
Abstract
Interferon-α (IFN-α) resistance is a major hurdle in the treatment of hepatocellular carcinoma (HCC). Signal transducers and activators of transcription 1 (STAT1) play a key role in exerting the antiproliferative and proapoptotic effects of IFN-α on tumors. In this study, we aimed to investigate whether resveratrol can promote IFN-α-induced growth inhibition and the apoptosis on HCC cells through the SIRT/STAT1 pathway. We found that IFN-α induced growth inhibition and apoptosis of SMMC7721 cells, and the effects could be significantly enhanced and blocked by resveratrol and EX527, respectively. Resveratrol not only activated SIRT1 but also induced phosphorylation of STAT1. Further study revealed that ablation of STAT1 reduced the combined antitumor effects of IFN-α and resveratrol, lowered the rate of apoptosis, and improved the viability of SMMC7721 cells. Whereas STAT1 overexpression strengthened the combined antitumor effects of resveratrol and IFN-α. Our findings suggest a novel strategy of using resveratrol to enhance the response of HCC to IFN-α treatment through the SIRT/STAT1 pathway.
Introduction
H
Interferons (IFNs) have been used with varying effectiveness as proapoptotic agents in the treatment of malignancies, and there is increasing evidence of the effectiveness of IFNs in the treatment and prevention of HCC (Ceballos and others 2014). IFNs, in combination with chemotherapeutics or agents such as proteasome inhibitors, could prevent HCC recurrence and improve survival (Sun and others 2006; Lee and others 2013; Li and others 2013). As an important therapeutic cytokine that exerts potent antineoplastic effects involving regulation of STAT1 gene, promyelocytic leukemia (PML) gene, and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) gene, interferon-α (IFN-α) is currently one of the important treatment choices for patients with HCC (Uka and others 2007; Herzer and others 2009; Wang and others 2012; Li and others 2013). Randomized studies have demonstrated a modest effect of IFN-α in advanced-stage HCC patients, and IFN-α is effective in reducing the incidence of HCC in patients with hepatitis B (HBV) and hepatitis C (HCV) viral infections (Sun and others 2006). However, a significant group of HCC patients show initial or acquired resistance to IFN-α therapy (Chung and others 2000; Tomimaru and others 2010). Thus, the need for novel strategies to help improve the efficacy of IFN-α is obvious. Sirtuins are highly conserved NAD-dependent deacetylases. Mammalian sirtuins have 7 homologs (SIRT1–7) that share a conserved catalytic domain as class III histone deacetylases (HDACs). Chloro-2,3,4,9-tetrahydro-1H-carbazole-1-carboxamide (EX527) is a specific inhibitor of SIRT1. Resveratrol is a natural agonist of SIRT1 and has been shown to exert anticancer effects (Andrade-Volkart and others 2017; Wilson and others 2017). Some studies demonstrate that resveratrol induces apoptosis of HepG2 cells through activation of the transcription factor FoxO3a and increases the expression of Bim protein (Liu and others 2016).
Several potential molecular pathways in HCC are targeted for therapeutic interventions such as the STATs (Stark and Darnell 2012; Yang and others 2016), epidermal growth factor receptor, retinoic acidinducible gene-I (Hou and others 2014), proline-rich tyrosine kinase 2 (Geng and others 2011), NF-κB, and p53 (Wu and others 2012; Zhou and others 2013). STAT1 plays an important role in promoting apoptotic cell death, and has been supported by the findings that growth-inhibiting and proapoptotic activities of IFNs are largely mediated by STAT1. STAT1-induced apoptosis is, in part, mediated by caspase-2 and -7 (Sironi and Ouchi 2004). Our previous study indicated that STAT1 exerts tumor-suppressive effects in HCC through the induction of cell cycle arrest and apoptosis (Chen and others 2013, 2015); however, the exact molecular mechanism underlying the suppressive effects has not been fully elucidated.
STAT1 can be activated by tyrosine phosphorylation by the members of the Janus tyrosine kinases (JAKs) in response to multiple cytokines and growth factors in addition to IFN-α, including interleukin-27 and -6 (IL-27 and IL-6) (Ho and others 2016). The importance of phosphorylation in regulating STAT1 functions has been a focus since the discovery of this transcription factor. However, increasing evidence indicates that acetylation of STAT1 is critically involved in IFNs and prolactin-initiated intracellular signaling (Wieczorek and others 2012; Zhuang 2013). A phosphorylation–acetylation switch is proposed to regulate STAT1 signaling. Some studies show that STAT1 acetylation regulates apoptosis by facilitating its binding to NF-κB and controls immune responses by suppressing STAT1 tyrosine phosphorylation (Krämer and others 2009). In contrast, others have shown that STAT1 signaling is not regulated by phosphorylation–acetylation switch (Antunes and others 2011). The functional role for acetylation/deacetylation of STAT1 in HCC has not yet been elucidated.
In this study, we tested the impact of resveratrol and IFN-α on proliferation, apoptosis, and cell cycle of SMMC7721 cell. Our results demonstrated that the naturally occurring phytoestrogen resveratrol can promote IFN-α-induced apoptosis and the growth inhibition of SMMC7721 cells through its effects on SIRT1/STAT1 pathway. Our data will provide new strategies for reducing tumor resistance to IFN-α through the targeting of STAT1.
