
Abstract
Interferon-tau (IFN-τ), a novel type I interferon, is produced by trophoblast cells in ruminants. Previous studies have confirmed that IFN-τ could induce immunological tolerance in humans and other species. However, there are few reports on whether IFN-τ has a protective effect on embryo implantation failure caused by excessive inflammation at the maternal–fetal interface. In our study, a mouse model of lipopolysaccharide (LPS)-induced implantation failure was successfully established, and we investigated the protective effects of IFN-τ. First, we showed that IFN-τ increased the number of implanted embryos in LPS-treated pregnant mice. Subsequently, quantitative real-time polymerase chain reaction (qPCR) and ELISA results showed that several inflammatory cytokines [IL-1β and tumor necrosis factor-alpha (TNF-α)] whose expression was upregulated by LPS were reversed by IFN-τ treatment. Furthermore, we performed Western blotting and found that IFN-τ restrained the LPS-induced phosphorylation of IκBα and NF-κB p65. Moreover, qPCR and immunohistochemistry analyses showed that IFN-τ decreased the LPS-induced expression of mouse major histocompatibility complex (MHC) class I genes (H-2K and H-2D) in LPS-treated pregnant mice. Taken together, these results suggest that IFN-τ has a protective effect in LPS-induced implantation failure by downregulating MHC class I genes expression and inhibiting the production of inflammatory cytokines.
Introduction
E
It has been generally accepted that the maternal immune response to embryo implantation shares many similarities with inflammatory responses, whereas excessive fetal–maternal inflammatory response may lead to implantation failure, spontaneous abortion, and preterm birth (Romero and others 2004b). Numerous clinical studies have shown that genital tract microbial infections are associated with an increased incidence of infertility and implantation failure (Cram and others 2002). Lipopolysaccharide (LPS), a bacterial endotoxin with various biological effects, is a major component of the outer membrane of gram-negative bacteria. Toll-like receptor 4 (TLR4) is involved in host immune responses against microbial pathogens and can recognize exogenous molecules such as LPS (Wu and others 2016c; Jiang and others 2017b). TLR4 activation by LPS leads to activation of the NF-κB pathway and eventually promotes the production of inflammatory cytokines [such as IL-1β and tumor necrosis factor-alpha (TNF-α)] in macrophages (Hernandez and others 2016; Jiang and others 2018). Previous reports have indicated that bacterial LPS suppresses blastocyst implantation in mice by increasing the production of inflammatory cytokines at the maternal–fetal interface (Deb and others 2005a, 2005b).
Interferon-tau (IFN-τ) is a novel type I interferon produced by trophoblast cells in ruminants (Pontzer and others 1997; Martal and others 1998; Zhao and others 2016a), and plays a critical role for successful implantation and establishment of pregnancy (Bazer and others 1997; Todd and others 1998). However, it has been reported that IFN-τ has a broad cross-species reactivity in humans and other animals such as mice (Alexenko and others 1999). Intriguingly, treatment of spontaneous abortion mice with recombinant IFN-τ can also reduce embryo mortality (Chaouat and others 1995). Besides, increasing evidence has demonstrated that administration with IFN-τ leads to an anti-inflammatory functional phenotype and induces immunological tolerance in inflammation-related diseases (Soos and others 1995; Bazer and others 2012). However, there are few reports on whether IFN-τ can prevent embryo implantation failure caused by excessive inflammation at the maternal–fetal interface. In this study, we investigated the effect of IFN-τ on embryo implantation using a mouse model of LPS-induced implantation failure and elucidated the possible mechanisms.
Materials and Methods
Reagents
Recombinant ovine IFN-τ (rOvIFN-τ) (>97%, high-performance liquid chromatography purity) was purchased from Creative Bioarray (NY). LPS (Escherichia coli 055:B5) was purchased from Sigma (St. Louis, MO). NF-κB antibodies were purchased from Cell Signaling Technology (Danvers, MA). Anti-mouse H-2K/H-2D antibody was purchased from BioLegend (San Diego, CA). All other chemical reagents were of reagent grade.
