
Abstract
Tumor vasculature plays a central role in tumor progression, making it an attractive therapeutic target. In this study, we explore the antiangiogenic potential of our melanoma gene therapy approach combining interferon β (IFNβ) and p19Arf gene transfer. Since these proteins are modulators of tumor vasculature, we explore the impact of IFNβ and p19Arf gene transfer on murine endothelial cells (tEnd). Adenovirus-mediated gene transfer of p19Arf to tEnd cells inhibited proliferation, tube formation, migration, and led to increased expression of genes related to the p53 cell death pathway, yet IFNβ gene transfer had no significant impact on tEnd viability. Alternatively, tEnd cells were exposed to the factors generated by transduced B16 (mouse melanoma) cells using either coculture or conditioned medium. In either case, transduction of B16 cells with the IFNβ vector, whether alone or in combination with p19Arf, resulted in endothelial cell death. Strikingly, treatment of tEnd cells with recombinant IFNβ did not induce death, demonstrating that additional factors produced by B16 cells contributed to the demise of tEnd cells. In this work, we have shown that our melanoma gene therapy strategy produces desirable negative effects on endothelial cells, possibly correlating with antiangiogenic activity.
Introduction
Melanoma is the most aggressive and deadliest form of skin cancer (Siegel and others 2014). The etiology of melanoma involves environmental, phenotypic, and genetic phenomena (Gandini and others 2005; Higgins and others 2015) and the tumor itself is composed of transformed melanocytes and a variety of stromal cells that provide support and even promote progression (Villanueva and Herlyn 2008). Particularly important are the tumor blood vessels that control the supply of nutrients (Dudley 2012), maintain blood flow, and regulate leukocyte trafficking, including recruitment of myeloid cells needed to promote angiogenesis (Schmid and Varner 2007). The blood vessels are composed of smooth muscle cells, pericytes, and endothelial cells, which are especially active in the amplification of cancer growth and its spread (Michiels 2003). Because the endothelial cells play such a critical role, we have chosen these as the subject of our study aimed at further developing a cancer gene therapy approach.
We have employed interferon β (IFNβ) and p19Arf for cancer gene therapy due to their multiple antineoplastic activities. IFNβ is a cytokine well known for its role in antiviral defense, immune modulation, and induction of cell death (Chawla-Sarkar and others 2003; Schoggins and Rice 2011; Medrano and others 2017). The p19Arf tumor suppressor protein (p19Arf in mice, p14Arf in humans) is best known for its role in preventing the interaction of p53 with MDM2, resulting in p53 activation and accumulation (Kamijo and others 1998; Zhang and others 1998; Milojkovic and others 2013). In fact, critical interaction between the IFN and p53/Arf pathways has been shown to aid antiviral defense and cell killing (Takaoka and others 2003; Sandoval and others 2004).
In addition, IFNβ and p19Arf each carry out antiangiogenic functions. IFNβ acts on endothelial cells, reducing the production of proangiogenic factors and inducing cell death (Bracarda and others 2010). Angiogenesis is negatively modulated by p19Arf through p53 dependent and independent mechanisms (Teodoro and others 2007; Kawagishi and others 2010; Ulanet and Hanahan 2010). Whether individually or in tandem, we expect that the use of IFNβ and p19Arf would impact the tumor vasculature by inhibiting angiogenesis and, possibly, destroying existing vessels.
Melanomas typically retain p53 in the wild-type form (Giglia-Mari and Sarasin 2003), possibly serving as a molecular target for treatment. To this end, we have developed adenoviral vectors wherein high-level transgene expression is controlled by p53 in a nonreplicating Ad5 vector (Bajgelman and Strauss 2008) or Ad5 with arginylglycylaspartic acid modification (Hunger and others 2017). We have used these vectors to transfer the p19Arf and IFNβ genes resulting in enhanced melanoma cell death in vitro and in vivo (Merkel and others 2013), induction of a protective immune response in models of melanoma and lung carcinoma (Catani and others 2016; Medrano and others 2016), and a strong bystander effect between treated and untreated melanoma cells (Hunger and others 2017). However, our previous studies did not include evaluation of the tumor vasculature upon treatment with our gene therapy approach.
In this study, we show that AdRGD-PG (nonreplicating Ad5 with RDG tripeptide modification and transgene expression under the control of p53) expressing p19Arf inhibits migration, tube formation, and cell proliferation and also induces cell death consistent with apoptosis upon transduction of the murine endothelial cell line (tEnd). Exposure of tEnd to the factors produced by B16 cells after their transduction with the adenoviral vector encoding IFNβ revealed a paracrine effect that resulted in endothelial cell death. Collectively these data indicate that our vectors have a desirable negative impact on endothelial cell activity consistent with the inhibition of angiogenesis upon melanoma gene therapy.
Materials and Methods
Cell lines
The murine melanoma cell line B16F10 (B16) was kindly provided by Dr. Roger Chammas, Instituto do Câncer do Estado de São Paulo, Faculdade de Medicina, Universidade de São Paulo, SP, Brazil; tEnd was kindly provided by Dr. Cristina Maria Fernandes, Instituto Butantã, SP, Brazil; and HEK293A (adenovirus-transformed human embryonic kidney) cell line was obtained commercially (Invitrogen Life Technologies, Carlsbad, CA). The B16 and tEnd cell lines were maintained in RPMI (Life Technologies) and HEK293 line was maintained in Dulbecco's Modified Eagle Medium (DMEM) (Invitrogen Life Technologies) plus 5% fetal bovine serum (FBS; Life Technologies) and 1% antibiotic-antimytotic (Invitrogen Life Technologies), maintained at 37°C in a humidified atmosphere with 5% CO2.
Production and transduction of adenoviral vectors
The AdRGD-PG vectors (nonreplicating Ad5 with RGD fiber modification and transgene expression under the control of a p53-responsive promoter, termed PG) encoding enhanced green fluorescent protein (GFP), p19Arf, or murine IFNβ have been described in detail elsewhere (Hunger and others 2017). AdRGD-CMVLacZ (Mizuguchi and others 2001) was kindly provided by Dr. Hiroyuki Mizuguchi (Osaka University, Japan). The adenoviruses were produced in the HEK293A cell line and purified by an iodixanol ultracentrifugation as described previously (Peng and others 2006). The viruses were titrated with Adeno-X Rapid Titer Kit (Clontech, Mountain View, CA) and this biological titer was used when calculating the multiplicity of infection (MOI). For transduction, the MOI indicated in each assay was applied in DMEM or RPMI medium with 5% FBS and 1% antibiotic-antimytotic and maintained at 37°C at 5% CO2 atmosphere.