Materials and Methods
Drugs and reagents
Resveratrol was purchased from Abcam, English. A 100 mM stock solution was prepared in dimethyl sulfoxide (DMSO) and further diluted in cell culture medium to 10, 20, and 30 μM working solutions. EX527 was purchased from Selleck. A 50 mM stock solution of EX527 was prepared in DMSO and further diluted in cell culture medium to 100, 200, and 300 nM working solutions. IFN-α was purchased from Abcam, English, and dissolved in phosphate-buffered saline (PBS), then diluted to 1,000, 2,000, and 3,000 U/mL with PBS medium.
Cell transfections and RNA interference
SMMC7721 cells (2 × 105/well) were cultured in 6-well plates at 70% confluence. The plasmids of pcDNA3.1 (EV), pcDNA3.1-STAT1, were transiently transfected into SMMC7721 cells using X-tremeGENE HP DNA transfection reagent (Roche, Mannheim, Germany). STAT1-specific or control siRNA was transiently transfected into SMMC7721 cells using X-tremeGENE siRNA transfection reagent (Roche), the transfection protocol used was described previously (Chen and others 2015), the STAT1-specifc and control siRNAs sequences are summarized in Table 1. SMMC7721 cells, overexpressing STAT1, or STAT1 siRNA were either left untreated or incubated with 1,000, 2,000, and 3,000 U/mL IFN-α only; IFN-α and resveratrol (10, 20, and 30 μM); or IFN-α and EX 527 (100, 200, and 300 nM). Cells were treated for the indicated time periods and then analyzed for flow cytometry, MTT assay, Western blot, and quantitative real-time reverse transcription-polymerase chain reaction (qRT-PCR).
STAT, signal transducers and activators of transcription.
Analysis of apoptosis and cell cycle
Cells were harvested and then stained using the Annexin V-FITC Apoptosis Assay Kit (Bestbio, Shanghai, China). The stained cells were analyzed by flow cytometry (BD FACSAria, R&D). Annexin V-positive and propidium iodide-negative cells were counted as apoptotic cells. For the cell cycle analysis, the cells were resuspended in 1 mL PBS solution with 40 μg propidium iodide (Sigma) and 100 μg RNase A (Sigma) for 30 min at 37°C then analyzed with the BD Accuri C6 system (Becton Dickinson). The distribution of cells in different phases of the cell cycle was calculated using the Modifit LT software. The analysis protocol was followed as described previously (Chen and others 2015).
MTT assay
Cells were seeded in 96-well plates at a density of 1 × 104 cells per well in 100 μL complete medium. After incubation with IFN-α, IFN-α, and resveratrol or IFN-α and EX527, 10 μL MTT solution (5 mg/mL) was added, and cells were incubated for an additional 4 h. Subsequently, 100 μL MTT solubilization buffer was added to the wells. After a 10 min mixing period, the absorbance was analyzed at 570 nm using a microplate reader. The background absorbance at 690 nm was subtracted from the 570 nm measurement. Each experiment was performed in triplicate and the mean value was calculated.
Quantitative real-time reverse transcription-polymerase chain reaction
Real-time PCRs were performed using 2 × SYBR® Premix Ex Taq™ II (Takara, Dalian, China), and amplified signals were detected using the ABI PRISM 7300 Real-Time PCR system (ABI), following the protocol as described previously (Chen and others 2015). The relative target mRNA levels were determined using the 2ΔΔCt method. The primer sequences are summarized in Table 1.
Western blot analysis
Samples containing equal amounts of protein (40 μg) were resolved by 10% SDS-PAGE and transferred to nitrocellulose membranes. After blocking for 2 h at room temperature with TBS-T (0.1 M Tris, 0.9% NaCl, and 0.05% Tween-20 at pH 7.5) containing 5% skim milk, membranes were probed at 4°C overnight with rabbit anti-STAT1 (1:2,000; Bioworld Technology), rabbit anti-p-STAT1 Y701 (1:400) (Beijing Biosynthesis Biotechnology Co., Beijing, China), rabbit anti-SIRT1 (1:1,000; Cell Signaling Technology), and mouse antiactin (1:8,000). Proteins were detected by exposing the blots to X-ray film (Kodak).
Coimmunoprecipitation
Cells were washed 3 times with PBS and lysed in Western/IP buffer (Beyotime Biotechnology, China), supplemented with 1 mM phenylmethylsulfonyl fluoride (PMSF, Beyotime Biotechnology) for 15 min. Whole cell extracts were collected by centrifugation (10,000 g) at 4°C for 15 min and used for immunoprecipitation and Western blot. For immunoprecipitation, cell lysates (100 μg) were precleared with 20 μL protein A + G agarose beads (Beyotime Biotechnology) at 4°C for 1 h. Supernatants were transferred to new tubes after centrifugation (10,000 g) at 4°C for 5 min. Supernatants were incubated with 50 μL protein A + G agarose beads for 1 h before adding 1 μg of anti-STAT1 antibody (1:400; Bioworld Technology) or anti-SIRT1 antibody (1:200; Cell Signaling Technology) and incubating overnight at 4°C. Beads were collected by centrifugation (3,000 g) at 4°C for 5 min and washed 3 times with PBS. The interaction between STAT1 and SIRT1 was detected by anti-SIRT1 or anti-STAT1 antibody.
Statistical analysis
SPSS version 17.0 software was used for all statistical analyses. All of the results are expressed as the mean ± SD. Statistical analysis was performed using standard 2-way ANOVA for repeated measurements, and the χ2 test was used to analyze the flow cytometry data. P values <0.05 were considered statistically significant.