Animals and treatment
Kunming mice (8 weeks old, weighing 25 ± 2 g) were obtained from Wuhan Institute of Biological Products Co., Ltd. (Wuhan, China). The mice were housed in a room maintained at 23°C with 60% humidity and had ad libitum access to food and water. All the animal experiments were carried out in strict accordance with the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health and approved by the Ethical Committee on Animal Research at Huazhong Agricultural University. The mice were caged overnight at a female:male ratio of 1:1 for mating. Successful mating was confirmed by the presence of a vaginal plug the next morning; if positive, that day was designated day 0.5 of pregnancy. The method for establishing the LPS-induced implantation failure model was described previously (Zhao 2011). In brief, gravid mice were intraperitoneally injected with 100 μL of LPS (4 μg) on day 3 of gestation. Gravid mice were randomly assigned into 4 groups (n = 10): (1) Control group: These mice received an injection of 100 μL of phosphate-buffered saline (PBS) on day 3 of gestation. (2) LPS group: Mouse model of implantation failure without drug treatment. (3 and 4) IFN-τ treatment groups: 6 h after LPS injection, these mice received 3 intraperitoneal injections of 4 or 8 ng/g IFN-τ at 6-h intervals. The IFN-τ doses used in the study were established in our previous studies (Wu and others 2016a).
On day 4.5 of gestation, half of the pregnant mice in each group were euthanized with sodium pentobarbital. The samples of uterus tissue were collected as previously described (Zhang and others 2015). One side of the uterine horns was carefully cleaned of fat tissue, and removed the fetuses by washing with ice-cold PBS. Subsequently, the uterine tissue was obtained and stored at −80°C for quantitative real-time polymerase chain reaction (qPCR) and Western blot analyses. The other side was fixed in 10% formalin for immunohistochemical analysis. The remaining mice were sacrificed on day 7 of gestation, and the number of implanted embryos was calculated.
Immunohistochemical analysis
Sections (4 μm thick) from the uteri were deparaffinized in xylene, rehydrated in a graded alcohol series, and then subjected to antigen retrieval in a microwave oven. To eliminate endogenous peroxidase, the slides were incubated in 0.3% hydrogen peroxide for 5 min. Subsequently, the samples were incubated in 10% normal horse serum for 30 min, followed by incubation with anti-mouse H-2K/H-2D antibody (1:200 dilution) at 4°C overnight. Samples were then incubated with horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG antibody at 37°C for 1 h. After staining with diaminobenzidine, images were captured using a light microscope.
Cell culture and treatment
RAW 264.7 murine macrophages were provided by the American Type Culture Collection (ATCC TIB-71™) and cultured in DMEM supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 50 U/mL penicillin, and 50 mg/mL streptomycin. The cells were pretreated with IFN-τ (20 or 40 ng/mL) for 1 h and then stimulated with LPS (0.5 μg/mL) for 24 h. Untreated cells served as controls.
RNA extraction and qPCR analysis
Total RNA was isolated from the uterus and cells with TRIzol reagent (Life Technologies) according to the manufacturer's instructions. Then, cDNA was synthesized by reverse transcription. PCR was performed using the SYBR Green Plus Reagent Kit (Roche, Basel, Switzerland) and an ABI Prism 7500 Sequence Detection System (Applied Biosystems). The primers used for target gene amplification are listed in Table 1. Relative quantification of target gene expression was normalized to GAPDH using the 2−ΔΔCt comparative method.
TNF-α, tumor necrosis factor alpha.
ELISA analysis
The tissue samples were homogenized in PBS and then centrifuged to collect the supernatants. The cell supernatants were also harvested. The levels of TNF-α, IL-1β, and IL-10 in these supernatants were measured using ELISA kits according to the manufacturer's directions.
Western blot analysis
Total protein was extracted from uterine tissue using radioimmunoprecipitation assay (RIPA) lysis buffer (Biosharp, China). Proteins were separated by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred onto a PVDF membrane. Subsequently, the membrane was incubated with specific antibodies against phosphorylated NF-κB p65 and β-actin (1:1,000 dilution).