X-Gal staining
tEnd cells were plated in 6 well dishes, 1 × 105 cells/well, transduced with the Ad-LacZ vector at an MOI of 200 and incubated for 24 h. After this period, the cells were fixed with 2% paraformaldehyde/0.2% glutaraldehyde for 5 min. For X-Gal staining, the cells were first washed with 0.1 M phosphate buffer (pH 7.3) supplemented with 2 mM MgCl2, and then immersed in X-Gal staining buffer (0.1 M phosphate buffer, pH 7.3, supplemented with 2 mM MgCl2, 5 mM potassium ferrocyanide, 5 mM potassium ferricyanide, and 1 mg/mL of X-Gal) and incubated at 37°C for 16 h.
Flow cytometric detection of GFP positive cells
B16 and tEnd cells were seeded in 6 well dishes, 5 × 104 cells/well, transduced with Ad-GFP at an MOI of 200, and incubated for 24 h. After harvesting the cells, they were analyzed for GFP expression by flow cytometry (Attune; Invitrogen Life Technologies). The percentage of GFP positive cells was determined by the Attune software.
Immunofluorescence detection of p19Arf
B16 and tEnd cells were seeded onto 13 mm round glass coverslips, 5 × 104 cells/well, in 24-well dishes, transduced the next day with adenoviral vectors at an MOI of 200, and fixed with methanol 24 h later. Then, cells were blocked with bovine serum albumin (Sigma-Aldrich, St. Louis, MO), probed with a rabbit polyclonal antibody for p19Arf (AB-1; Cal-Biochem, San Diego, CA), followed by an Alexa488-labeled antirabbit secondary antibody (Molecular Probes, Eugene, OR). Hoechst 33258 (20 μg/mL; Molecular Probes) was used for nuclear staining. Cells were visualized by fluorescent microscopy (EVOS FL; Invitrogen Life Technologies).
Enzyme-linked immunosorbent assay for detection of IFNβ
Fifty thousand B16 or tEnd cells were transduced with the adenoviral vectors at an MOI of 200, and after incubation for 24 h the supernatants were collected. Detection of IFNβ was performed by ELISA (PBL Biomedical™, New Brunswick, NJ) following the manufacturer's protocol and using a Victor™ plate reader (Perkin-Elmer, Waltham, MA) where absorbance was measured at 450 nm.
Wound healing assay
The tEnd cells were seeded, 1 × 104 cells in 12-well plates, and 24 h later a pipette tip was used to create a wound in the confluent monolayer cells and transduction was performed using an MOI of 100 and cells were incubated in the presence of Mitomycin C (20 μg/mL). Photos were taken at 0 h (time of wounding and transduction) and 48 h later. Pictures were taken using an EVOS FL microscope (Invitrogen Life Technologies). Calculations were performed as follows: [(M 0 − M 48)/M0]*100 = X, where M 0 is the area of the wound at time 0, M 48 is the area 48 h later, and X is the resulting percentage of wound closure (area filled) due to migration.
Tubulogenesis assay
The tEnd cells were previously transduced using an MOI of 200 and incubated for 6 h. Then they were harvested, counted, and 1 × 104 cells/well were seeded in 24-well plates containing basic Matrigel (BD Biosciences, San Jose, CA). Photographs were taken 48 h after seeding of the cells and the number of tubes was quantified. Tubes were defined as closed tubes formed by >2 cells.
Clonogenic assay
The tEnd cells were transduced with the adenoviral vectors (MOI 200) and incubated for 6 h. Then, cells were harvested, counted, and 1 × 103 cells were seeded per 100 mm plate and incubated for 10 days. For the clonogenic assay with conditioned medium, 500 tEnd cells were seeded with conditioned medium in 6-well plates, and incubated for 10 days. After incubation time, cells were fixed with acetic acid–methanol (5%:95%) solution and stained with Trypan Blue. Colonies were quantified manually.
Cell viability assay
For the MTT assay, 1 × 103 cells were plated in 96-well dishes. The next day, they were transduced (MOI 200), then incubated for 48 h before determination of cell viability. Plates were incubated with 25 μL of MTT solution [5 mg/mL in 1 × phosphate-buffered saline (PBS)], 37°C during 4 h. The dish was then removed and the precipitate solubilized by the addition of 100 μL lysis buffer (20% sodium dodecyl sulfate in 50% dimethylformamide (DMF)/2% acetic acid, pH adjusted to 4.7) before analysis using Victor™ plate reader (Perkin-Elmer) at 570 nm.
Identification of hypodiploid, Annexin-V, and caspase 3/7 positive cells
B16 and tEnd cells were transduced with the viral vectors at an MOI of 200 or treated with medium containing recombinant interferon β (rIFNβ, 10 U/mL; Millipore, Temecula, CA, USA) and incubated for 48 h. Alternatively, cells were plated and the following day incubation in the presence of conditioned medium was initiated and then maintained for 48 h. For quantification of hypodiploid cells, cells were harvested, fixed with 70% ethanol, and stained with propidium iodide. For identification of phosphotydilserine exposed at the cell surface, fresh cells were labeled with Annexin-V-Alexa488 (Invitrogen Life Technologies) immediately upon harvesting. In this case, the Ad-LacZ vector was used as a control. For identification of caspase 3/7 activity, fresh cells were treated with CellEvent Caspase 3/7 Detection Reagent (Invitrogen Life Technologies). All analyses were performed by flow cytometry and associated software (Attune; Invitrogen Life Technologies).
Coculture assay
We first modified tEnd cells with the Lego-iT2 lentiviral vector (kindly provided by Kristoffer Rieken, University Medical Center Hamburg, Germany), which offers constitutive expression of the dTomato fluorescent protein to generate the tEndTO cell line. Cells were transduced with Lego iT2 vectors at an MOI of 1 in the presence of 8 μg/mL polybrene and expanded to a 100 mm dish. Then clones with the highest fluorescence levels were screened to isolate a cell line with fluorescence detectable in 100% of the cells. For the assay, 2 × 104 B16 cells were transduced (MOI 300), incubated for 6 h, and then washed to remove virus particles. We then seeded 1 × 104 tEndTO cells with the previously transduced B16 cells. The 2 types of cells were cocultured for 72 h. Photographs (EVOS FL; Invitrogen Life Technologies) were taken at 24 and 72 h after seeding. Quantification of each cell type was aided by use of the Image J program. Results from each time point in each independent experiment were normalized using the following formula: N T/N C = V, where N T is the average number of treated cells per field, N C is the average number of control cells per field, and V is the resulting value.
