
Abstract
This study investigated the anti-inflammatory effects and possible underlying mechanisms of Salvia miltiorrhiza polysaccharides (SMP) in lipopolysaccharide (LPS)-stimulated RAW264.7 cells. The cytotoxicity of SMP was detected by the MTT method. The morphological change of RAW264.7 was observed by Diff-Quik staining. Enzyme-linked immunosorbent assay was used to evaluate the production of cytokines in LPS-induced RAW264.7 cells. The nitric oxide (NO) kit assay detected the NO release from LPS-induced RAW264.7 cells. Real-time polymerase chain reaction was used to detect the transcriptions of tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), inducible NO synthase (iNOS), and cyclooxygenase (COX)-2 in LPS-induced RAW264.7 cells. The protein expression of nuclear NF-κB was measured by Western blot. The results showed that the safe medication range of SMP was less than 3 mg/mL. Compared with the LPS model group, SMP (2, 1, and 0.5 mg/mL) improved the degree of cell deformation and reduced the amount of pseudopodia, and statistically reduced the secretions of cytokines in cells induced by LPS (P < 0.01) at different time points. SMP significantly inhibited the mRNA transcriptions of TNF-α, IL-6, iNOS, and COX-2 and the protein expressions of NF-κB, p-p65, and p-IκBa. In conclusion, this study preliminarily proved the protective effect of SMP on LPS-induced RAW264.7 macrophage. Its mechanism might be related to inhibition of NF-κB signal pathway and the gene expressions and secretion of cytokines.
Introduction
T
Recent studies have shown that the Chinese herbal medicine and its active ingredients exert an inhibitory effect on inflammation (Huang and others 2012; Lai and others 2016), and has less side effects and adverse reactions to the body. Therefore, the underlying mechanism of inflammatory immune responses has drawn great attention in recent years.
Macrophages are important immunoregulatory cells that play a vital role in inflammatory response and tissue repair and maintain the body's steady state. At the same time, macrophage cells, a plastic and pluripotent cell population, show significant functional differences and secrete different inflammatory factors in different microenvironments. Macrophages are not only important effector cells in inflammation but also the antigen-presenting cells of microbial infection. RAW264.7 cell is a kind of macrophage of rodent mouse. The endotoxin is the main component of the cell wall of gram-negative bacteria. Lipopolysaccharide (LPS) can induce macrophages to differentiate into M1, release inflammatory cytokines, and resistance to pathogen invasion. However it also causes inflammation and tissue damage (Chen and others 2014). Thus, RAW264.7 stimulated by LPS is widely used as a target cell for studying inflammatory responses.
Salvia miltiorrhiza belongs to the plant family Oregano and is harvested in spring and autumn. The root part is commonly used to activate blood circulation to remove its stasis, cool the blood, and eliminate toxic materials. This herbal has been used in traditional Chinese medicine for more than 2,000 years to prevent and treat various human diseases such as hepatitis, coronary artery diseases, apoplexy, tumor, and immunological disorders. The modern pharmacological studies have found that one of the effective components, Tanshinone (Xie and others 2014; Maione and others 2015; Ma and others 2016), of S. miltiorrhiza has an anti-inflammatory and immunomodulation effect. However, whether another active ingredient of Salvia, S. miltiorrhiza polysaccharides (SMP), has an anti-inflammatory effect or not is unclear so far. Therefore, this study used LPS to induce inflammatory responses in macrophage RAW264.7 and established inflammation cell model to investigate the effects of SMP on inflammation. The aim of this report was to study the anti-inflammatory mechanisms of SMP in vitro and provide a theoretical and experimental evidence for the development of the new medication.
Materials and Methods
Extraction and purification of polysaccharide from S. miltiorrhiza
Dried roots of S. miltiorrhiza were purchased in a local market. S. miltiorrhiza roots were dried at 75°C and ground to fine powder. The ground powder samples were refluxed to remove lipids with acetoacetate:methanol solvent (1:1, v/v). After filtering, the residues were air-dried and then refluxed again with 75% ethanol at 75°C to remove oligosaccharides. The residues were extracted thrice in boiled water and filtered. The combined filtrates were concentrated by a rotavapor at 65°C and then precipitated using 95% ethanol. After filtering and centrifuging, the precipitate was collected, vacuum-dried, and represented a 3%–3.5% yield of raw material dry weight. The obtained SMP were stored in a refrigerator till further use.
LPS model and herbal treatment
The handling method of plate A was that the macrophage RAW264.7, purchased from the National Cell Resource Sharing Platform (Beijing, China), was seeded in 96-well plates with a density of about 1 × 106/mL. To determine the proper concentrations of SMP for the protection study, different solutions of SMP at 4, 3.5, 3, 2.5, 2, 1, and 0.5 mg/mL were added to the wells. Three concentrations were selected for use in plate B.