Immunofluorescent detection of NF-κB p65 localization
Macrophages were grown on coverslips overnight and then treated with 0.5 μg/mL LPS alone or with IFN-τ (20 and 40 ng/mL). The cells were fixed in 4% paraformaldehyde for 15 min and then permeabilized with 0.2% Triton X-100 in PBS for 10 min. Nonspecific binding sites were blocked with 5% bovine serum albumin. After incubation with rabbit anti-NF-κB p65 antibody overnight at 4°C, the FITC-conjugated goat anti-rabbit IgG was applied for 30 min at room temperature. Then, the cells were stained with DAPI, and the NF-κB p65 subunit was examined under an inverted fluorescence microscope (Leica, UK).
Statistical analysis
All data are presented as the mean ± SEM. Differences between groups were analyzed by 1-way analysis of variance (ANOVA) using SPSS 15.0. A P-value of <0.05 was considered statistically significant.
Results
Effect of IFN-τ treatment on embryo implantation
The number of implanted embryos in different groups was calculated on day 7 of gestation. As shown in Fig. 1B, LPS significantly reduced the number of implanted embryos compared with the control. However, IFN-τ treatment prevented this reduction in the number of embryos.
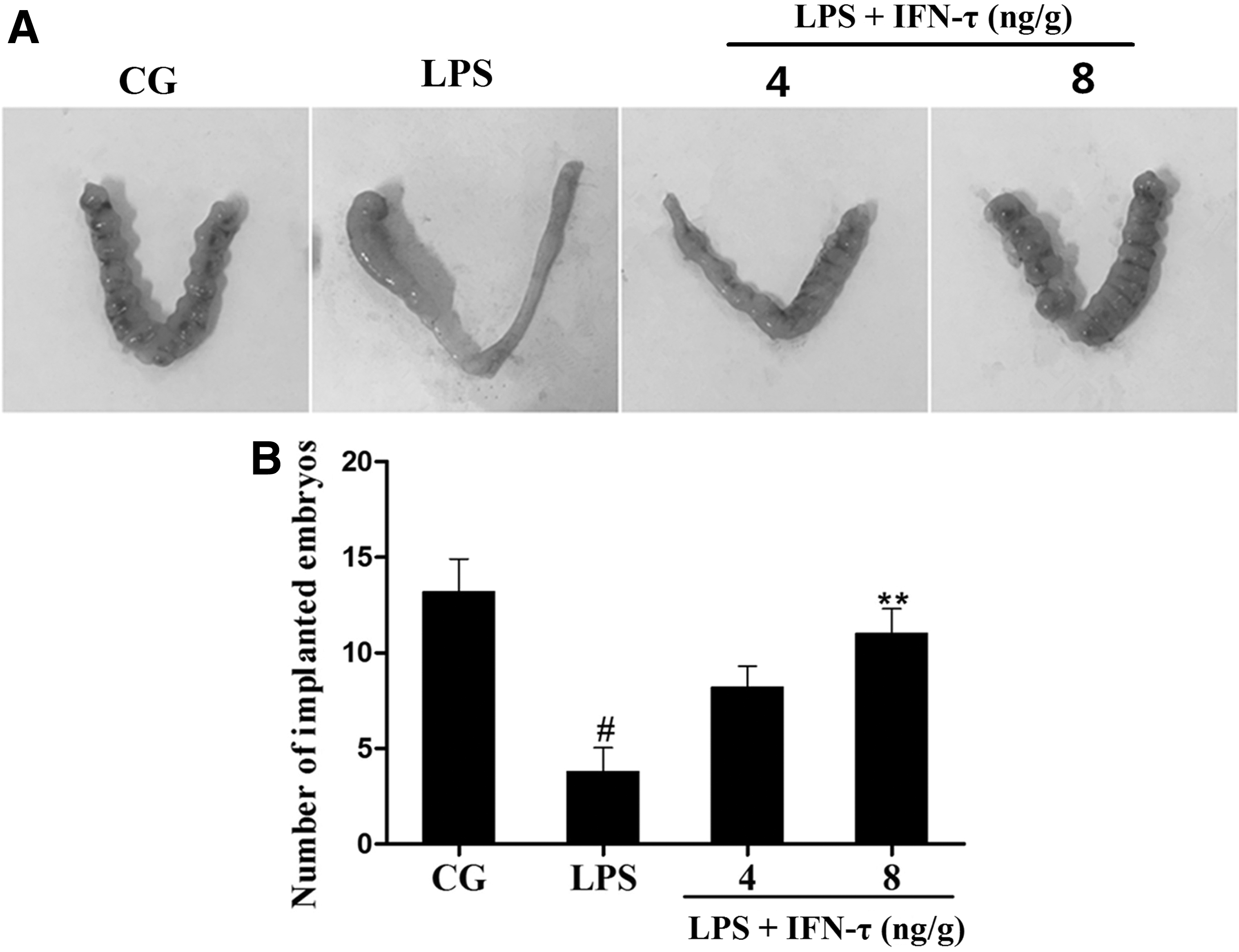
Effect of IFN-τ on mouse embryo implantation.