Production of conditioned medium
To produce conditioned medium, 5 × 104 B16 cells were transduced (MOI 300) and incubated for 6 h and then washed 4 times with 1 × PBS to remove virus particles. Fresh culture medium was then added and cells were incubated for 48 h. Then, medium was collected, centrifuged to separate out cellular debris, and stored at −80°C. The conditioned medium derived from B16 cells was named cm-B16 (nontransduced control), cm-B16-GFP (control transduced with Ad-GFP or Ad-LacZ), cm-B16-IFN (transduced with Ad-IFN), cm-B16-p19 (transduced with Ad-p19), and cm-B16-IFN+p19 (cotransduced with Ad-IFN and Ad-p19).
Reverse transcriptase-quantitative polymerase chain reaction analysis
The tEnd cells were plated in 6 well dishes, 5 × 104 cells/well, and transduced with the adenoviral vectors (MOI 200). After 24 h, total RNA was isolated using Trizol (Invitrogen Life Technologies) following the fabricant's instructions. Concentration of RNA was measured by absorbance at 260 nm. Quality of RNA was assessed by protein and salt concentration (absorbance 280 nm) and by visualizing the 18S and 28S ribosomal RNA bands in a 1% agarose gel. Primers (Supplementary Table S1) were designed and then analyzed using Primer-BLAST (
Statistical analysis
Data are expressed as the mean ± SD from at least 3 experiments. Statistical analysis was carried out using analysis of variance (ANOVA) followed by Tukey's test, significant at a level of P < 0.05. Analysis was performed by using GraphPad Prism 5.0.
Results
Validation of adenoviral vectors and efficiency of transduced cells
Expression of the GFP, LacZ, IFNβ, and p19Arf transgenes upon transduction with the recombinant adenoviral vectors was analyzed by fluorescence microscopy, staining with X-Gal, ELISA, or immunofluorescence, respectively (Supplementary Fig. S1). These assays demonstrate reliable expression of the transgenes when tEnd (mouse immortalized endothelial cells) or B16 (mouse melanoma cells) were transduced with our adenovectors. As seen in Supplementary Fig. S1, tEnd and B16 were transduced at similar levels (74.8% and 64.3%, respectively) as seen by detection of exogenous p19Arf. These data indicate that transduction of endothelial or melanoma cells with our RGD-modified vectors results in reliable gene transfer and transgene expression.
p19Arf inhibits angiogenic characteristics of transduced tEnd cells
We next transduced tEnd cells with our recombinant adenoviruses and analyzed indicators of angiogenic potential, specifically migration and tube formation. In the migration assay, the wound was formed and immediately followed by transduction and treatment with Mitomycin C (0 h). We observed significantly reduced migration when the tEnd cells had been transduced with Ad-p19 alone or in combination with Ad-IFN (Fig. 1A, B). In the tubulogenesis assay, cells were transduced with the vectors, incubated for 6 h, and then plated in wells containing Matrigel. After 48 h of incubation, the cells were photographed and the number of tubes formed was quantified. tEnd cells transduced with Ad-p19 alone or in combination with Ad-IFN showed significantly fewer tubes (Fig. 1C, D).
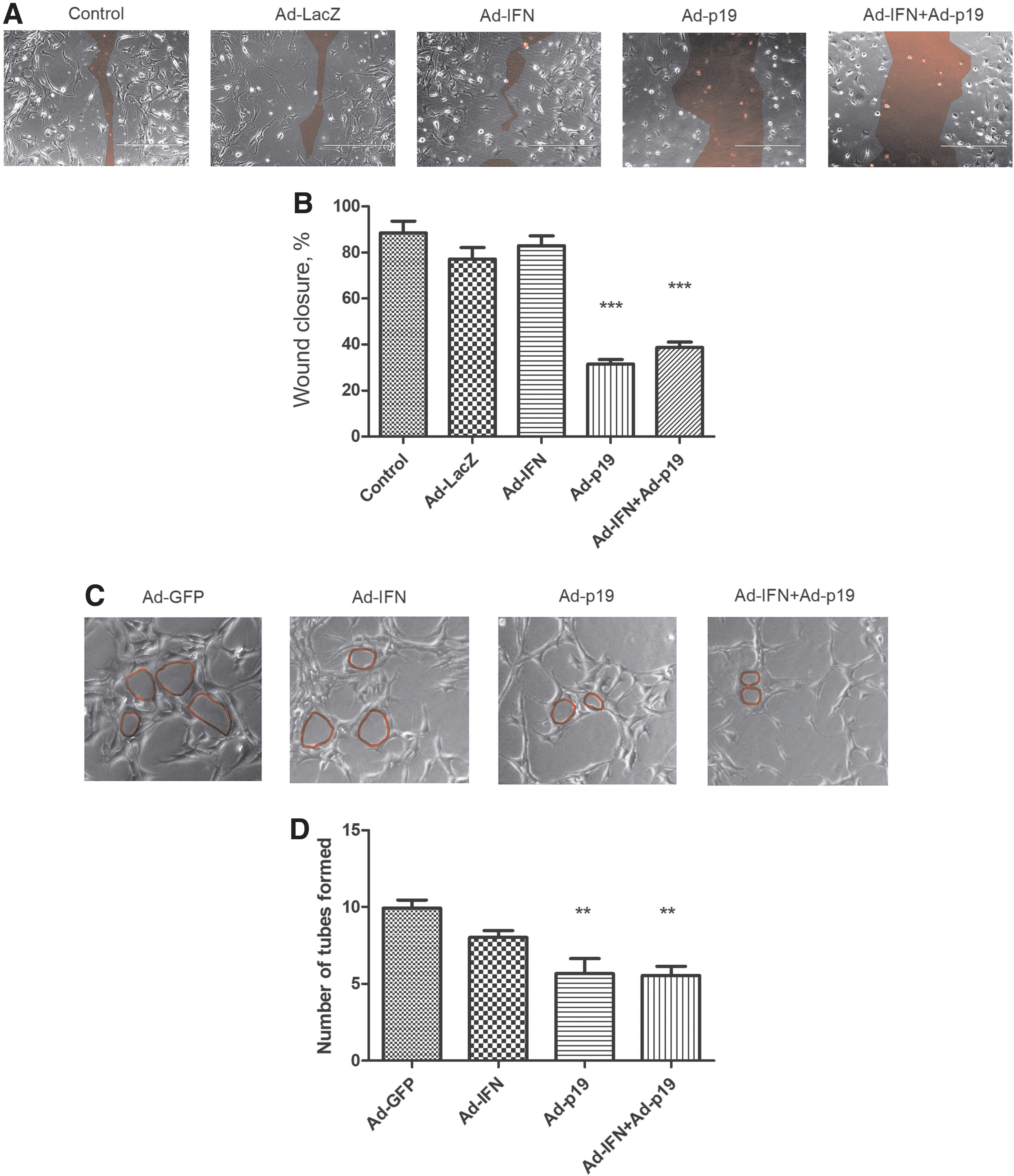
Analysis of angiogenic characteristics after transduction of enthelial cells.