The handling method of plate B was that RAW264.7 was seeded in cell culture plates with a density of about 1 × 106/mL. Then they were divided into group A, the control group; group B, the LPS model group (LPS 1 μg/mL, Sigma Company); group C, LPS+SMP-H (2 mg/mL) group; group D, LPS+SMP-M (1 mg/mL) group, and group E, LPS+SMP-L (0.5 mg/mL) group. Various dosages of SMP were added to cell culture plates for protection (groups C, D, and E), while the control group and LPS model group received the same volume of serum-free medium. Four hours after SMP treatment, the LPS (1 μg/mL)-containing medium was added to the wells of groups B, C, D, and E, while the control group received the same volume of serum-free medium. Two plates were incubated for 24 h. Twenty microliters MTT at 5 g/L (Sigma Company) was added to each well. After the 4-h incubation at 37°C, the medium was removed and 150 μL DMSO added to dissolve crystals with gently shaking for 15–20 min. Then the supernatants were collected and the absorbance (a) value at 520 nm measured using a Microplate Reader (Bio Tek Company USA) to indicate cell viability.
Morphological changes of the cells
The RAW264.7 cells at logarithmic phase were seeded into culture plates. The cells were grouped and treated in the same manner as plate B (LPS model and herbal treatment section). After being cultured for 24 h, the cell supernatants were discarded and the Iml Diff-QuiK dye solution R1 was added to the wells for 5 min. Then the Diff-QuiK dye solution R1 was casted off and Diff-QuiK dye R2 was added. After the 1-min staining, the Diff-QuiK dye R2 was discarded. And then, the Diff-QuiK dye R3 was added for 5-min staining and then casted off. The plate was rinsed twice with PBS and the cell morphological changes were observed under an inverted microscope (CKX41; Olympus, Japan).
Expression of inflammatory mediators
The RAW264.7 cells at logarithmic phase were seeded into culture plates. The cells were grouped and treated in the same manner as plate B (LPS model and herbal treatment section). The supernatants of each group were collected to measure the absorbance at 12, 18, and 24 h post-LPS treatment. The concentrations of tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6,) IL-10, and Il-1β were detected using enzyme-linked immunosorbent assay (ELISA) kits according to the manufacturer's instructions (R&D System). The content of nitric oxide (NO) in cell supernatant was determined by Griss method (Promega Company).
The transcription levels of TNF-α, IL-6, cyclooxygenase-2, and inducible NO synthase
The RAW264.7 cells at logarithmic phase were seeded into culture plates. The cells were grouped and treated in the same manner as plate B (LPS model and herbal treatment section). At the end of the culture, total RNAs were extracted from each group by the Trizol method. According to the manufacturer's instructions, the mRNA was reversely transcribed as cDNA, which was used to detect the relative transcription levels of TNF-α, IL-6, and cyclooxygenase (COX)-2, and inducible NO synthase (iNOS) genes by real-time polymerase chain reaction (RT-PCR). PCR primers are shown in Table 1. RT-PCR conditions were 95°C for 3 min; 95°C for 15 s and 60°C for 20 s, cycled 40 times; and 55–95°C cycled 81 times. The amount of mRNA transcription was calculated using the 2−ΔΔCt method and β-actin was used as the internal reference gene.
COX, cyclooxygenase; IL-6, interleukin-6; iNOS, inducible nitric oxide synthase; PCR, polymerase chain reaction; TNF-α, tumor necrosis factor alpha.
Western blot for NF-κB expression
The RAW264.7 cells at logarithmic phase were seeded into 6-well culture plates. The cells were grouped and treated in the same manner as plate B (LPS model and herbal treatment section). After a 2-h incubation, total protein and nuclear protein were extracted according to the kit descriptions. Protein concentrations of each group were determined with BCA (ThermoUSA) method. Ten microliter protein sample (containing 40 μg protein) was added to the centrifuge tube mixed with 3 μL protein buffer, denatured in boiling water for 10 min. The mixture was concentrated at 5% gel and separated at 10% gel 100 V for SDS-PAGE electrophoresis 2.5 h. The gel was transferred by the semidry transfer (Bio-Rad) method at 12 V for 1 h to PVDF membrane and blocked with 5% BSA for 1 h. Then, it was incubated with anti-NF-κB antibody overnight (4°C) and washed with TBST 4 times at 10-min interval. After that, HRP-labeled secondary antibody was added and reacted at room temperature for 2 h. Then, the reaction was visualized in ECL after TBST washing for 10 min × 4 times. Finally, the expression of target protein was recorded and calculated.
Statistical analysis
Data were recorded using the Excel database and further analyzed using SPSS19.0 software (IBM Corporation, Armonk, NY). Differences in parameters were analyzed by analysis of variance (ANOVA). The significant levels were P < 0.05(*) or P < 0.01(**).