Results
IFN-α induced apoptosis and increased the expression of p-STAT1 and SIRT1 of SMMC7721 cells
SMMC7721 cells were cultured to 70%–80% confluence, and incubated in serum-free medium for 10 h, cells were exposed to IFN-α at the concentrations of 0 (control), 1,000, 2,000, and 3,000 U/mL with 10% fetal calf serum for 1, 2, 3, and 4 h. The cell viability was assessed using MTT colorimetric assay (R&D). The results of the MTT assay indicated that IFN-α caused growth inhibition of SMMC7721 cells compared with the control, and it had the strongest inhibitory effect at the concentrations of 3,000 U/mL for 3 h (Fig. 1A). Flow cytometry analysis showed that apoptosis was more prevalent after incubation with 3,000 U/mL IFN-α (18.4%) than in control (5.7%) and other concentrations (2,000 U/mL, 13.6%; 1,000 U/mL, 9.1%) (Fig. 1B, D). Cell cycle analysis showed that SMMC7721 cells had a lower proportion of cells in G2 phase after incubation with IFN-α (3,000 U/mL, 8.20%; 2,000 U/mL, 8.02%; and 1,000 U/mL, 11.83%) than control cells (15.80%) (Fig. 1C, E). We measured the expression of STAT1, p-STAT1, and SIRT1 proteins after stimulation with IFN-α. The results from these experiments indicated that stimulation of SMMC7721 cells with IFN-α resulted in significant phosphorylation of STAT1. The total STAT1 and SIRT1 proteins expression levels were also significantly increased (Fig. 1F). We also monitored the mRNA expression of these proteins using qRT-PCR and obtained similar results (Supplementary Fig. S1A; Supplementary Data are available online at
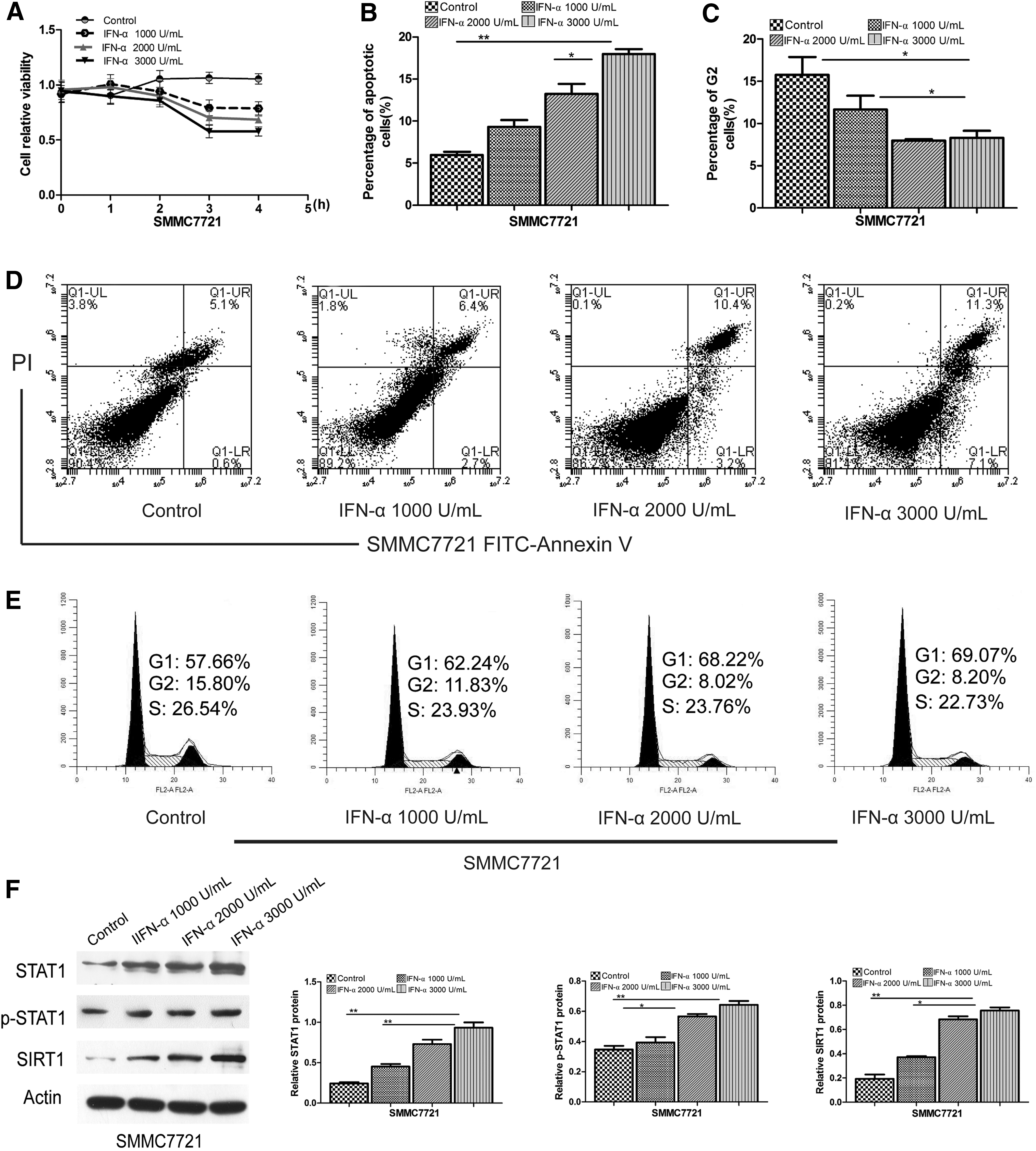
IFN-α-induced growth inhibition and apoptosis of SMMC7721 cells. SMMC7721 cells were left untreated (control) or treated with IFN-α at the concentrations of 1,000, 2,000, and 3,000 U/mL.