Transduction of tEnd with Ad-p19 triggers cell death
The reduction in angiogenic potential already described may be due, in part, to alterations in cell proliferation and viability. To test this possibility, tEnd cells were transduced with an MOI of 200 before colony formation, viability, cell cycle, and apoptosis were evaluated. Only combined Ad-p19 and Ad-IFN gene transfer significantly reduced clonogenicity and viability (MTT assay) of tEnd cells, although a tendency was noted with individual Ad-p19, but not Ad-IFN, treatment (Fig. 2A–C). Interestingly, Ad-p19 either alone or in combination with Ad-IFN strongly induced accumulation of hypodiploid (sub-G1), Annexin-V positive cells, and activation of caspases 3/7 (Fig. 2D–F), indicating that Ad-p19 was sufficient to induce cell death by a mechanism consistent with an apoptosis. As expected, p19Arf gene transfer, but not IFNβ, was associated with the activation of p53-responsive genes, such as Cdkn1a (p21), Mdm2, and Puma (Bbc3) (Supplementary Fig. S2). These assays suggest that Ad-p19 is the dominant factor for inducing tEnd cell death upon transduction.
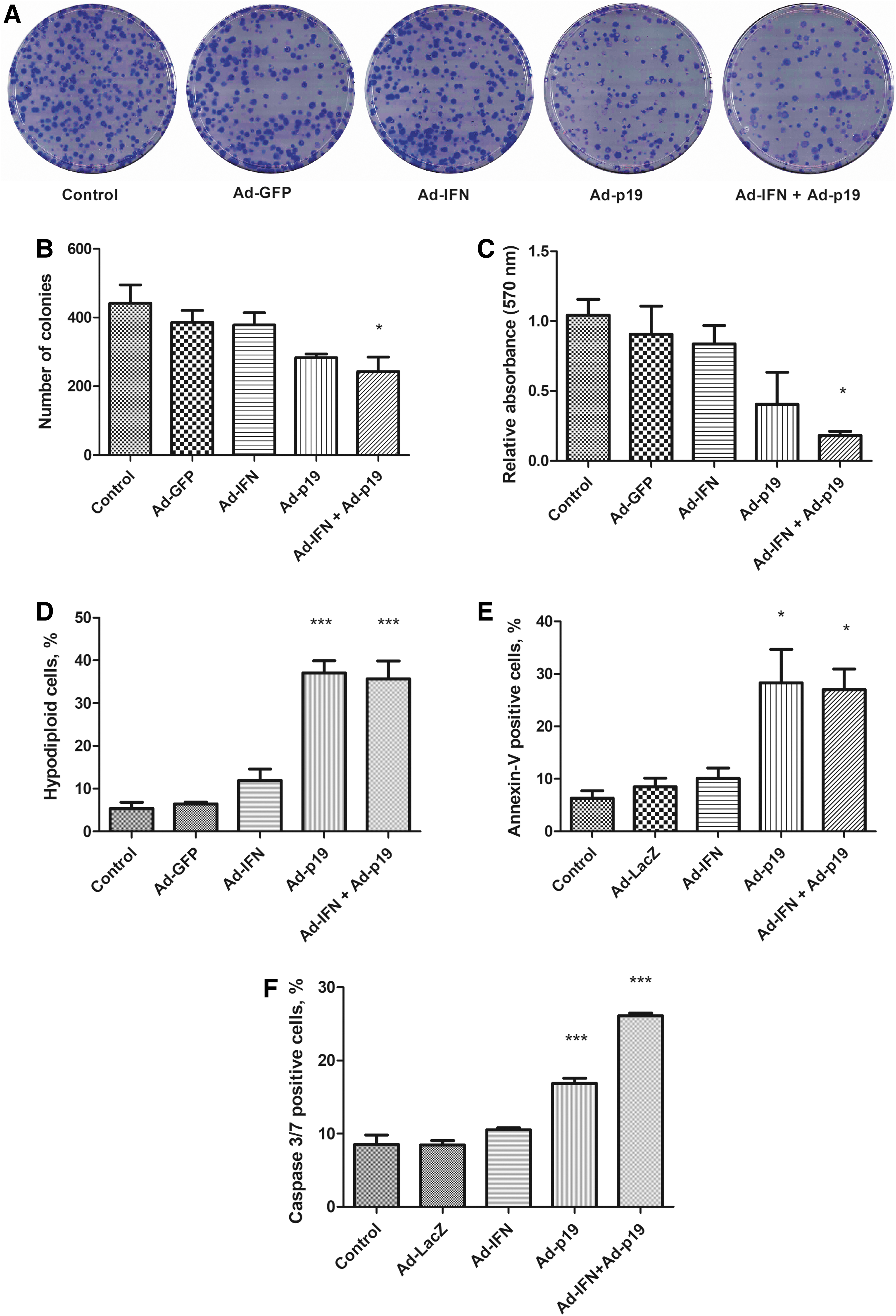
Assessment of viability and cell death upon transduction of tEnd cells.
Coculture of tEnd cells with transduced B16 melanoma cells reveals critical role of IFNβ
Up to this point in our study we have shown that direct transduction of tEnd cells with an adenoviral vector encoding p19Arf results in cell death, revealing a possible benefit of our melanoma gene therapy approach. We next examined whether the factors produced by the transduced tumor cells would have an impact on the endothelial cell line. For this, we used a coculture assay since this is a relevant means of simulating the interaction of different types of cells in the tumor microenvironment (Danhier and others 2012). To differentiate the 2 cell lines in the same plate, we labeled tEnd cells with constitutive expression of the dTomato fluorescent protein, generating the tEndTO cell line. In this assay, B16 cells were transduced (MOI 300, 6 h), washed, collected, and 1 × 104 of these cells were plated together with an equal number of tEndTO cells and observed for up to 72 h (Fig. 3A). Interestingly, the tEndTO population was reduced in the presence of B16 cells treated with Ad-IFN, whether alone or in combination with Ad-p19 (Fig. 3B). Consistent with our previous findings (Merkel and others 2013; Medrano and others 2016; Hunger and others 2017), transduction of B16 cells inhibits their proliferation when one or both of the genes were applied (Fig. 3C). This coculture assay shows that the secretome produced by B16 cells upon transduction with Ad-IFN, but not Ad-p19, can negatively impact the population of tEnd cells.