Results
Effect of SMP on cell viability
No decreased cell viability was observed when the concentration of SMP was at 3 or 2.5 mg/mL. SMP at 3.5 and 4 mg/mL significantly decreased cell viability, while the cell viability was significantly upregulated at 2, 1, and 0.5 mg/mL SMP treatments (Fig. 1A). When 2, 1, and 0.5 mg/mL SMP were co-cultured with 1 mg/mL LPS, no decreased cell survival rates were observed; in contrast, a significant increase was seen in 2 mg/mL SMP treatment (Fig. 1B).

The effects of SMP on viability of RAW264.7 cells.
Protective effects of SMP on LPS-stimulated RAW264.7
As shown in Fig. 2, compared with the normal cells in the control group, the cells of LPS-treated group were severely deformed, large number of cells showing false tail or pseudopodia (the arrows), which indicates successful LPS-induced RAW264.7 damage. Compared with the LPS group, SMP improved the degree of cell deformation with less pseudopodia.

Result of RAW264.7 Diff-Quik staining.
Effects of SMP on TNF-α secretion of RAW264.7
As shown in Fig. 3, compared with the control group, the release of TNF-α was significantly increased (P < 0.01) in the LPS group at 12, 18, and 24 h after coculture. However, compared with the LPS group, the SMP groups significantly reduced the content of TNF-α (P < 0.01). The content of TNF-α in LPS group was significantly higher than that in SMP groups at 18–24 h postculture.