Effect of IFN-τ treatment on IL-1β, TNF-α, and IL-10 expression in the uterus
The expression of IL-1β, TNF-α and IL-10 in uterine tissue was analyzed by qPCR and ELISA, which showed that IL-1β and TNF-α expression was significantly increased in the LPS group, whereas IL-10 expression was slightly decreased. In contrast, IFN-τ treatment significantly decreased IL-1β and TNF-α levels and promoted IL-10 secretion (Fig. 2).
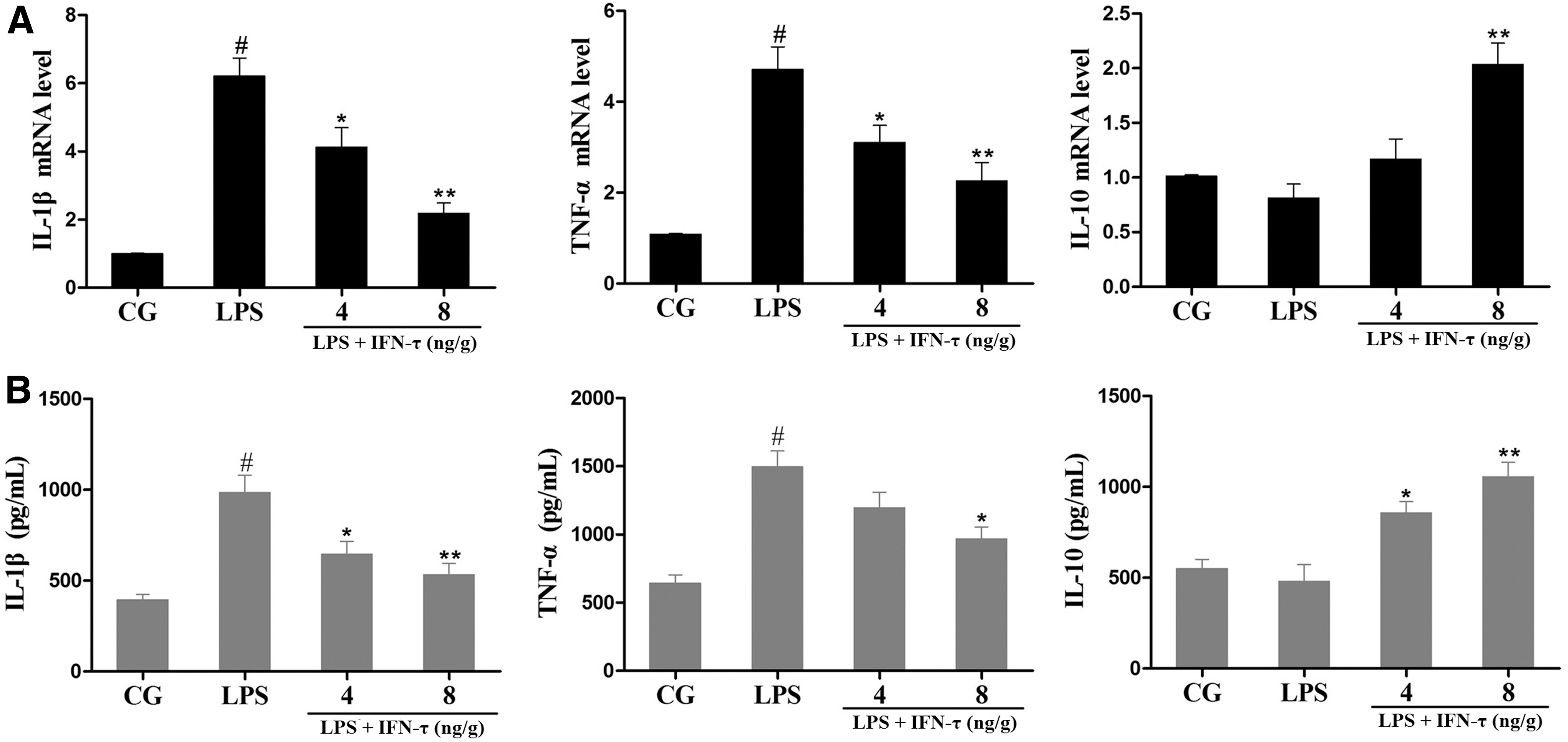
Effect of IFN-τ on cytokine production in uterine tissue.