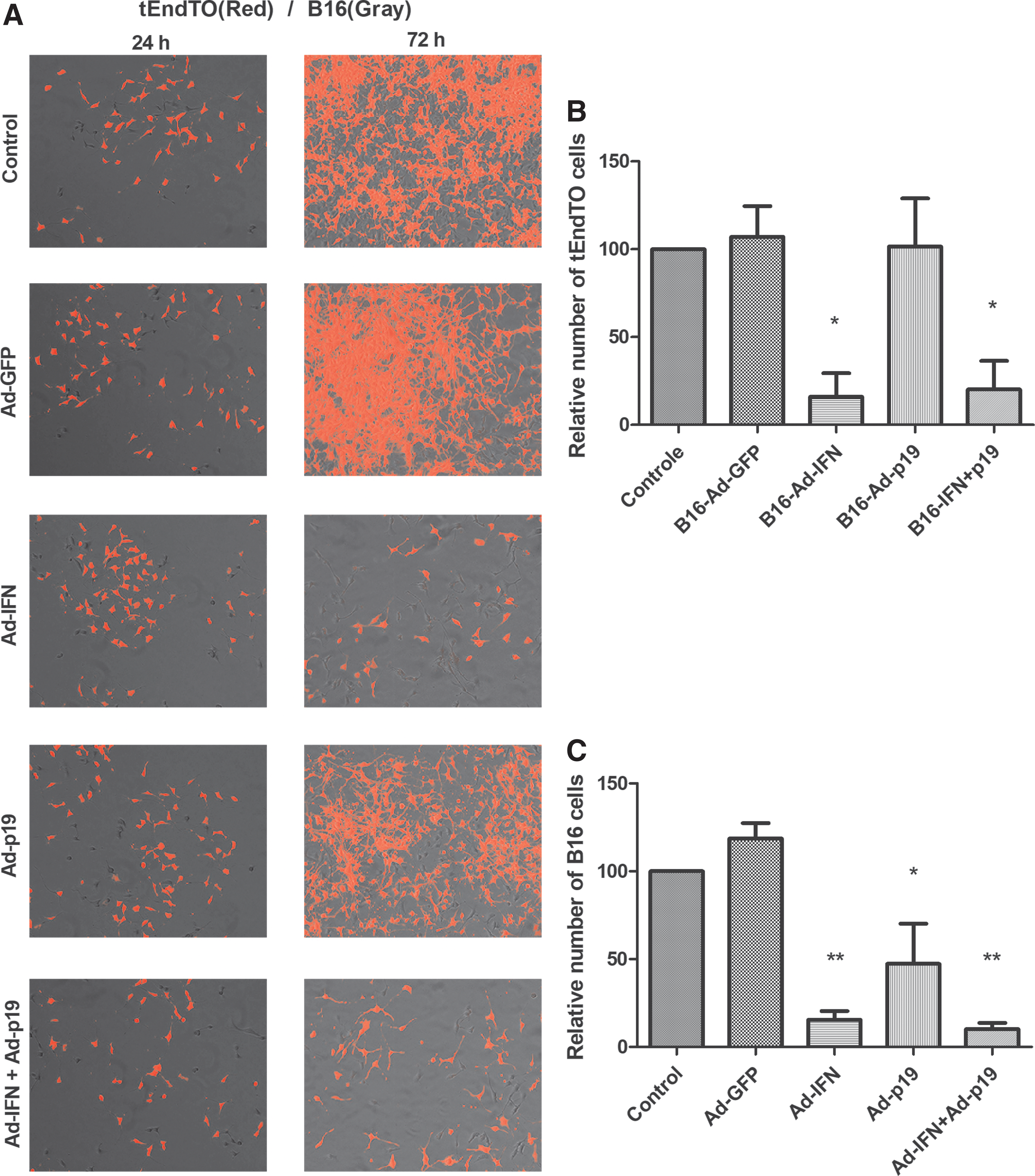
Coculture of tEndTO with previously transduced B16 cells. B16 cells were transduced (MOI 300), incubated for 6 h, washed, collected, and plated together with tEndTO cells at equal proportions. The coculture was incubated for up to 72 h.
Conditioned medium derived from B16 transduced with the Ad-IFN vector induces tEnd cell death
Next, we used the conditioned medium derived from B16 cells (cm-B16) transduced with the recombinant adenoviral vectors for the cultivation of tEnd cells and examined the impact of the tumor secretome on these endothelial cells. The clonogenic assays showed that tEnd cells treated with cm-B16-IFN or cm-B16-IFN+p19 significantly reduced the number of colonies formed (Fig. 4A, B). In the cell viability, cell cycle, and apoptosis analyses, tEnd cells were treated with conditioned medium for 48 h and then the tests were performed. The 3 tests showed that conditioned medium from cm-B16-IFN and cm-B16-IFN+p19 decreased viability, increased the population of hypodiploid, Annexin-V labeled cells, and cells with caspase 3/7 activity (Fig. 4C–F, respectively). Note that the use of Ad-p19 did not seem to influence these assays, suggesting a critical role for IFNβ. These data suggest that the conditioned medium containing the secretome from B16 transduced with the Ad-IFN vector induces tEnd cell death by a mechanism consistent with apoptosis.