The effect of SMP on TNF-α in RAW264.7 macrophages induced by LPS. Data represented mean ± SD; n = 10 per group. Group A, the control group; group B, the LPS model group (LPS 1 μg/mL); group C, LPS+SMP-H (2 mg/mL) group; group D, LPS+SMP-M (1 mg/mL) group; and group E, LPS+SMP-L (0.5 mg/mL) group. **P < 0.01, compared with A group. ## P < 0.01, compared with B group. TNF-α, tumor necrosis factor alpha.
Effects of SMP on IL-6 secretion of RAW264.7
Compared with the control group, the release of IL-6 in LPS group was significantly increased (P < 0.01) at 18 or 24 h post-LPS treatment. Compared with the LPS group, SMP significantly decreased the contents of IL-6 (P < 0.05 or P < 0.01). The expression level of IL-6 was lower at 12 h post-LPS treatment. There was no difference observed between each group at this time point (Fig. 4).
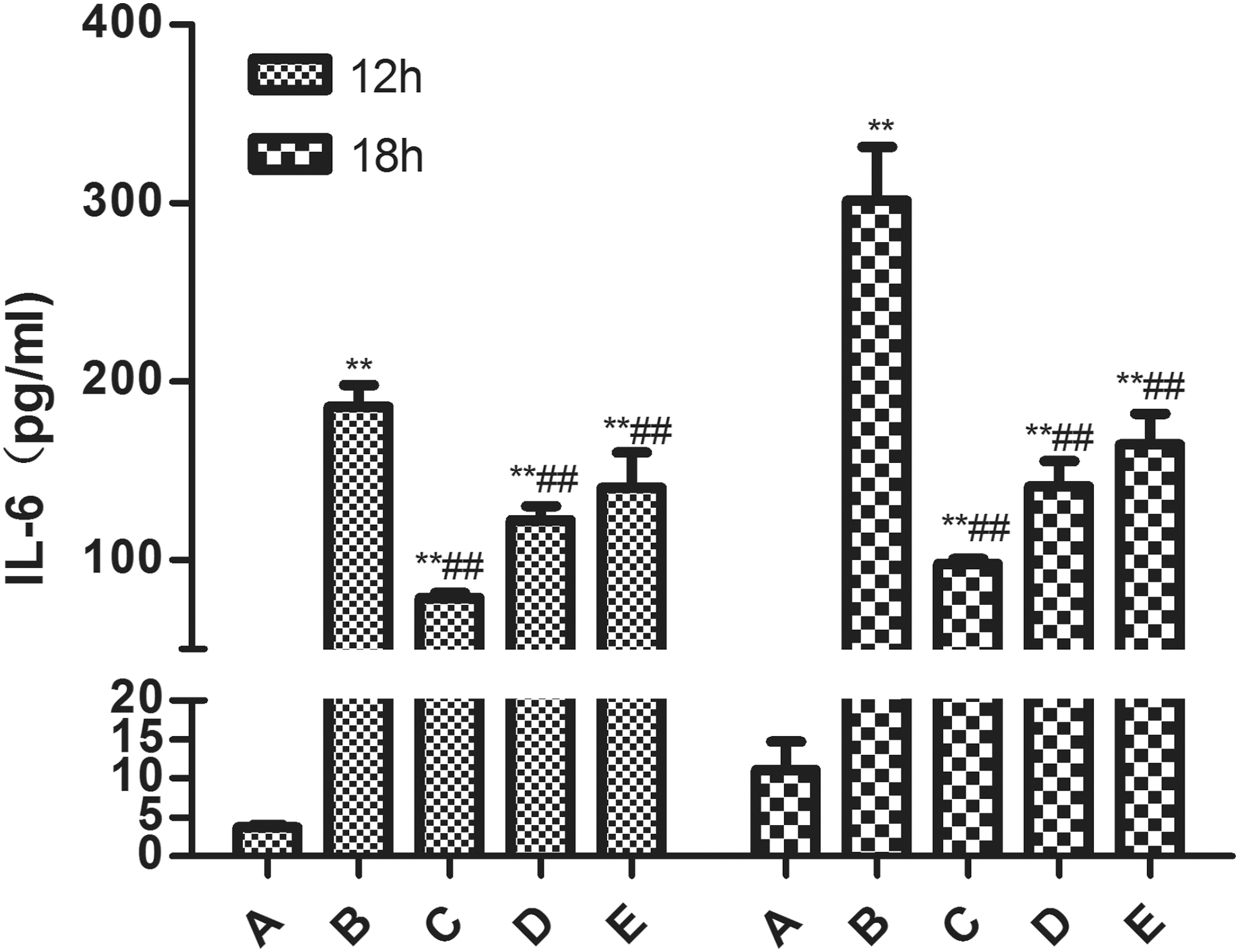
The effect of SMP on IL-6 in RAW264.7 macrophages induced by LPS. Data represented mean ± SD; n = 10 per group. Group A, the control group; group B, the LPS model group (LPS 1 μg/mL); group C, LPS+SMP-H (2 mg/mL) group; group D, LPS+SMP-M (1 mg/mL) group; and group E, LPS+SMP-L (0.5 mg/mL) group. **P < 0.01, compared with A group. ## P < 0.01, compared with B group. IL-6, interleukin-6.
Effects of SMP on IL-1β and IL-10 secretions of RAW264.7
Compared with the control group, the levels of IL-10 and IL-1β in the LPS group were significantly increased (P < 0.01) at 24 h posttreatment. Compared with the LPS group, SMP significantly reduced the releases of IL-10 and IL-1β. While the expression levels of each group were lower, no difference was found between groups at 12 or 18 h posttreatment (Fig. 5A, B).
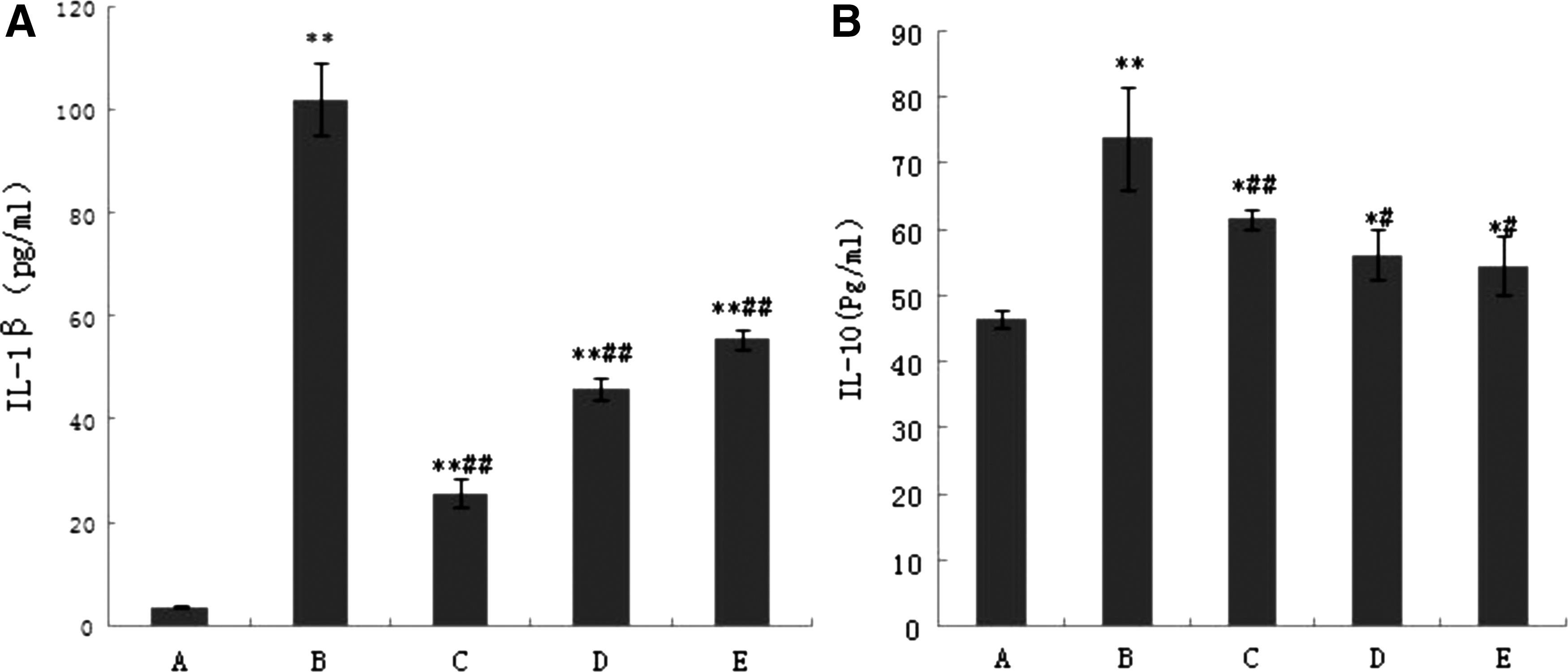
The effect of SMP on IL-1β
Effects of SMP on NO secretion of RAW264.7
