
Abstract
Multifunctional pro-inflammatory cytokine CXCL8 is a small peptide of 8–10 kDa in size and it functions as a monomer or dimer. CXCL8 harbors 2 disulfide bonds for its stability. Although production of the CXCL8 protein in a large quantity in both mammalian and bacterial systems has been reported, the processes are complicated and lengthy. Here, we develop a new bacterial expression system for recombinant CXCL8 and simplify the purification system to yield a high amount of protein quickly. The purified CXCL8 protein from our new system develops a crystal structure that is identical to that produced through the mammalian expression system. Thus, we have established a simple and efficient recombinant CXCL8-producing system, which can be easily operated and is suitable to those requiring a large quantity of CXCL8.
Introduction
CXCL8,
CXCL8 plays important roles in various physiological and pathophysiological processes, including proliferation, survival, angiogenic responses, migration, and tumor metastasis. CXCL8 has also been used as a potential biomarker for clinical diagnosis, screening, follow-up, and prognosis of certain diseases (Liu and others 2015). The biological effects of CXCL8 are mediated by interacting with receptors CXCR1 and CXCR2, both of which are G-protein coupled receptors (Holmes and others 1991).
After forming a heterotrimeric complex with CXCR1/2, CXCL8 induces conformational change and activates various transcription factors and multiple signaling pathways, such as hypoxia inducible factor 1, nuclear factor kappa B, and activator protein 1 transcription factors, as well as PI3-K/Akt/mTOR, PLC/PKC, and MAPK signaling cascades (Hebert and Baker 1993; Waugh and Wilson 2008; Liu and others 2016).
Due to the many functions of CXCL8, CXCL8 has been extensively investigated and the recombinant CXCL8 protein is highly needed for research and possible applications. Although production of the recombinant CXCL8 protein in both mammalian and bacterial systems has been reported, it is difficult to obtain a high quality and a large quantity of CXCL8 protein due to insolubility and complicated purification procedures. Here, we demonstrate a simple and efficient recombinant CXCL8-producing system, from which the purified CXCL8 protein develops crystal structure identical to that produced through the mammalian expression system. Importantly, our recombinant CXCL8-producing system can be easily operated and is suitable to those requiring a large quantity of CXCL8.
Materials and Methods
Materials
Escherichia coli strain JM83 (#35607) was purchased from American Type Culture Collection (ATCC, VA, USA). GC5 competent cells (#42-650) were purchased from Genesee Scientific (CA, USA). Expression vector pASK75-strepII was kindly provided by Dr. Arne Skerra (Technische Universität München, Freising-Weihenstephan, Germany) (Skerra 1994). Plasmid extract kit was purchased from Qiagen (MD, USA). Primers were synthesized by Integrated DNA Technologies (IA, USA). Phusion High-Fidelity DNA Polymerase was purchased from New England Biolabs (MA, USA). LB agar, LB broth, ampicillin, hen egg-white lysozyme, and anhydrotetracycline were purchased from Fisher Scientific (IL, USA).
Ni Sepharose columns (HisTrap™ HP, 5 mL) and HiLoad™ 26/600 Superdex™ 200 pg columns were purchased from GE Healthcare (NJ, USA). Amicon Ultra centrifugal filter devices −10,000 nominal molecular weight limit (10 kDa) were purchased from EMD Millipore (MA, USA). The protein marker PageRuler™ Prestained Protein Ladder (#26616) was purchased from Thermo Fisher Scientific (MA, USA). The equipment for gel electrophoresis comprised products of Bio-Rad (CA, USA). SaltRx HT (#HR2-136), Crystal Screen HT (#HR2-130), and Index HT were purchased from Hampton Research (CA, USA). All reagents used were of reagent grade.