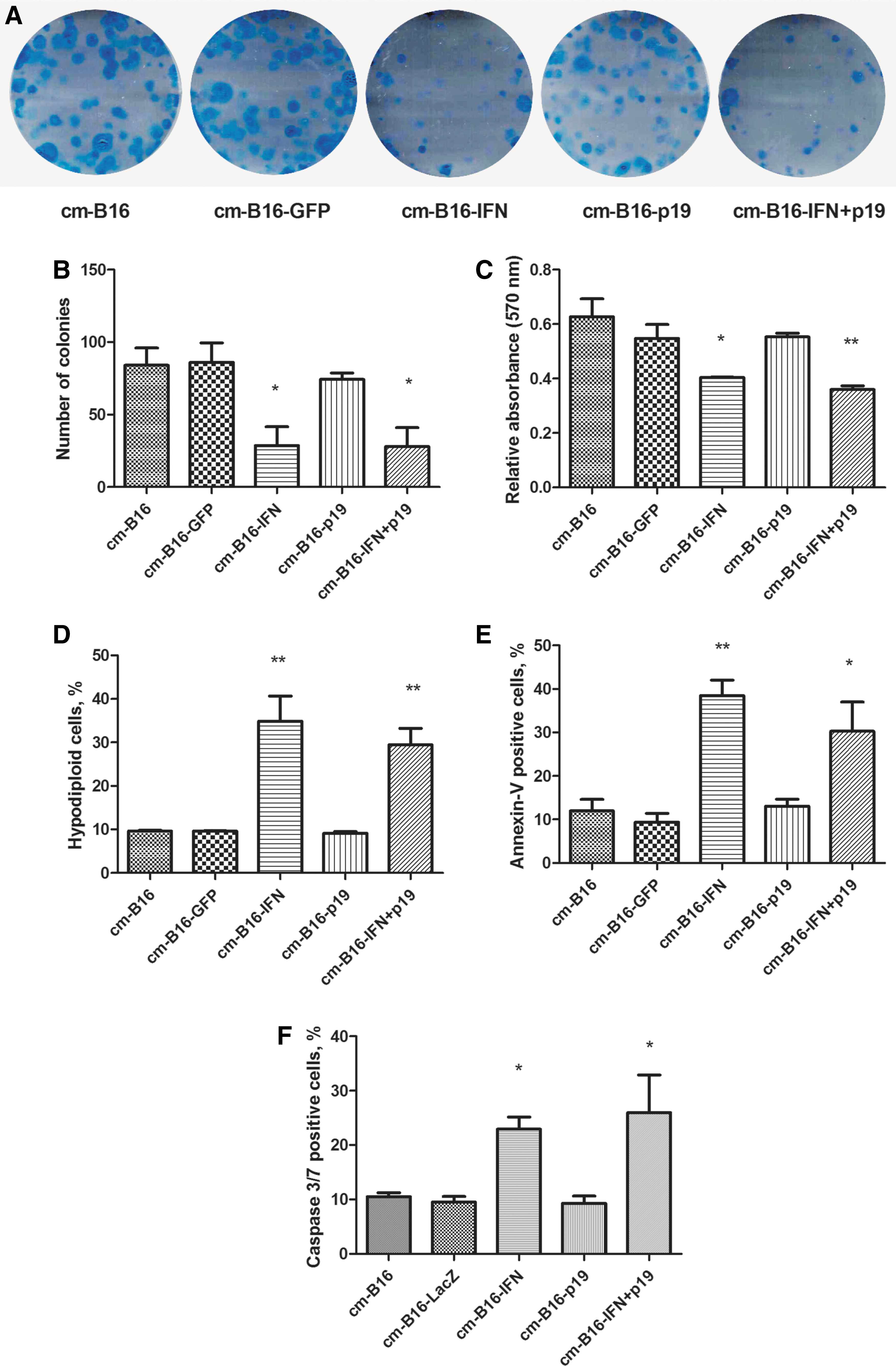
Conditioned medium derived from transduced B16 cells was used for the cultivation of tEnd cells. B16 was transduced (MOI 300) with the adenoviral vectors, incubated for 6 h, washed, and fresh medium was added. After incubation for 48 h, the conditioned medium was collected and stored at −80°C before use. The conditioned medium was named according to the transgene: cm-B16 (nontransduced), cm-B16-GFP, cm-B16-IFN, cm-B16-p19, and cm-B16-IFN+p19.
rIFNβ is not an inducer of tEnd cell death
Since the conditioned medium including IFNβ induced death of tEnd cells, we examined whether rIFNβ would be sufficient to bring about the same effect. We treated B16 and tEnd cells with rIFNβ 10 U/mL for 48 h before analyzing the cellular response. The percentage of hypodiploid, Annexin-V-labeled cells, and cells with activated caspases 3/7 was significantly increased only for B16, but not for tEnd, in the presence of rIFNβ (Fig. 5). This finding raises the intriguing possibility that one or more factors produced by B16 cells in addition to IFNβ are necessary for the killing of tEnd cells, as shown in Fig. 6.
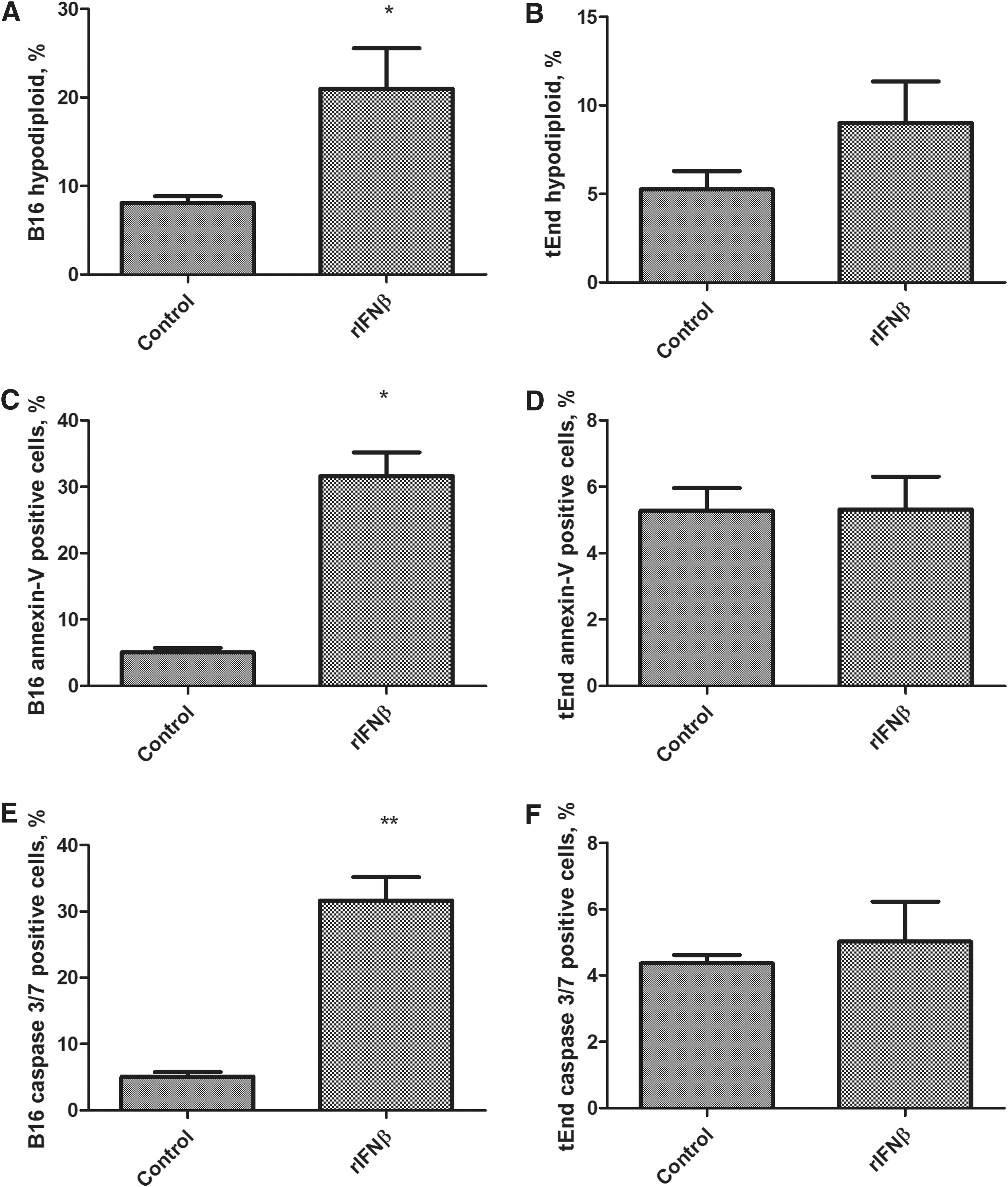
rIFNβ is a cell death inducer for B16 but not for tEnd cells. B16 and tEnd cells were incubated with rIFNβ 10 U/mL for 48 h. After treatment, cells were collected, fixed, and stained with PI
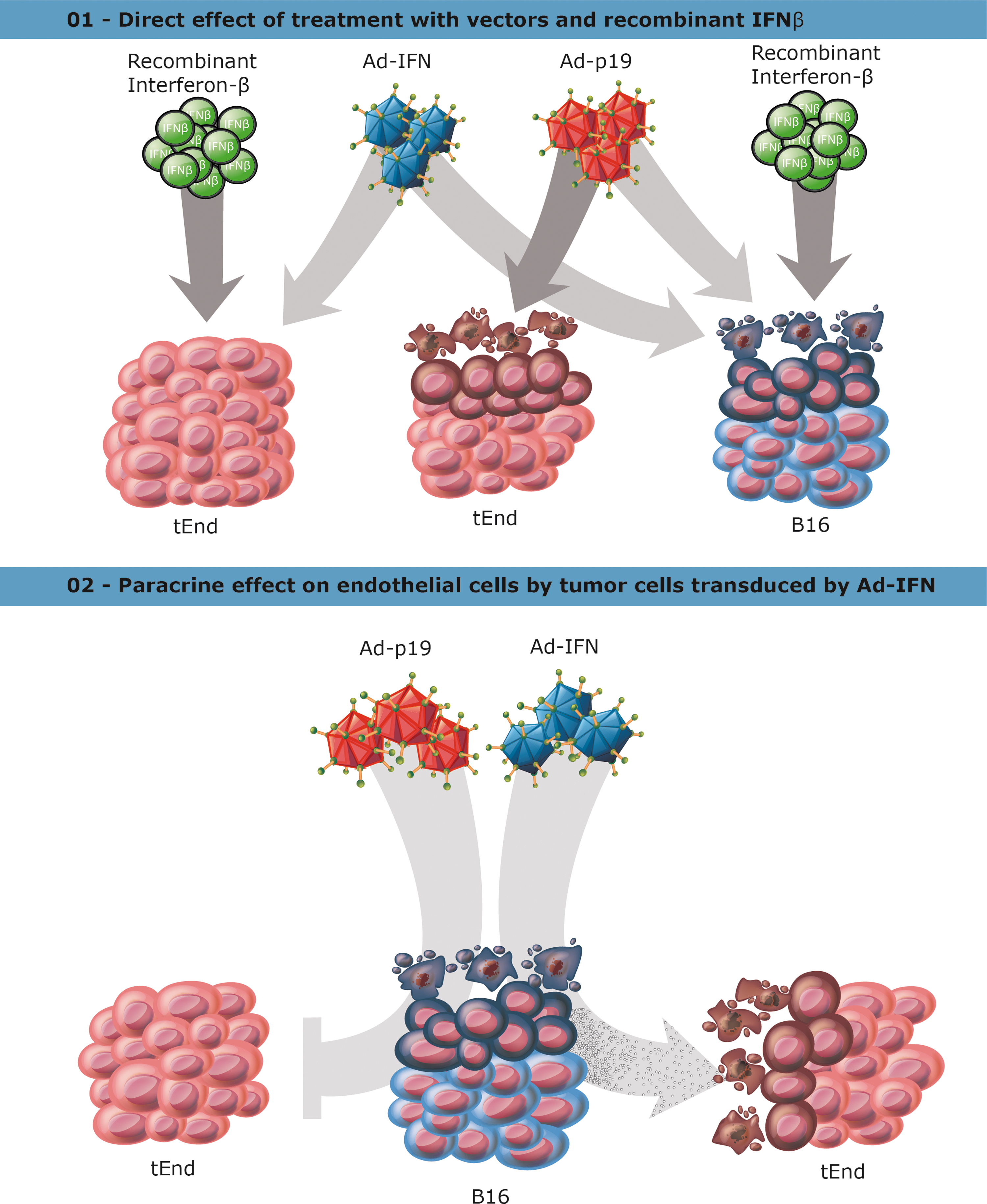
Schematic representation of cellular response to direct transduction or paracrine exposure to secreted factors.
Discussion