Our results showed that compared with the control group, the release of NO was significantly increased (P < 0.01) in the LPS group at 12, 18, and 24 h posttreatment. Compared with the LPS group, SMP significantly reduced the increased level of NO (P < 0.01). The content of TNF-α in the LPS group was significantly higher than that in the SMP group at 12–24 h posttreatment (Fig. 6).

The effect of SMP on NO in RAW264.7 macrophages. Data represented mean ± SD; n = 10 per group. Group A, the control group; group B, the LPS model group (LPS 1 μg/mL); group C, LPS+SMP-H (2 mg/mL) group; group D, LPS+SMP-M (1 mg/mL) group; and group E, LPS+SMP-L (0.5 mg/mL) group. **P < 0.01, compared with A group. *P < 0.05, compared with A group. ## P < 0.01, compared with B group. # P < 0.05, compared with B group. NO, nitric oxide.
The gene relative transcription levels of TNF-α, IL-6, iNOS, and COX-2
Compared with the control group, the transcriptions of TNF-α, IL-6, iNOS, and COX-2 mRNA were significantly increased (P < 0.01) in the LPS-treated group. When compared with the LPS-treated group, SMP groups significantly inhibited TNF-α, IL-6, COX-2, and iNOS mRNA transcription (P < 0.01 or P < 0.05) (Fig. 7). Also, the reduction in TNF-α, iNOS, and COX-2 transcription showed a dose-dependent manner on the SMP.
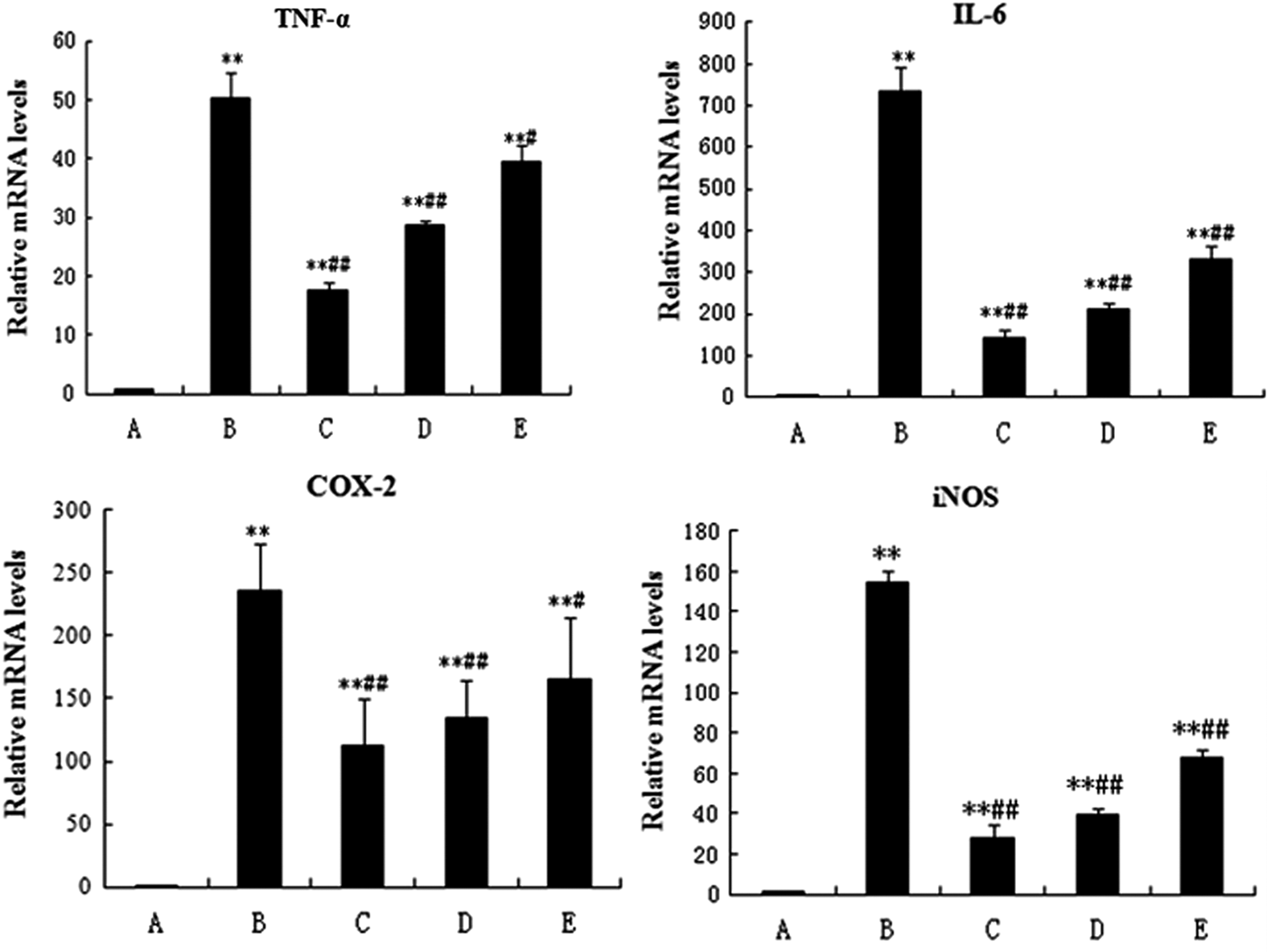