Cloning
Human CXCL8 cDNA (NCBI:NP_000575) was amplified by PCR from plasmid containing full-length human CXCL8 (Origene, MD, USA) and cloned into pASK75, with a C-terminal His6-tag and an N-terminal OmpA signal sequence for periplasmic secretion in E. coli (Schiefner and others 2015). The cDNA was integrated into the expression vector pASK75 by the ligation-independent PCR cloning method (Li and others 2011). Forward and reverse primers used were pASK75_F: 5′-CATCACCATCATCACCATTAATAAGC-3′, pASK75_R: 5′-AGCCTGCGCTACGGTAGCGAAACC-3′, CXCL8_F28: 5′-TCGCTACCGTAGCGCAGGCTAGTGCTAAAGAACTTAGATGT-3′, and CXCL8_R99: 5′-TGGTGATGATGGTGATGTGAATTCTCAGCCCTCTTCAAAAAC-3′, respectively. The nucleotide sequence of the resulting construct was confirmed by sequencing. The recombinant plasmids were amplified in GC5 competent cells, and the plasmid was subsequently transformed into E. coli JM83.
Protein expression and purification
Human CXCL8-His6 was produced in the periplasm of E. coli JM83. For expression, cells were incubated in an LB medium supplemented with 100 μg/mL ampicillin at 37°C and 250 rpm for overnight. Fifteen milliliters of the overnight culture was used to inoculate 900 mL of the LB medium and incubated at 220 rpm and 22°C under agitation. At an optical density of OD550 of 2.0, expression of CXCL8 was induced by 0.2 mg/L anhydrotetracycline for 4 h. Cells were harvested by centrifugation at 7,000 rpm for 15 min, and the supernatant was discarded. The pellet from 900 mL cell culture was resuspended in periplasmic fraction buffer (0.5 M sucrose, 1 mM EDTA, 0.1 M Tris/HCl pH 8.0) supplemented with 0.2 mg/mL hen egg-white lysozyme.
After incubation on ice for 30 min, spheroplasts were removed by centrifugation. The resulting periplasmic extract was dialyzed against immobilized metal ion affinity chromatography (IMAC) buffer (1 M NaCl, 50 mM NaPi pH 7.5) (Schiefner and others 2015). The recombinant protein was purified by IMAC on Ni Sepharose columns charged with Ni2+ ions by using a linear imidazole concentration gradient. Appropriate fractions were pooled and finally purified to homogeneity by high-resolution size-exclusion chromatography (SEC) on a HiLoad™ 26/600 Superdex™ 200 pg columns equilibrated with 50 mM NaCl, 10 mM Hepes (pH 7.4). The fractions were combined and concentrated by 10 kDa molecular weight cut-off Amicon Ultra centrifugal filter units.
Protein crystallization
For protein crystallization, automatic dispenser Crystal Gryphon from Art Robbins Instruments (CA, USA) was used for high-throughput crystallization of CXCL8, and commercially available crystallization reagent systems were used for an initial screening: SaltRx HT, Crystal Screen HT, and Index HT.
X-ray diffraction data collection and protein structure analysis
The crystals were harvested by using nylon loops after removing excess mother liquor. Harvested crystals were flash-frozen and stored in liquid nitrogen. An x-ray diffraction data set with Bragg spacings to 1.25 Å was collected on a Pilatus3 6M detector at the 5.0.2 beamline at the ALS, LBNL. Data were processed with iMOSFLM software (Battye and others 2011). The structure of the human CXCL8 was solved by the molecular replacement method by using BALBES with 5D14 as the search model (Long and others 2008). Crystallographic refinement was performed by using a combination of PHENIX 1.12 (Adams and others 2010) and BUSTER 2.9 (Bricogne and others 2010).
Manual rebuilding and adjustment of the structure were carried out by using the graphics program Coot (Emsley and Cowtan 2004). Data processing and refinement statistics are shown in Supplementary Table S1. Molecular figures were created by using PyMOL software (Schrödinger). The crystal structure was deposited in PDB under ID code 6N2U.
Cell proliferation assay
Human prostate carcinoma LNCaP cells were cultured in RPMI 1640 medium supplemented with 1% antibiotic-antimycotic and 10% fetal bovine serum (FBS), and they were incubated at 37°C in a humidified atmosphere containing 5% CO2. The cells were seeded in 96-well plates at a density of 1 × 104 cells per well. After culturing in charcoal stripped FBS containing media with CXCL8 (1 μg/mL) or control (phosphate-buffered saline) for 48 h, 10 μL of MTT dye was added and incubated for 4 h at 37°C. Then, the original media was removed, and 100 μL of dimethylsulfoxide was added to each well and gently shaken for 10 min on a rocker. The plate was measured at 570 nm with a reference wavelength of 630 nm by using a 96-well plate reader (Tecan, USA).