This study reveals the response of endothelial cells to our combined IFNβ+p19Arf gene transfer approach for the treatment of melanoma. In this study, we employed our AdRGD-PG vectors, containing the RGD-modified fiber and where transgene expression is controlled by p53, to transduce endothelial cells directly and observed that Ad-p19, but not Ad-IFN, was essential for impeding the angiogenic potential of tEnd cells and for the induction of their death. Alternatively, the secretome produced by B16 melanoma cells upon transduction with the adenoviral vectors also led to the death of tEnd cells; however, in this case B16 transduced with Ad-IFN was essential for inducing this response. Interestingly, rIFNβ was not sufficient to induce death in tEnd cells, but this response was not seen when treating B16 cells under the same conditions. We suspect that the secretome may contain important factors in addition to IFNβ, since direct transduction of tEnd with the vector encoding IFNβ had little impact. In either case, we postulate that our gene therapy approach would reduce tumor angiogenesis, by either direct or indirect signaling.
Adenoviral vectors have been widely used for cancer gene therapy due to their relative stability, efficient gene transfer in vivo, and broad tropism in terms of cell type (Crystal 2014; Kasala and others 2014). Even so, transduction by serotype 5 adenovirus (Ad5) relies on the expression of a specific cellular receptor [coxsackievirus and adenovirus receptor (CAR)], thus tropism may be considered limited if the target cell does not express CAR. Extensive efforts have been made to overcome CAR dependence and to direct adenoviral transduction to specific cell types (Khare and others 2011; Beatty and Curiel 2012; Alonso-Padilla and others 2015; Schmid and others 2018). A well-known approach for altering tropism involves the introduction of the RGD tripeptide in the adenovirus fiber protein, thus alleviating dependence on CAR and directing the virus to interact with cells expressing integrins (Mizuguchi and others 2001).