The effect of SMP on gene transcription of TNF-α, IL-6, iNOS, and COX-2 in RAW264.7 macrophages. Data represented mean ± SD; n = 10 per group. Group A, the control group; group B, the LPS model group (LPS 1 μg/mL); group C, LPS+SMP-H (2 mg/mL) group; group D, LPS+SMP-M (1 mg/mL) group; and group E, LPS+SMP-L (0.5 mg/mL) group. **P < 0.01, compared with A group. ## P < 0.01, compared with B group. # P < 0.05, compared with B group. iNOS, inducible nitric oxide synthase; COX, cyclooxygenase.
Protein expression of nuclear transcription factor NF-κB
We found that LPS significantly promoted the phosphorylation of p65 and IκBα (P < 0.01) (Fig. 8), whereas pretreatment with SMP for 4 h significantly inhibited the LPS-induced phosphorylation of p65 and IκBα with the high dose being the most effective (Fig. 8). The results indicate that SMP inhibits the inflammatory responses by regulating the expression of NF-κB.
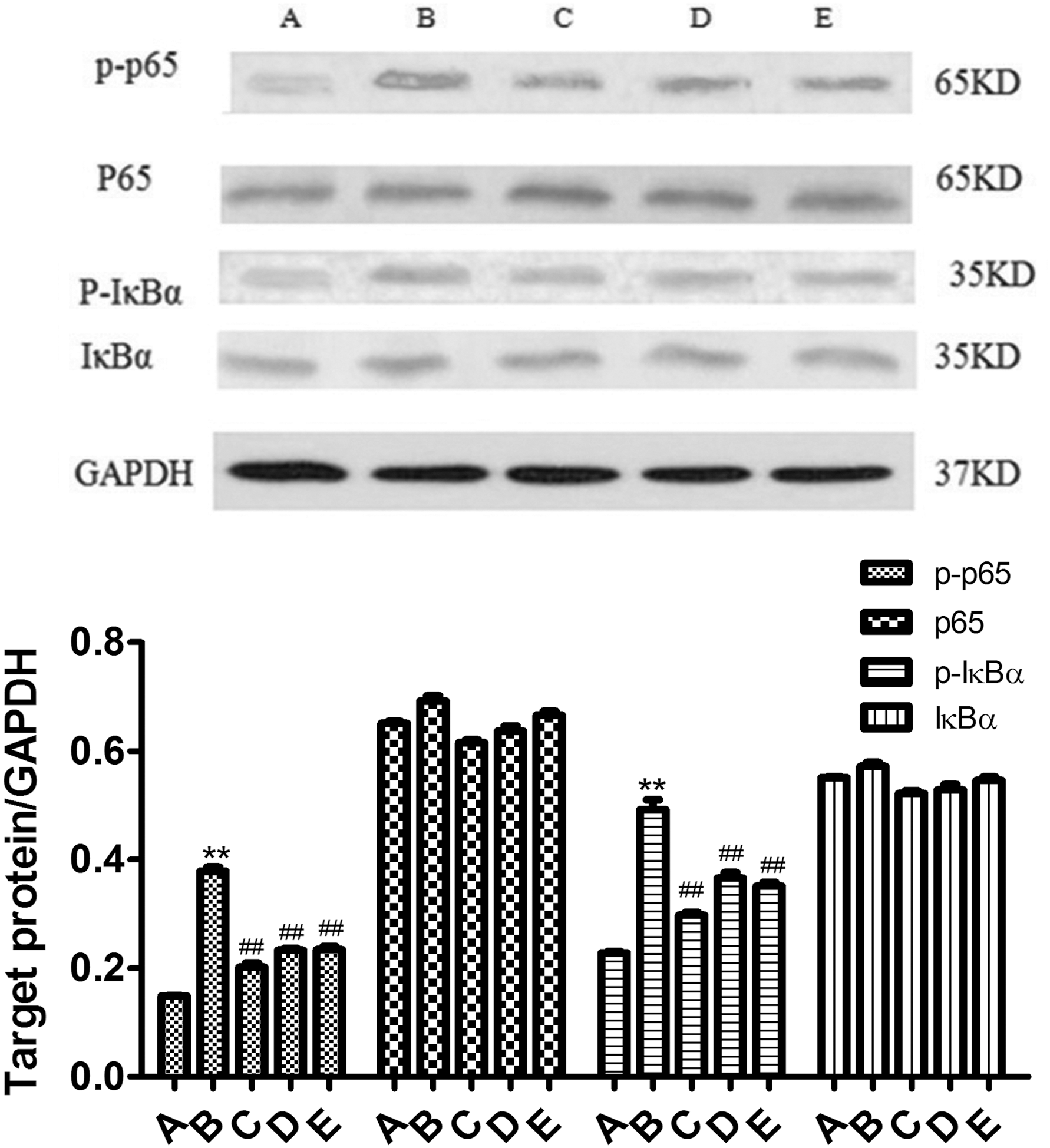
Effect of SMP on p-p65, p65, p-IκBα, and IκBα expressions in RAW264.7 cells. Data represented mean ± SD; n = 10 per group. Group A, the control group; group B, the LPS model group (LPS 1 μg/mL); group C, LPS+SMP-H (2 mg/mL) group; group D, LPS+SMP-M (1 mg/mL) group; and group E, LPS+SMP-L (0.5 mg/mL) group.**P < 0.01, compared with A group. ## P < 0.01, compared with B group.
Discussion
Inflammation is part of the complex biological response of body tissues to harmful stimuli, such as pathogens, damaged cells, or irritants (Ferrero-Miliani and others 2007), and is a protective response involving immune cells, blood vessels, and molecular mediators. The function of inflammation is to eliminate the initial cause of cell injury, clear out necrotic cells and tissues damaged from the original insult and the inflammatory process, and initiate tissue repair. Inflammatory