Resveratrol promoted IFN-α-induced antitumor effects of SMMC7721 cells
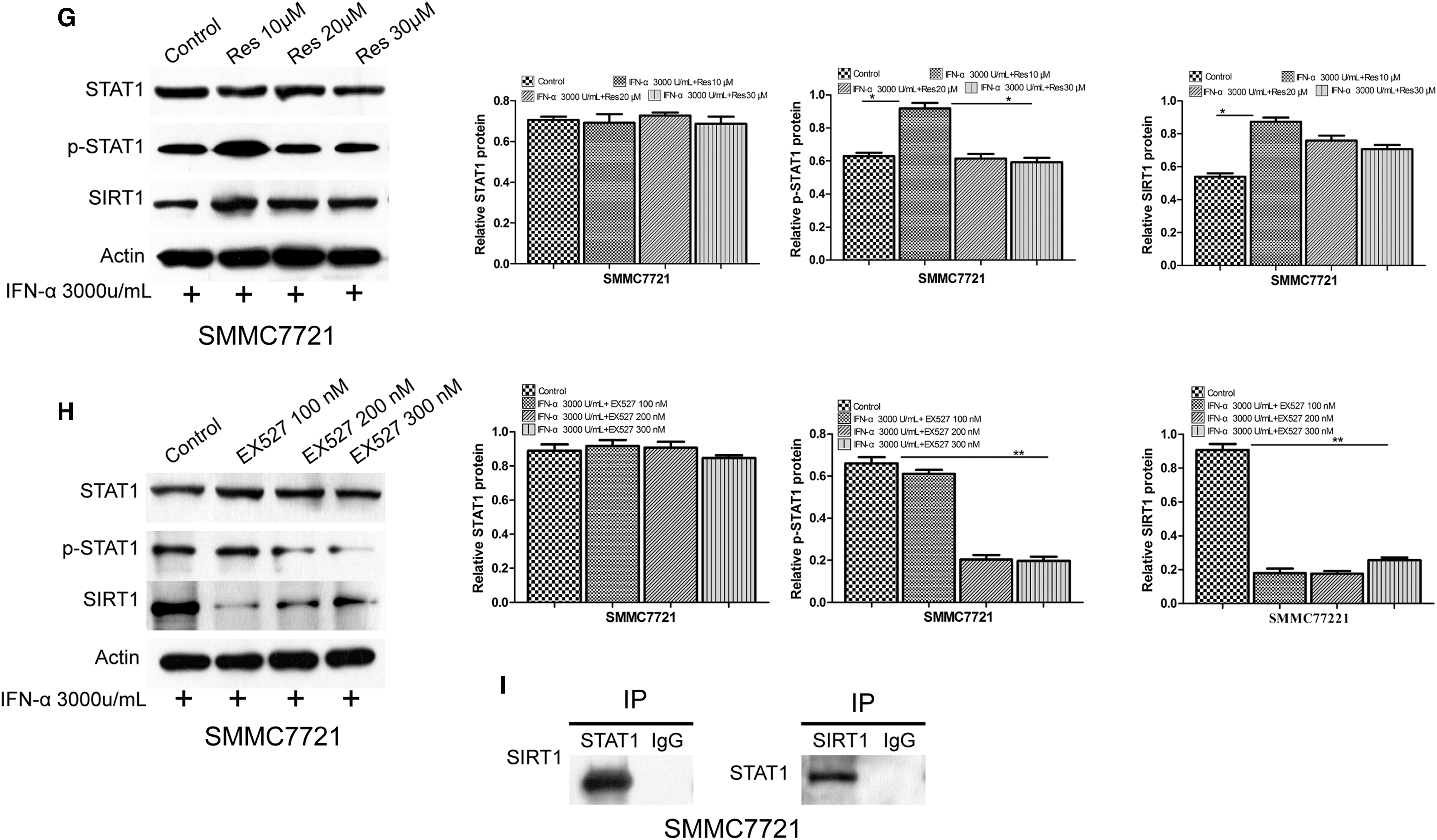
SMMC7721 cells were treated with 3,000 U/mL IFN-α and resveratrol (10, 20, and 30 μM) or EX527 (100, 200, and 300 nM) for 1, 2, 3, and 4 h. MTT assay showed that the combined treatment of IFN-α (3,000 U/mL) and resveratrol (10 μM) prominently suppressed cell viability compared with the control and other agents (Fig. 2A). In contrast, the inhibitory effect was significantly reduced in the presence of EX527 (Fig. 2B). Flow cytometry analysis showed that the combined treatment of IFN-α and resveratrol induced a significant increase in the levels of apoptosis (26.2%) compared with control (6.5%), IFN-α (16.1%), IFN-α, and EX527 (9.6%) (Fig. 2C, E). Cell cycle analysis showed that G2 phase was remarkably decreased after the combined treatment (3.42%) compared with control (17.47%), IFN-α (9.87%), IFN-α, and EX527 (12.05%) (Fig. 2D, F). We assayed the interaction of SIRT1 and STAT1 by immunoprecipitation with anti-STAT1 or anti-SIRT1 antibody, and Western blotting with anti-SIRT1 or anti-STAT1, respectively. The results showed an interaction between SIRT1 and STAT1 (Fig. 2I). We measured the expression of STAT1, p-STAT1, and SIRT1 proteins in response to IFN-α stimulation with resveratrol or EX527. When IFN-α was combined with resveratrol, p-STAT1 and SIRT1 were significantly enhanced compared with IFN-α treatment alone (Fig. 2G). In contrast, the expression levels of p-STAT1 and SIRT1 were significantly decreased when IFN-α was combined with EX527 (Fig. 2H). Similar results were also observed for the mRNA expression of these proteins (Supplementary Fig. S1B, C).