Effect of IFN-τ treatment on NF-κB p65 levels in uterine tissue
The NF-κB pathway plays a critical role in the production of inflammatory cytokines (Jiang and others 2017c; Zhao and others 2017). Thus, the protein levels of NF-κB p65 and IκBα in the uterus were detected by Western blot. As expected, LPS significantly induced the phosphorylation of p65 and IκBα. However, the levels of phosphorylated p65 and IκBα were dramatically diminished by IFN-τ treatment in a dose-dependent manner (Fig. 3).

Effect of IFN-τ on the NF-κB pathway.
Effect of IFN-τ treatment on IL-1β, TNF-α, and IL-10 expression in macrophages
IL-1β, TNF-α, and IL-10 expression in macrophages was also measured by qPCR and ELISA. As shown in Fig. 4, the expression of IL-1β and TNF-α was significantly increased in the LPS group compared with the control group. However, IL-10 expression was not influenced significantly by LPS. IFN-τ treatment suppressed the production of IL-1β and TNF-α in a dose-dependent manner and simultaneously enhanced the level of IL-10.
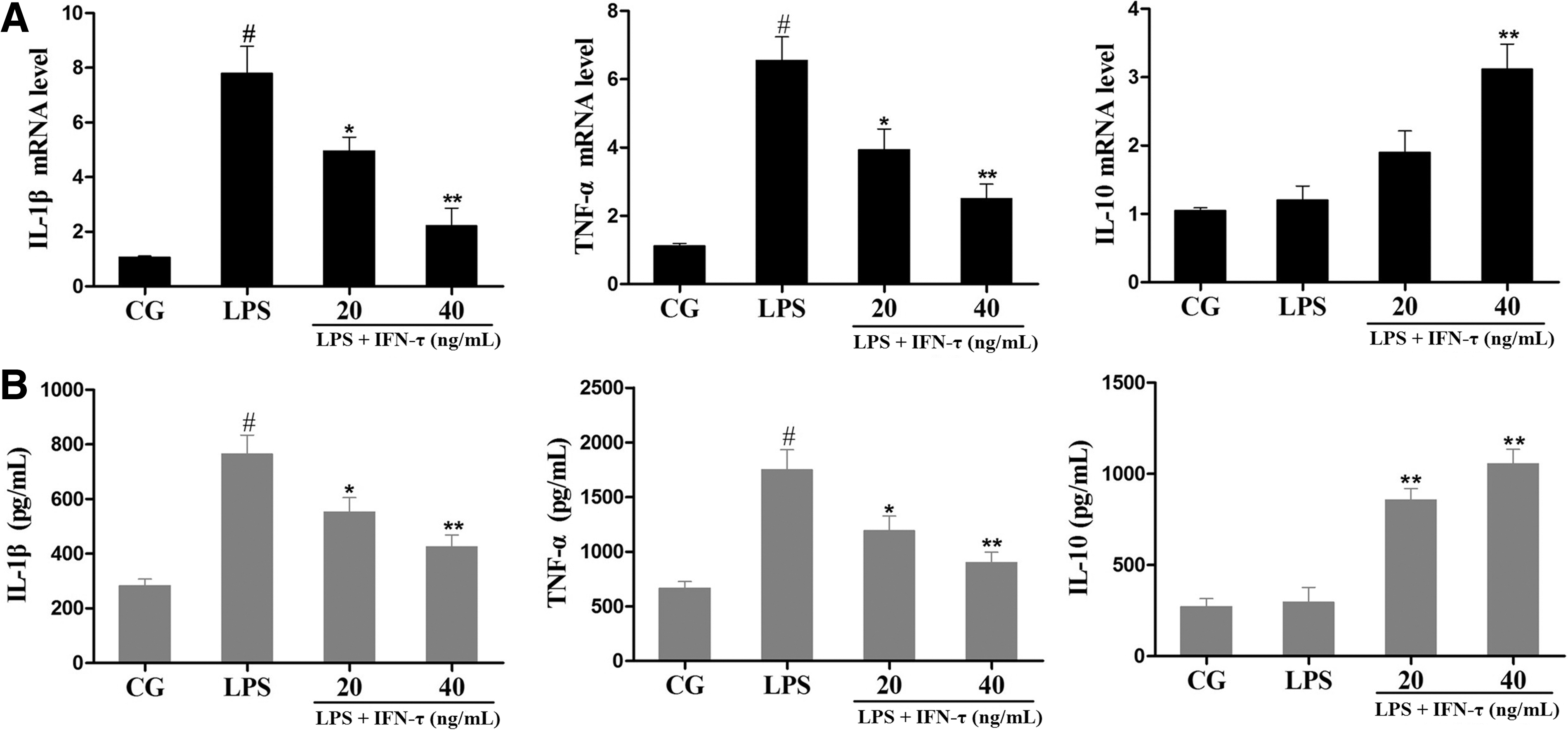
Effect of IFN-τ on cellular cytokines levels.
Effect of IFN-τ treatment on the nuclear translocation of NF-κB p65 in macrophages
Our results showed that LPS exposure induced the translocation of NF-κB p65 from the cytosol to the nucleus. However, IFN-τ treatment significantly suppressed NF-κB p65 nuclear translocation, consistent with the above results (Fig. 5).

Effect of IFN-τ on NF-κB p65 nuclear translocation. RAW 264.7 cells were pretreated with IFN-τ (20 or 40 ng/mL) for 1 h before stimulation with LPS, and NF-κB p65 nuclear translocation was observed with an inverted fluorescence microscope. Color images available online at
Effect of IFN-τ treatment on mouse major histocompatibility complex class I genes expression in the uterus
