptosis of THP-1 cells at the same concentration, which exerted protection on osteoarthritis, indicating the low cytotoxicity of curcumin and veiled anti-inflammatory mechanisms.
In accordance with the results obtained from mouse osteoarthritis model, curcumin significantly inhibited IL-1β and TNF-α expression. IL-1β is a potent proinflammatory cytokine involved in a wide range of biological responses and its production is strictly regulated (Mezzasoma and others 2016). Primarily, NF-kB activation is required to produce pro-IL-1β (Ha and others 2016) and NLRP3 inflammasome/caspase-1 activation is required to cleave pro-IL-1β into the active secreted protein (Mezzasoma and others 2016). Due to the pleiotropism in a large variety of signals, IL-1β and NALP3 dysregulation is a common feature of a wide range of diseases and many of them are autoinflammatory diseases. One common characteristic of these autoinflammatory diseases is rapid disease alleviation on treatment with IL-1β blockade (Dinarello 2011).
However, to our best knowledge, there has been no report about the regulation of curcumin on IL-1β/NALP3/caspase-1 expression in the osteoarthritis progression. In this study, we compared NALP3/caspase-1 expression in LPS-/ATP-stimulated human THP1 monocytes with curcumin administration to DMSO group, finding that the protein level of cleaved caspase-1 was significantly decreased with the administration of curcumin. Since the adaptor protein ASC must become linearly ubiquitinated and phosphorylated for inflammasome assembly to occur (Guo and others 2015), we used anti-ASC antibody to immunoprecipitate the inflammasome complex and detected the protein levels of NALP3/caspase-1.
The results showed that curcumin-treated cells showed remarkably increased pro-caspase-1 and decreased cleaved caspase-1. For the first time, our data provided new evidence of the direct involvement of NALP3-/caspase-1-mediated IL-1β release in osteoarthritis. In particular, curcumin inhibited NALP3/caspase-1 activation leading to pro- and mature IL-1β downregulation, attributing to the cartilage-protective and anti-inflammatory roles of curcumin during the progression of osteoarthritis.
Curcumin has been shown to significantly decrease advanced glycation end product-stimulated TNF-α and MMP-13 mRNA and suppressed the NF-ĸB activation (Yang and others 2013); it would be interesting to know whether the down-regulation of IL-1β would be mediated by the NF-ĸB activation. Despite the consistency of the alleviated IL-1β levels in the animal osteoarthritis model and LPS-/ATP-induced cell line, it would be more straightforward to detect the activation of inflammasome complex in mouse surgical DMM model of osteoarthritis to conclude the relevance of IL-1β level with inflammasome NLRP3 release in further investigation.
Taken together, our data provide the first evidence that curcumin exerts protection on osteoarthritis by inhibiting the activation of inflammasome NLRP3, leading to the downregulation of inflammatory cytokines. These data could aid in the understanding of the inflammatory pathways in the progression of osteoarthritis and provide theoretical guidance for the clinical treatment of osteoarthritis, and ultimately enable development of targeted disease-modifying osteoarthritic drugs.
Conclusion
This study provides the first evidence that curcumin exerts protection on osteoarthritis by inhibiting the activation of inflammasome NLRP3, leading to the downregulation of inflammatory cytokines.
Footnotes
Author Disclosure Statement
The authors declare that they have no conflicts of interest.