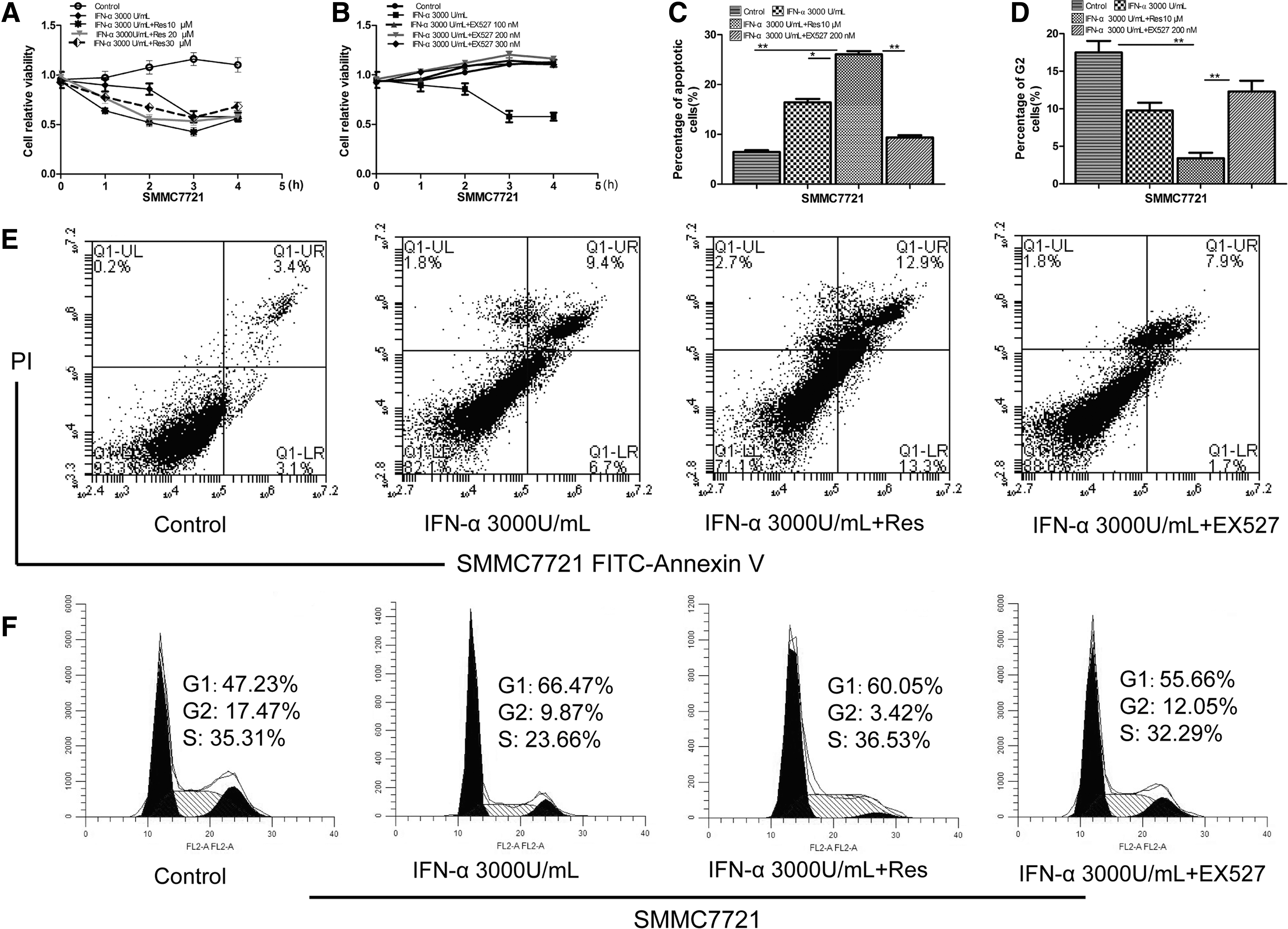
Resveratrol promoted IFN-α-induced antitumor effects and upregulated the expression of p-STAT1 and SIRT1. SMMC7721 cells were treated with 3,000 U/mL IFN-α and resveratrol (10, 20, and 30 μM) or EX527 (100, 200, and 300 nM) for 1, 2, 3, and 4 h. Cell proliferation was quantified by MTT assay.