response can surround and kill the invading pathogens through congestion and exudation reaction, and repair and heal the damaged tissue (Arroyo and Iruela-Arispe 2010; McCarthy 2015) through the regeneration of substantial and interstitial cells. Macrophages play an important role in the inflammatory process. Microorganisms stimulate macrophages to produce a variety of proinflammatory cytokines and chemokines, which in turn cause exudates and activate other types of immune cells (such as T cells). Some factors comediate the formation of immune inflammatory response (Biswas and others 2012; Mantovani and others 2013). Macrophages induced by LPS to produce inflammatory responses are often used to mimic microbial-induced inflammation in the body. Under the stimulation of LPS, macrophages secrete NO, TNF-α, IL-1, and IL-6 and other inflammatory cytokines involved in the occurrence and development process of inflammation (Takeuchi and others 1999; Ikonomidis and others 2012; Potnis and others 2013). RAW264.7 as a mouse macrophage cell line is widely used in inflammation tests (Ahn and others 2017; Kim and others 2017). Based on the above-mentioned references, this study used LPS to induce an inflammatory response model in macrophages to explore the anti-inflammatory effects and possible mechanisms of SMP in vitro. We found that 1 μg/mL of LPS successfully stimulated RAW264.7 to produce inflammatory responses. The MTT test revealed that the concentrations of SMP below 3 mg/mL showed no toxic effects on the cells and inhibited LPS-induced inflammatory response in macrophages.