Results
Cloning and expression of CXCL8
The human CXCL8 sequence encoding the 72 amino acid (SAKELRCQCIKTYSKPFHPKFIKELRVIESGPHCANTEIIVKLSDGRELCLDPKENWVQRVVEKFLKRAENS) with a His-tag at C-terminal was inserted into pASK75 vector behind a N-terminal OmpA signal sequence (Skerra 1994). The established plasmid, named pASK75-CXCL8-His, was transformed into E. coli JM83, and the expression of CXCL8 protein was induced by anhydrotetracycline. The protein was subjected to sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE), and the expression of CXCL8 protein was confirmed by Coomassie staining.
Purification of CXCL8 by fast protein liquid chromatography
To isolate CXCL8 protein in periplasm of E. coli JM83, the pellet from large-scale culture was resuspended and lysed in periplasmic fraction buffer. After incubation, the periplasmic lysate was centrifuged and the supernatant was collected. The resulting periplasmic extract was dialyzed against IMAC buffer as described in the Materials and Methods section. Since the recombinant protein has a His-tag at C-terminal, His-tagged affinity chromatography using Sepharose columns was chosen to capture the protein. The dialyzed periplasmic extract was loaded onto the HisTrap™ HP 5 mL, and the CXCL8-containing fraction was eluted and collected in 96-well collection plates. Two peaks were detected from the CXCL8-containing fraction in the chromatogram (Fig. 1), and the fractions were analyzed by SDS-PAGE (Fig. 2).