STAT1 overexpression strengthened the combined antitumor effects of IFN-α and resveratrol
SMMC7721 cells were engineered to transiently express high levels of a recombinant plasmid encoding STAT1 sequence (pcDNA3.1-STAT1) or control empty vector pcDNA3.1 (EV). The levels of STAT1 expression were determined by qRT-PCR (Supplementary Fig. S2A) and Western blot assays (Fig. 3A). We measured the expression of STAT1, p-STAT1, and SIRT1 proteins in STAT1 overexpression cells after IFN-α stimulation with or without resveratrol or EX527. Western blot analysis showed that p-STAT1 and SIRT1 proteins were significantly increased after treatment with IFN-α and resveratrol, whereas the expression levels of p-STAT1 and SIRT1 were significantly decreased after treatment with IFN-α and EX527 (Fig. 3B, C). Similar results were also obtained for the mRNA expression of these proteins (Supplementary Fig. S2B). The results of the MTT assay showed that overexpression of STAT1 caused significant growth inhibition after treatment with IFN-α and resveratrol compared with the control, IFN-α, and EX527 (Fig. 3F). Flow cytometry analysis showed that STAT1 overexpression after treatment with IFN-α and resveratrol induced significant apoptosis (30.05%) compared with the control (12.1%), IFN-α (22.3%), IFN-α, and EX527 (12.0%) (Fig. 3D, E). Cell cycle analysis showed that G2 phase was remarkably decreased (2.18%) after treatment with IFN-α and resveratrol compared with the control (8.42%), IFN-α (5.56%), IFN-α, and EX 527 (7.17%) (Fig. 3G, H).
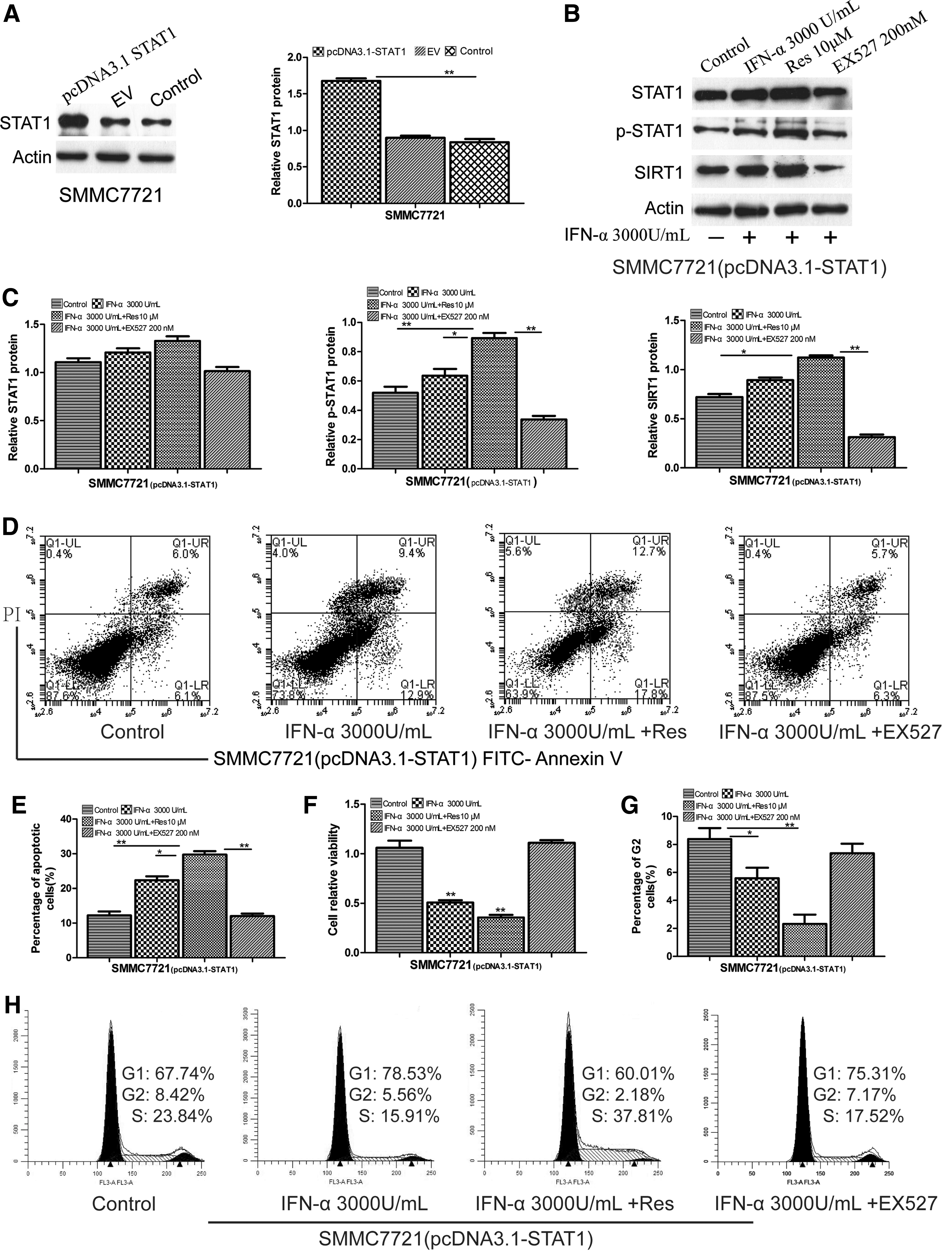
The impact of STAT1 overexpression on the combined antitumor effects. SMMC7721 cells were engineered to transiently express high levels of a recombinant plasmid encoding STAT1 sequence (pcDNA3.1-STAT1).