Cytokines are a class of small molecule proteins with extensive biological activity and complex functions. TNF-α is mainly produced by monocyte/macrophages and is considered one of the most important cytokines that promote inflammatory responses, which can cause acute inflammatory diseases and tissue damage (Ahmad and others 2015; Yamanaka 2015). LPS stimulation and excessive production of NO will cause macrophage TNF-α secretion and induce further production of IL-1β and IL-6. IL-6 is also a major factor in the induction of inflammatory response, which can promote the differentiation of T lymphocytes and B lymphocytes; and other inflammatory cytokines synergistically cascade to promote and amplify the inflammatory response to tissue damage (Savale and others 2009; Yamanaka 2015). Thus, TNF-α and IL-6, as inflammatory markers, have an indication of the anti-inflammatory effects of drugs (Sen and others 2011). In this study, SMP significantly suppressed the releases of TNF-α and IL-6 in the cell inflammatory reaction and inhibited intracellular gene transcriptions of TNF-α and IL-6, indicating that SMP downregulates the increased proinflammatory factors by inhibiting the gene transcription. IL-1β is a critical proinflammatory cytokine that is secreted primarily by macrophages and monocytes. It plays a key role in the initiation of an inflammatory response and can induce the expression of other adhesion factors and cytokines or chemokines (Manjeet and Ghosh 1999; Budai and others 2013), causing adhesion of T lymphocytes to neutrophils and endothelial cells to participate in the process of vascular regeneration, to accelerate the formation of foam cells and atherosclerotic plaques (Dinarello 2009). Our study found that 24-h LPS treatment stimulated an increased IL-1β release significantly in RAW264.7 cells, whereas SMP significantly reduced the release of IL-1β in a dose-dependent manner. It is well known that the high expression of IL-10 has the effect of inhibiting inflammation and enhancing immunity (Wang and others 2009). Our experiment has demonstrated that a large number of LPS-stimulated RAW264.7 macrophages can lead to cellular inflammatory reactions, and thus the cells secrete large amounts of IL-10 to inhibit inflammation to further enhance. SMP and IL-10 synergistically exert anti-inflammatory effects, which may prevent a portion of IL-10 to release. Of course, the above speculation needs further testing.
NO participates in the regulation of the physiological processes of the body and the immune response, while excessive production of NO leads to tissue damage and inflammatory response. NO in vivo is produced by L-arginine (L-Arg) catalyzed by NOS. NOS can be divided into structural endothelial NOS (eNOS) and iNOS. eNOS produces only a small amount of NO and plays a normal physiological role. Pathogenic microbial infection and tissue damage can induce iNOS expression and promote a large amount of NO production. NO functions in both anti-inflammatory and proinflammatory responses (Welsh and others 2017). A small amount of NO can inhibit the inflammatory activity by inhibiting the adhesion of neutrophils and endothelial cells (Zhou and Zhu 2009). On the contrary, in the process of the inflammatory disease, excessive NO through the activation of NF-κB induced TNF-α, IL-6, and other proinflammatory cytokines to promote an inflammatory response. These cytokines can activate iNOS and promote the body to produce more NO. NO and cytokine secretion can be sustained, so that the inflammatory responses become more durable and intense (Medzhitov 2001). COX is a rate-limiting enzyme that converts the 4-dilute acid of peanut into prostaglandin. COX has 3 isozymes, in which COX-2 is an inducible expression. LPS and other proinflammatory factors induce COX-2 synthesis and further lead to a large amount of PGs synthesis, which participate in and amplify inflammatory responses (Wang and Dubois 2010; Chen and others 2012; Debprasad and others 2012). In the inflammatory process, if the release of inflammatory mediators NO and PGE2 and its catalytic enzyme activation were blocked, the inflammatory responses would be effectively inhibited. Our study showed that SMP acting on inflammatory cells can significantly decrease LPS-induced NO secretion and inhibited iNOS and COX-2mRNA transcription. The results of our study show that SMP inhibition of inflammation rate-limiting enzyme gene expression to decrease inflammatory mediator secretion is one of its anti-inflammatory mechanisms.
In addition, NF-κB as an important nuclear transcription factor plays an important role in the regulation of inflammatory responses, of which p65 is the most important subunit and participates in a variety of gene expression and regulation as indicator for activated cells (Li and others 2011). At resting state, NF-κB and IκB form a complex in the cytoplasm. IκB will be phosphorylated by activated kinetics kinase complex (IKB kinase, IKK), leading to NF-κB and IκB separation. The freed NF-κB p65 will be rapidly transferred to the cell nucleus and bound to the κB binding site to initiate inflammatory mediators and the proinflammatory cytokine transcription and expression (Gugasyan and others 2000; Bhatt and Ghosh 2014; Oh and others 2014; Bawadekar and others 2015; Zhao and others 2015). Our study found that SMP significantly inhibited the phosphorylation of IκB and p65. This indicated that SMP inhibited the dissociation of NF-κB and IκBa by reducing the phosphorylation of IκB and p65. Furthermore, SMP downregulated the activation and nuclear transfer of NF-κB, and reduced the expression of inflammation-related genes and inflammatory mediators.
Footnotes
Acknowledgment
This study was financially supported by the Ministry of Agriculture, China (No. 201403051-08).
Author Disclosure Statement
No competing financial interests exist.