Since melanomas typically retain wild-type p53, we propose that this may serve as a molecular target of treatment. To this end, we have developed a series of viral vector platforms, including retrovirus, adenovirus, and adeno-associated virus, where transgene expression is under the control of p53 (Strauss and Costanzi-Strauss 2004; Bajgelman and Strauss 2008; Bajgelman and others 2013). Using the AdPG vector (native Ad5 fiber), we showed that the combination of IFNβ+p19Arf is beneficial for killing B16 cells in vitro and in vivo (Merkel and others 2013) and plays an essential role in NK cell activation and inducing a protective immune response (Medrano and others 2016). We then constructed the AdRGD-PG platform to improve viral tropism and have used these vectors to show the induction of immunogenic cell death in B16 (Hunger and others 2017) and for the stimulation of a protective immune response with critical participation of neutrophils in a mouse model of lung carcinoma (Catani and others 2016). These previous studies highlight the importance of the upregulation of IFNβ+p19Arf combination for the induction of cell death and immune activation.
As presented in this article, we have identified novel aspects of our gene transfer approach. In agreement with our previous work, the vectors were shown to express high levels of the transgene. Although not studied directly, we hypothesize that adenoviral vectors bearing the RGD motif in the adenoviral fiber protein may aid in the transduction of endothelial cells known to present high levels of integrins on the cell membrane (Danhier and others 2012). Certainly, the importance of integrins and the RGD motif for endothelial cell biology (Li and others 2008; Dettin and others 2015) and for the targeting of viral vectors (Mizuguchi and others 2001; Beatty and Curiel 2012; Pesonen and others 2012) has been extensively explored. Although the AdRGD-PG approach was not specifically developed for targeting tumor vasculature, this may indeed prove to be a benefit when using this vector platform.
Angiogenic characteristics of endothelial cells, such as migration and tube formation, were inhibited upon transduction of the tEnd cells with the Ad-p19. In addition, cell death by a mechanism consistent with apoptosis was also observed in this scenario. Since the expression of critical factors in the p53 pathway was concomitantly increased, we infer that these genes contributed to the cellular response since p21 participates in cell cycle control (el-Deiry and others 1993) and Puma induces apoptosis (Nakano and Vousden 2001; Chipuk and others 2005). Based on these results, we postulate that our gene therapy approach may indeed offer an antiangiogenic benefit when endothelial cells are transduced with a vector encoding p19Arf. Certainly, further exploration is necessary to confirm this point in vivo. It has been shown that activation of the p53/Arf pathway can significantly decrease cell motility by repression of phosphoinositide 3-kinase and Rac1 activity and can induce senescence (Guo and others 2003; Debidda and others 2006), additional activities that would favor an approach involving transduction of both the tumor and its microenvironment.
Using coculture or conditioned medium from transduced B16 cells, we observed the induction of tEnd cell death when IFNβ was included in the gene transfer step. Again, our melanoma gene therapy approach may indeed have the desired negative impact on angiogenesis since the secretome generated by the tumor cells treated with IFNβ was lethal for the endothelial cells. Intriguingly, rIFNβ was not sufficient to induce death of tEnd cells, suggesting that additional products secreted by B16 may be necessary for the killing of tEnd cells.
Our study raises several interesting possibilities. First, our melanoma gene therapy approach depends on the cooperation of IFNβ with p19Arf. However, we did not see evidence of cooperation that may yield an antiangiogenic effect, suggesting distinct mechanisms in B16 versus tEnd cells. Even so, the observations made in this study are potentially beneficial for tumor treatment. Second, we saw distinct activities when the endothelial cells were transduced as compared with their exposure to the factors released by the treated tumor cells. This too was not an expected finding, but still points to the antiangiogenic potential of our approach. Finally, we demonstrated that treatment with rIFNβ did not impact tEnd cells, although they were killed when exposed to factors released by transduced B16 cells. This is especially intriguing, suggesting that some factors other than IFNβ may have been responsible for this observation. Recall that transduced tEnd cells produced just as much IFNβ as B16 cells, yet the endothelial cells only responded to the tumor secretome generated after IFNβ gene transfer. Since IFNβ was present in both scenarios, we postulate that the latter involved additional factors, a point that will require extensive investigation in the future.
Based on examples from the literature, we would expect that IFNβ present in conditioned medium should have an impact on the exposed cultured cells. That is to say, we would expect a paracrine or bystander effect mediated by IFNβ. For example, conditioned medium from melanoma cells transfected with a plasmid encoding IFNβ showed a strong paracrine effect, where conditioned medium rich in soluble cytotoxic factors was cytotoxic for nontransfected melanoma cells (Rossi and others 2015). It has been shown that not only are the tumor cells targeted by a paracrine effect mediated by IFNβ, but also other cells in the tumor microenvironment are affected, undergoing apoptosis and reducing angiogenesis due to IFNα/β activity (Benedict and others 2004). Ex vivo gene transfer of IFNβ to bladder cancer cells delays growth of the tumor due to the induction of apoptosis of endothelial cells upon implantation in vivo (Izawa and others 2002), and IFNβ activates expression of genes related to decline in survival and proliferation (Gomez and Reich 2003). In another study, systemic IFNβ produced by liver-targeted adenoassociated virus (AAV) resulted in maturation of tumor vessels and improved blood flow, yet inhibited tumor growth by impeding further angiogenesis (Dickson and others 2007). In fact, our own work has shown a significant bystander effect where presence of p19Arf sensitized B16 cells to the effects of neighboring B16 cells that had been transduced with the IFNβ vector (Hunger and others 2017). Certainly, further testing, especially in vivo studies, will be required to reveal the full impact of our melanoma gene therapy approach on the tumor microenvironment in general and angiogenesis in particular.
With this study we have taken the first steps to show that our gene therapy approach combining upregulation of IFNβ and p19Arf may indeed impact endothelial cells. The use of an adenoviral vector containing the RGD tripeptide in the fiber protein yielded efficient transduction of the tEnd and B16 cell lines, direct transduction with the vector encoding p19Arf or exposure to the tumor secretome produced after transduction with the vector encoding IFNβ both resulted in endothelial cell killing. Although much remains to be elucidated, this study shows that our approach warrants further development and investigation.
Footnotes
Acknowledgments
We thank Dr. Roger Chammas (Instituto do Câncer do Estado de São Paulo, SP, Brazil) for continued support and helpful discussions and Dr. Cristina Maria Fernandes (Instituto Butantã, SP, Brazil) for providing the endothelial cell line, an essential component of this and future endeavors. Funding was provided by São Paulo Research Foundation, grant 13/25167-5, 15/26580-9 (BES) and fellowships 11/21256-8 (R.E.T.); 08/55963-0 and 11/10656-5 (A.H.). This study was financed, in part, by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES)—Finance Code 001 (I.L.V.).
Author Disclosure Statement
No competing financial interests exist.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.