STAT1 knockdown inhibited the combined antitumor effects of IFN-α and resveratrol
We knocked down STAT1 in SMMC7721 cells by siRNA transfection, and STAT1 expression levels were evaluated by Western blot (Fig. 4A) and qRT-PCR (Supplementary Fig. S2C). The expression levels of STAT1, p-STAT1, and SIRT1 were significantly decreased in STAT1 siRNA cells after treatment with IFN-α, IFN-α, and resveratrol/EX527 (Fig. 4B, C). The mRNA expression levels of these proteins were similar to those obtained by Western blot (Supplementary Fig. S2D). MTT assay results suggested that STAT1 knockdown reduced the combined antitumor effects of resveratrol and IFN-α (Fig. 4G). Flow cytometry analysis showed that STAT1 knockdown lowered the rate of apoptosis (3.4%) compared with IFN-α (6.4%), IFN-α and resveratrol (6.7%), and IFN-α and EX527 (4.4%) (Fig. 4D, E). Cell cycle analysis showed that G2 phase was remarkably increased (17.85%) compared with IFN-α (11.74%), IFN-α and resveratrol (11.05%), and IFN-α and EX527 (14.04%) (Fig. 4F, H).
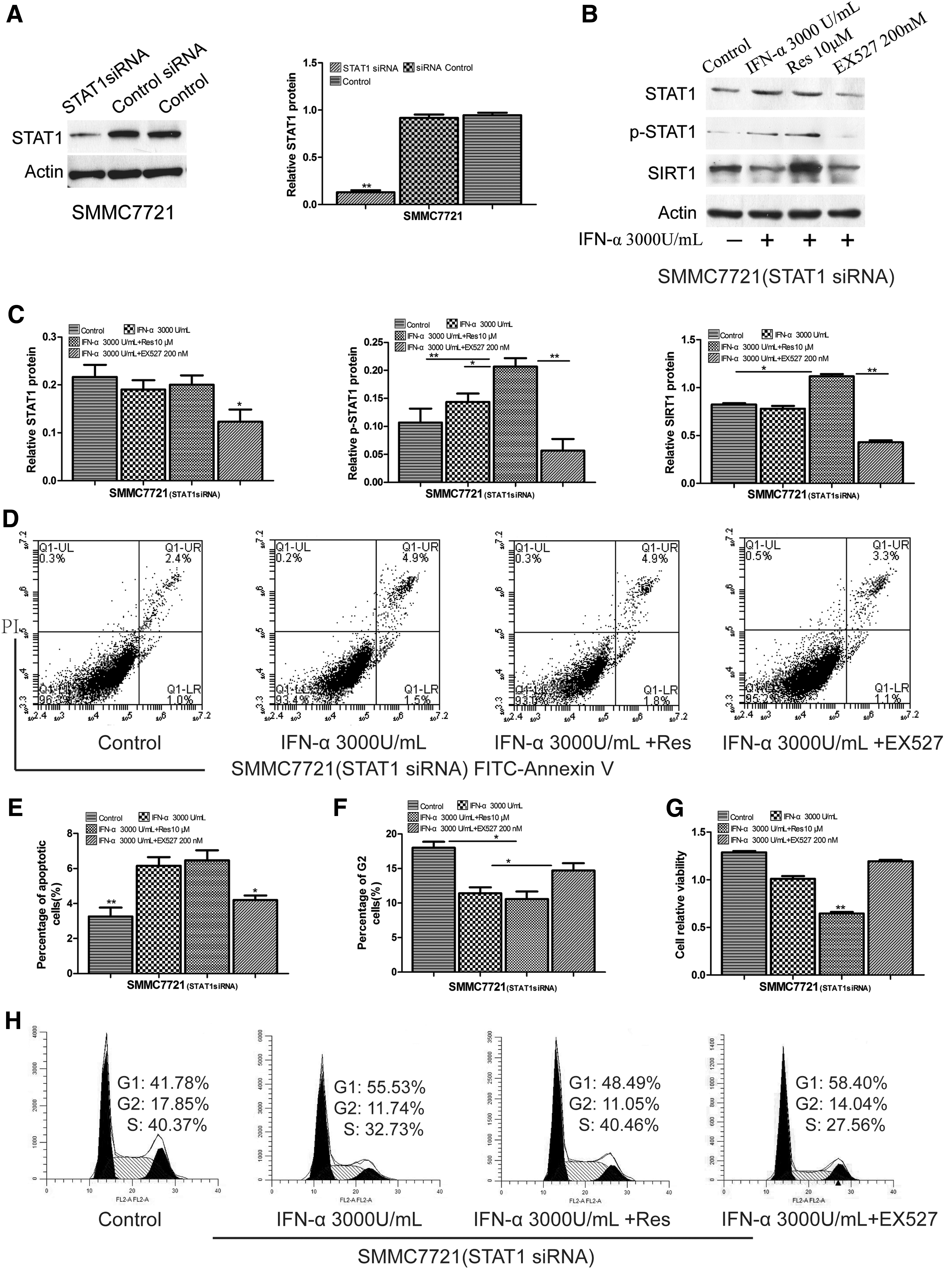
The impact of knockdown STAT1 on combined antitumor effects. SMMC7721 cells were transfected with STAT1 siRNA or control siRNA.