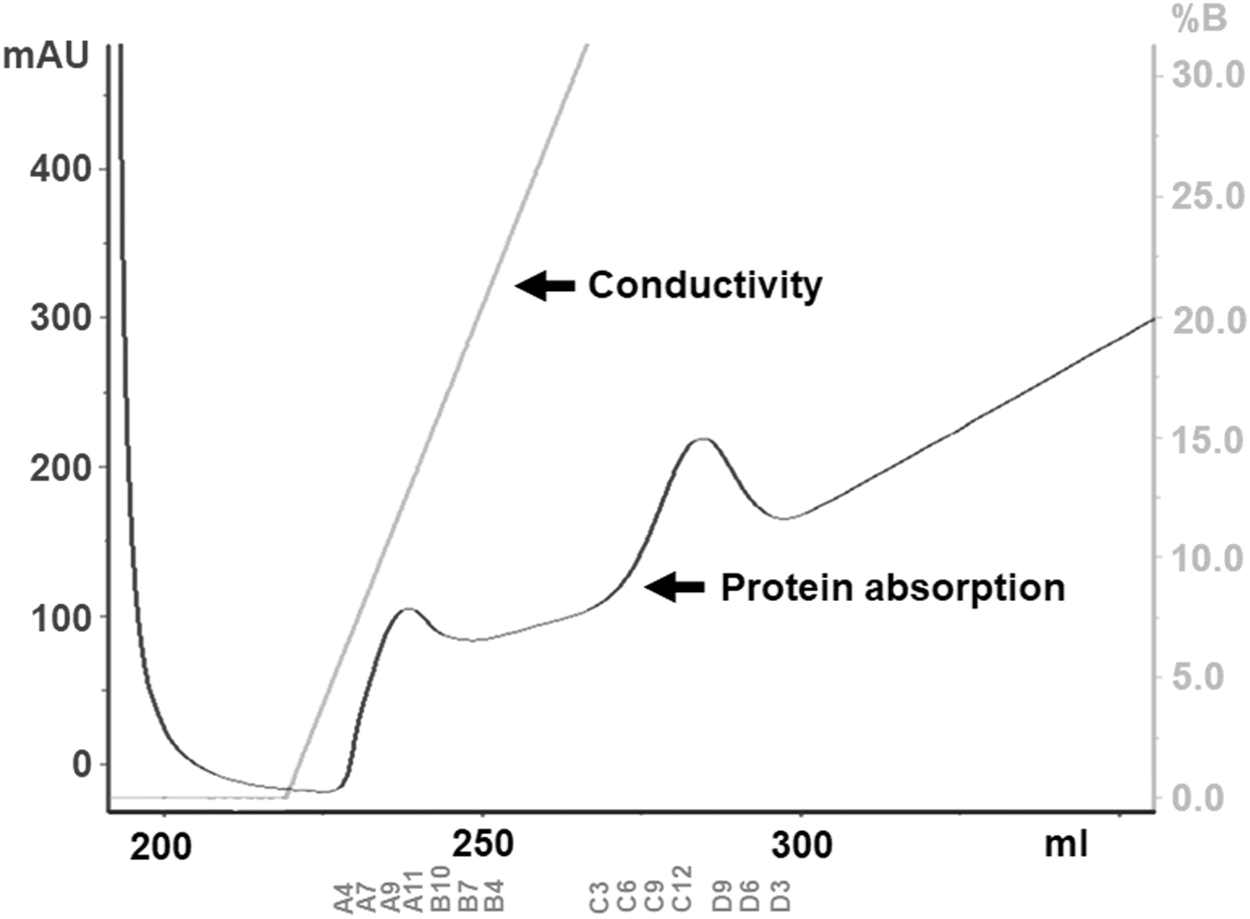
Chromatogram of CXCL8 protein purification by FPLC. Protein absorption at 280 nm is shown as a dark gray line, and conductivity is shown as a light gray line. CXCL8 peaks are shown on immobilized metal ion affinity chromatography (IMAC) by using Ni Sepharose columns (HisTrap™ HP, 5 mL). FPLC, fast protein liquid chromatography.
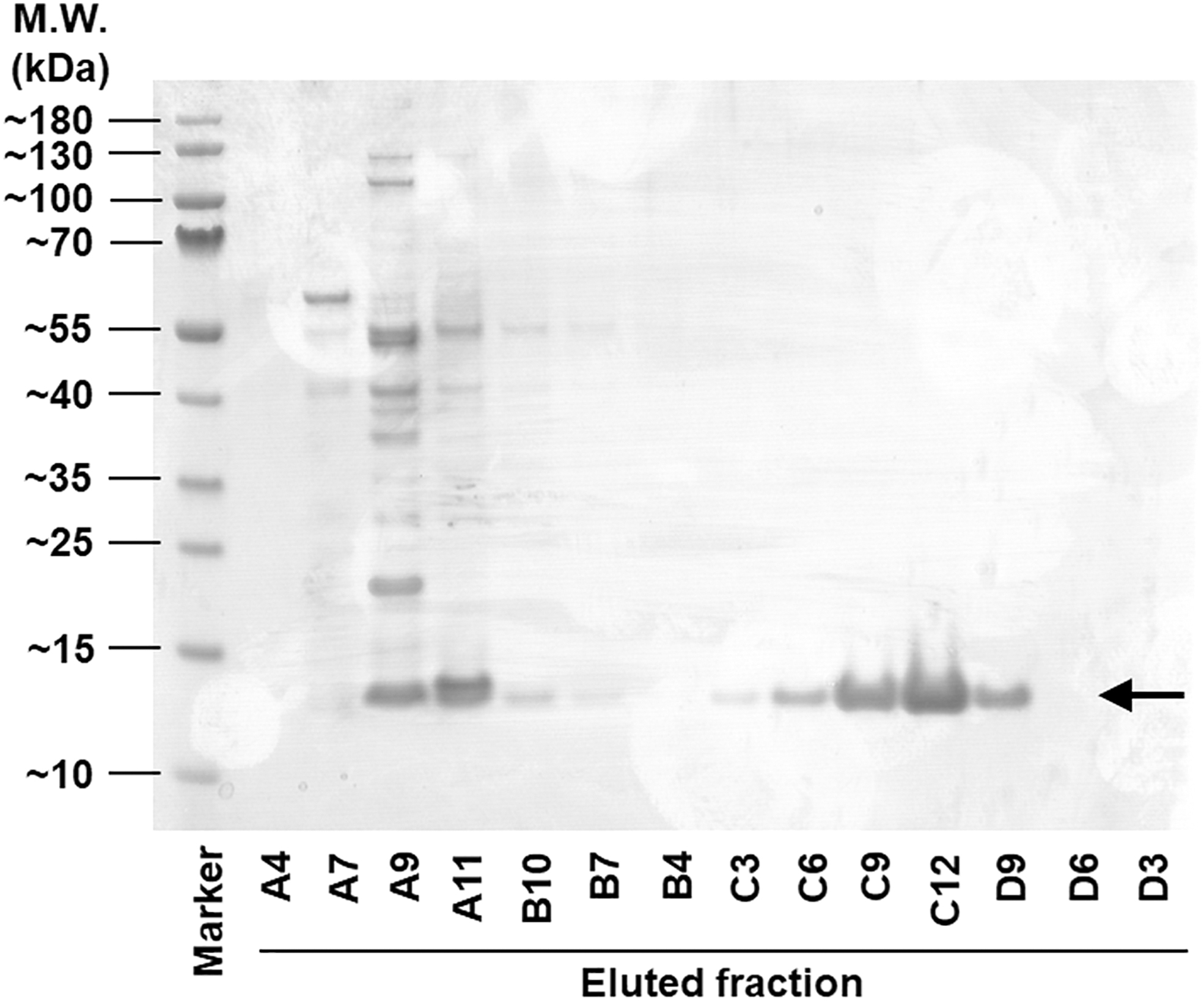
CXCL8 protein in different chromatography fractions. Chromatography fractions were loaded onto a gradient SDS-PA gel, and they were stained with Coomassie blue R250. Alphabetical sequence numbers indicate eluted fractions number from a 96-well collection plate. Arrow indicates the monomer of CXCL8 (∼12.5 kDa).
Although the theoretical monomer mass of CXCL8 is 8–10 kDa, the protein bands of CXCL8 monomer were observed at ∼12.5 kDa in the SDS-PAGE. This phenomenon is frequently observed in the production of recombinant chemokine proteins due to its elevated isoelectric point (pI) (Guan and others 2015; Gao and others 2019). In addition, a His-tag at its C-terminal also contributed to the increased molecular weight. However, although the OmpA signal sequence was likely removed during secretion, and may have yielded a polypeptide that starts with the mature CXCL8N-terminus, this was not confirmed by MS or EC50 determination and thus the sequence identity of the purified protein remains known only approximately. The CXCL8 protein was visualized by Coomassie staining from A9 to A11, and from C3 to D9 in sequential elution order as indicated by an arrow in Fig. 2.
Since the fractions from A9 to A11 expressed proteins with various sizes, indicating the expression of nonspecific proteins, CXCL8 containing fractions only from C3 to D9 were collected to remove contaminants. The collected fraction was separated according to the differences in size and structure by SEC to increase the protein homogeneity. For crystallization, CXCL8-containing fractions from SEC were combined and concentrated to ∼3.4 mg/mL with 10 kDa molecular weight cut-off Amicon Ultra centrifugal filter unit. The final yield of CXCL8-His6 was 1.2 mg/L E. coli culture.
Crystallization of CXCL8 protein
CXCL8 crystals were screened by an automatic dispenser with crystallization reagent systems as described in the Materials and Methods section. Several initial hits were observed in SaltRx HT, and the crystallization condition was systematically optimized to obtain high-quality crystals. For secondary screening, individual chemicals were added by serially changing pH value or buffer concentration by the hanging drop method. The best-hit condition for obtaining diffraction quality crystals was in the presence of 1.8 M sodium phosphate monobasic monohydrate, potassium phosphate dibasic, pH 8.2. The crystals were fully grown within 7 days, and its morphology exhibited a rhombic prism (Fig. 3A).