The expression of mouse major histocompatibility complex (MHC) class I genes (H-2K and H-2D) was measured by qPCR. As shown in Fig. 6, H-2K and H-2D expression was dramatically increased in the LPS group but decreased in the IFN-τ treatment group. Furthermore, immunohistochemistry revealed more intense expression of H-2K/H-2D in the LPS group compared with the control group, and this expression was reduced by IFN-τ treatment. In addition, H-2K/H-2D mainly localized in the uterine glandular epithelium and stroma (Fig. 7).
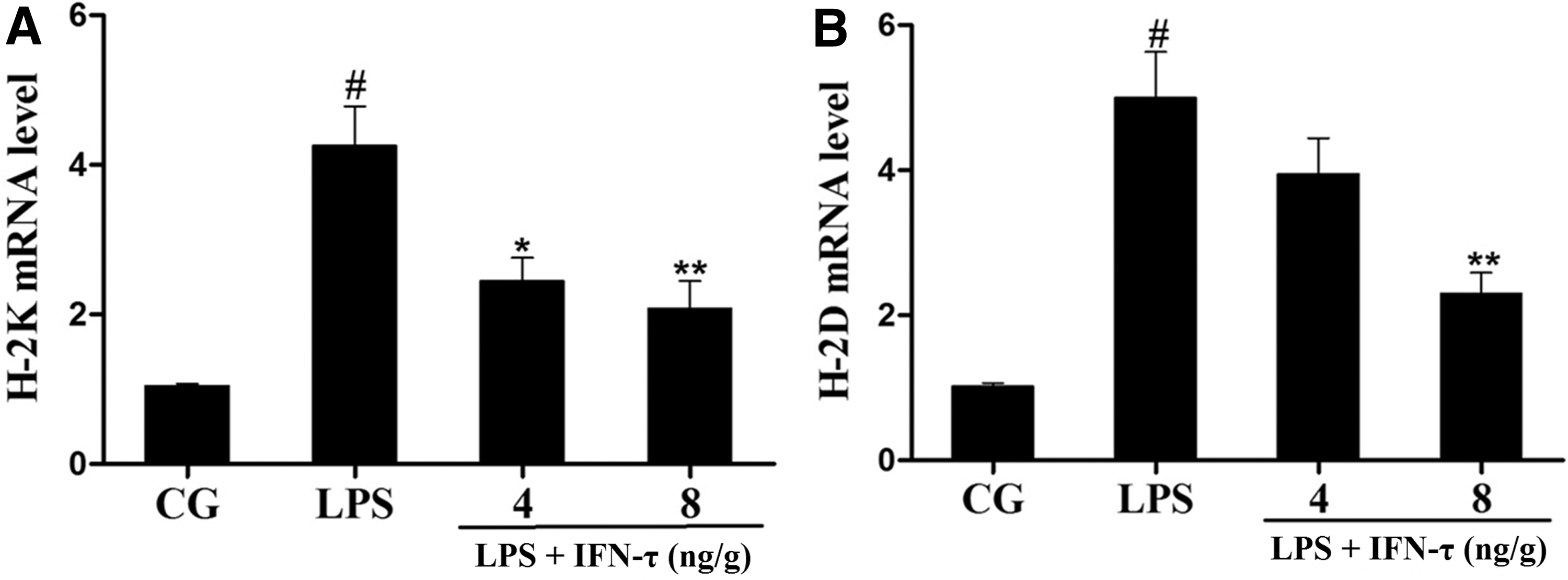
Effect of IFN-τ on MHC class I genes expression in uterine tissue. The expression of MHC class I genes (H-2K and H-2D) was measured by qPCR. GAPDH was used as a control.

Immunohistochemical analysis of H-2K/H-2D in the uterus. The arrows represent the positive staining for H-2K/H-2D.
Discussion
A successful or healthy pregnancy requires complex immune regulation in the uterine endometrium, eventually resulting in tolerance of the implanted embryos (Norwitz and others 2001). It has been well established that early pregnancy failure is usually caused by uncontrolled inflammatory responses at the maternal–fetal interface (Clark and others 1998). In such cases, a series of unfavorable factors may propel the uterine immune system toward anti-fetal immune responses. Indeed, microbial infection and other inflammatory stimuli are closely associated with early pregnancy failure in humans (Romero and others 2004a). E. coli is a common pathogenic bacterium that accounts for ∼80% of genital tract infections. Moreover, E. coli has been implicated in abnormal embryo development (Rupasri and others 1995). Thus, the development of effective therapeutics against infection-related pregnancy failure is urgently needed.