Discussion
IFNs are pleiotropic cytokines that have been shown to be important regulators of cell growth. IFN-α has been recognized to harbor therapeutic potential in prevention and treatment of HCC. However, the therapeutic effects of IFN-α on HCC cells appear to be tissue- and cell-type specific, ranging from potent apoptosis induction to complete apoptosis resistance. Cells resistance to IFN-α is a major hurdle in the management of HCC, and strategies to help improve the efficacy of IFN-α are greatly needed to achieve a better control of HCC. In this study, similar effects of growth inhibition and apoptosis were observed after treatment with IFN-α, but only 18.4% of SMMC7721 cells underwent apoptosis after treatment with IFN-α (3,000 U/mL) (Fig. 1B, D). However, IFN-α-induced growth inhibition and apoptosis were significantly enhanced when IFN-α was combined with 10 μM resveratrol (Fig. 2C, E). In contrast, the IFN-α-induced effects of apoptosis were downregulated when SMMC7721 cells were treated with IFN-α and EX527 (Fig. 2C, E). Our finding may provide a potential approach for clinical application to improve the efficacy of IFN-α on control HCC.
The JAK-STATs signal pathway is a major mechanism for the antiproliferative and proapoptotic effects of IFN-α. STAT1 signaling regulates the expression of important genes controlling cell growth, differentiation, apoptosis, and immune functions. STAT1 cascade in tumor is controversial, despite strong data indicating that STAT1 downregulation was most prominent in the tumor cells themselves when compared with the surrounding stroma and infiltrating lymphocytes (Chan and others 2012). STAT1 controls antitumorigenic effects in part by the upregulation of cyclin-dependent kinase inhibitor 1A, IFN-regulatory factor 1, and p53 (Townsend and others 2004) and downregulation of the B cell chronic lymphocytic leukemia/lymphoma 2 family. In contrast, other groups have found that in certain cellular contexts, STAT1 may mediate tumor cell growth. Overexpression of IFN/STAT1 is associated with poor prognosis in different types of cancer (Khodarev and others 2007, 2009). Serine phosphorylation, acetylation, methylation, and sumoylation are involved in STAT1 posttranslational modifications. Lysine acetylation has recently been appreciated as a novel mechanism regulating signal transduction events relying on STAT proteins. STAT1 acetylation depends on the balance between STAT1-associated HDACs and histone acetyltransferases such as CREB-binding protein. Increasing evidence indicates that acetylation/deacetylation of STAT1 is critically involved in IFNs and prolactin-initiated intracellular signaling (Wieczorek and others 2012; Zhuang 2013).
SIRT1 is a highly conserved class-III HDAC that depends on NAD+ as a cofactor to catalyze the deacetylation of histone and nonhistone proteins (Vaziri and others 2001). SIRT1 is known to regulate a host of cellular events through its deacetylase activity and has numerous beneficial effects on a variety of human diseases. Moreover, the recent reports note that SIRT1 signal is an emerging factor for many biological processes such as inflammatory response and tumorigenesis (Kim and others 2016). However, its role in tumorigenesis is somewhat controversial, as some reports have indicated that it acted as a tumor promoter, whereas others have found that it has tumor suppressor function (Yang and others 2013; Zhang and others 2017). Resveratrol is the most effective natural agonist of SIRT1, exhibiting a wide range of biological and pharmacological properties, including anticancer, anti-inflammatory, antioxidant, and vasorelaxant activities through regulation of cell signaling pathways and autophagy (Qureshi and others 2012; Ji and others 2015; Farooqi and others 2018). Recent studies have proposed that resveratrol has a direct inhibitory effect on HCC through Gli-1 and Akt/FoxO3a/Bim pathway (Liu and others 2016).
In this study, resveratrol could upregulate the expression of SIRT1 and p-STAT1, enhancing IFN-α-induced growth inhibition and apoptosis of SMMC7721 cells (Fig. 2). Coimmunoprecipitation and Western blot demonstrated functional and physical interactions between STAT1 and SIRT1 (Fig. 2I). These results indicated that SIRT1 may be a modulator of resveratrol and IFN-α-induced growth inhibition and apoptosis of HCC cells. We transiently transfected STAT1 expression vectors and STAT1-specific siRNA into SMMC7721 cells to investigate the function of STAT1 on resveratrol/IFN-α-induced growth inhibition and apoptosis. Our data showed that ablation of STAT1 reduced the combined antitumor effects of resveratrol and IFN-α (Fig. 4G), and lowered the rate of apoptosis (Fig. 4D, E). In contrast, STAT1 overexpression strengthened the combined antitumor effects (Fig. 3). The SIRT1/STAT1 interactions suggest that STAT1 may play an important role in regulating resveratrol-activated SIRT1 during HCC cell growth inhibition and apoptosis.
In conclusion, our data indicate that resveratrol enhances IFN-α-induced growth inhibition and apoptosis of SMMC7721 cells through the SIRT1/STAT1 pathway. The activation of SIRT1 through resveratrol induces the upregulation of p-STAT1, thereby sensitizing more SMMC7721 cells to IFN-α-induced apoptosis and growth inhibition. Our findings may provide a basis for the design of new therapeutic methods by using resveratrol to improve the response of HCC to IFN-α treatment, and may lay the groundwork for clinical application of IFN-α and resveratrol therapy of HCC. In addition, although the STAT1 is a key in the synergetic antitumor effects, STAT1 involvement in oncogenesis and its modification are more complex. Further studies are needed to disclose new possibly critical pathways that may be involved in the combined antitumor effect of IFN-α and resveratrol.
Footnotes
Acknowledgments
This work was supported by China Association of Higher Education (Grant No. 16YB161); Society of Medical Education, Chinese Medical Association (Grant No. 2016-RC006); and Science and Technology Research Project of Education Department of Heilongjiang Province of China (Grant No. 12531271).
Author Disclosure Statement
No competing financial interests exist.