IFN-τ shares sequence similarity and functional homology with conventional members of the type I interferons family, such as IFN-α and IFN-β and particularly INF-ω (Jamwal and others 2009; Chon and Bixler 2010). Compared with IFN-α and IFN-β, IFN-τ exhibits very low cytotoxicity even at high dosages, which provides strong evidences for a clinic application of IFN-τ (Pontzer and others 1991). Although IFN-τ is generally recognized as the pregnancy recognition signal in ruminants, it has been reported to induce immunological tolerance in humans and other species. For instance, administration with recombinant IFN-τ decreases fetal resorption in spontaneous abortion mouse models by upregulating IL-10 levels, which prevents inflammatory immunity during gestation (Chaouat and others 1995). Furthermore, recent research has also revealed that IFN-τ alleviates obesity-induced adipose tissue inflammation and spontaneous autoimmune diabetes in mice, and suppresses LPS-induced pneumonia in a mouse model of acute lung injury (Soos and others 2002; Ying and others 2014; Zhao and others 2016b). Our previous studies also demonstrated that IFN-τ alleviates Staphylococcus aureus-induced endometritis (Jiang and others 2017a). However, we are not uncertain whether IFN-τ can block embryo implantation failure caused by excessive inflammation at the maternal–fetal interface. Previous evidence suggested that LPS may change the normal uterine receptivity necessary for embryo implantation and thus induce implantation failure in mice (Deb and others 2005a). Therefore, in the present study, we used LPS to induce embryo implantation failure in mice and investigated the protective effect of IFN-τ on embryo implantation. Our results found that the number of implanted embryos was decreased dramatically after LPS stimulation, but the reduction was prevented by IFN-τ treatment, indicating that IFN-τ has a protective effect on implantation failure.
The immune system balance, especially that of Th1/Th2 cytokines, at the maternal–fetal interface is necessary for successful embryo implantation (Piccinni and others 2001). An increase in the levels of Th2 cytokines (such as IL-10), accompanied by a parallel decrease in the levels of Th1 cytokines (TNF-α), is observed in successful pregnancies (Marzi and others 1996). In addition, IL-1β has also been implicated in embryo implantation, and it can induce the secretion of endometrial leukemia inhibitory factor, which is critical for implantation (Arici and others 1995). The possible mechanism of LPS-induced implantation failure might be due to the production of cytokines by activated macrophages, which enhanced the cytotoxicity of uterine natural killer cells. Priming of macrophages with LPS resulted in the release of proinflammatory cytokines, including IL-1β and TNF-α, which are involved in the delicate immune balance at the maternal–fetal interface (Kauma 2000; Zhong and others 2008). In our study, we found that endometrial IL-1β and TNF-α levels were significantly increased and IL-10 level was slightly decreased in LPS-challenged mice. Uterine macrophages represent a major subset of leukocyte during pregnancy, and its aberrant activity is linked to poor pregnancy outcome (Nagamatsu and Schust 2010). Thus, we also investigated the effect of IFN-τ on macrophages. In the in vitro experiments, IL-1β and TNF-α expression was significantly increased in LPS-stimulated macrophages, which was similar to the in vivo results. Meanwhile, IL-10 expression was not influenced by LPS. The aberrant production of IL-1β and TNF-α might be responsible for the implantation failure. IFN-τ treatment significantly reduced IL-1β and TNF-α levels and promoted IL-10 secretion, ultimately restoring the Th1/Th2 balance. NF-κB is involved in LPS-induced inflammatory responses, and its activation promotes the production of inflammatory cytokines (Wu and others 2016b). Western blot results showed that IFN-τ treatment obviously inhibited the phosphorylation of NF-κB p65. Immunofluorescence analyses demonstrated that IFN-τ treatment suppressed the nuclear translocation of NF-κB p65 in macrophages, in accordance with the Western blot results.
In addition, MHC is considered another important regulatory factor at the maternal–fetal interface, and the formation of MHC can cause immune rejection and be detrimental to pregnancy success. Several lines of evidence suggest that the regulation of MHC class I genes is essential for protecting the embryo from immunological attack (Low and others 1990; Al Naib and others 2011). Past research has also found that IFN-τ could silence the expression of MHC class I genes in the uterus of the pregnant sheep so as to repress maternal immune rejection responses to the conceptus (Choi and others 2001). Some cytokines, such as IFN-γ and IL-1β, have been reported to increase the expression of MHC class I genes (such as RT1-A) during early pregnancy in rats (Sun and others 2005; Xia and others 2006). Thus, we also examined the expression of mouse MHC class I genes (H-2K and H-2D) in the uterus. Intriguingly, our results found that H-2K and H-2D expression was increased by LPS but decreased by IFN-τ. The immunohistochemical results showed the same results. Taken together, we hypothesized that IFN-τ downregulates MHC class I genes expression, perhaps by inhibiting the production of inflammatory cytokines.
In conclusion, our findings demonstrate that IFN-τ has a protective effect in LPS-induced implantation failure by promoting embryo implantation. The possible mechanisms for the protective role of IFN-τ involve inhibiting inflammatory cytokines and MHC class I genes and upregulating IL-10. However, further studies are necessary to elucidate fully the effects and mechanisms of IFN-τ in the context of the complex regulatory network during embryo implantation.
Footnotes
Acknowledgments
We thank American Journal Experts for its linguistic assistance during the preparation of this article. This study was supported by the National Natural Science Foundation of China (No. 31472254 and 31772816).
Author Disclosure Statement
No competing financial interests exist.