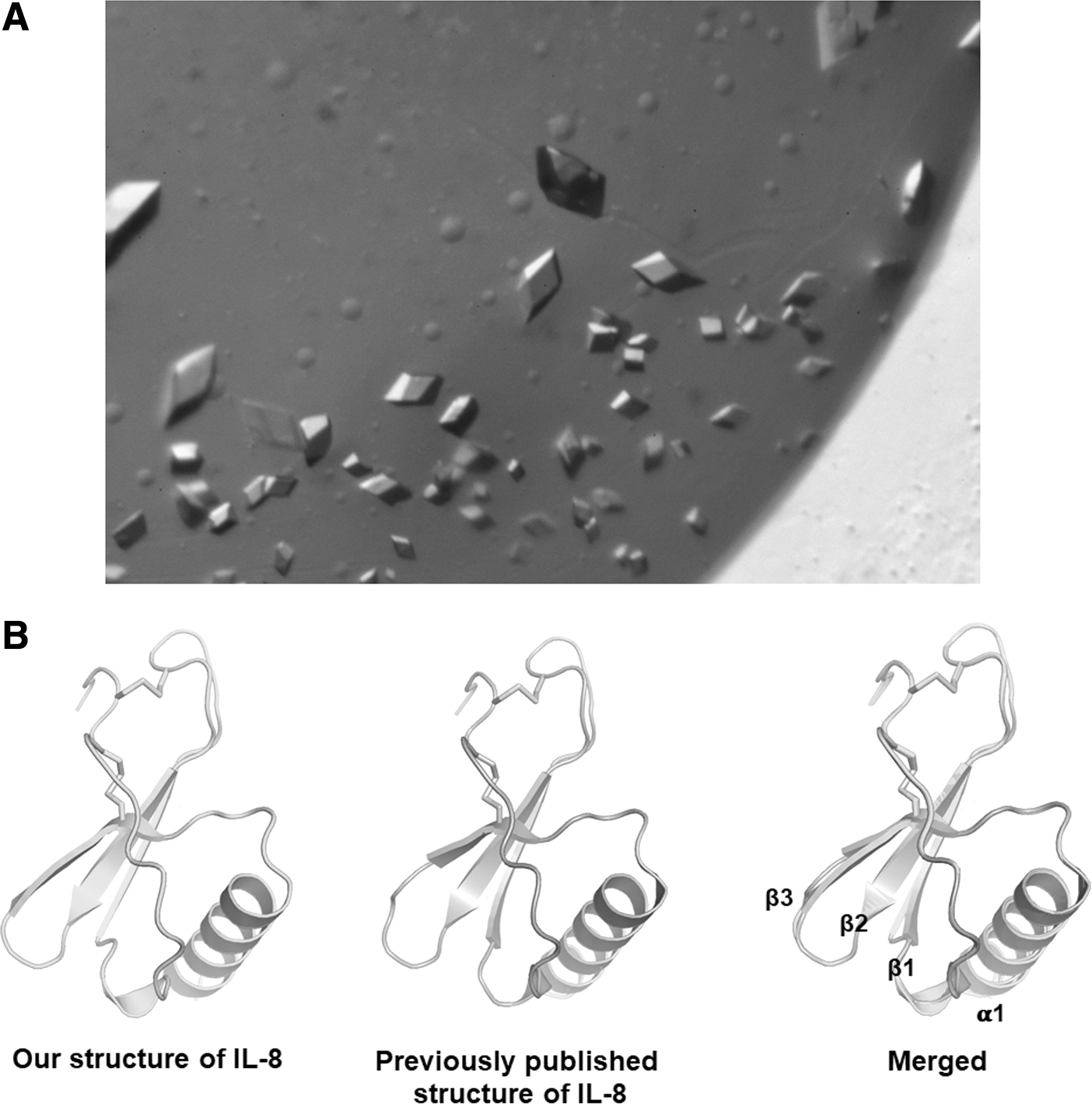
Crystallization of human CXCL8.
Three-dimensional structure of CXCL8
After selecting the best crystals, X-ray diffraction data were collected at the Advanced Light Source. The structure of the CXCL8 was determined by the molecular replacement method followed by crystallographic refinement of the atomic models. The 3-dimensional structure of CXCL8 contains the NH2-terminal loop, 3 anti-parallel β-strands connected by loops, and the 1 C-terminal α-helix (Fig. 3B). Turquoise is our structure of CXCL8, and the gray is a previously published structure (Fig. 3B). To compare similarity between the 2 structures, the alignment was performed and the root-mean-square deviation (RMSD) was calculated. The RMSD between the 2 structures was 0.196 over 64 Cα positions, indicating that the structures are nearly identical.
The recombinant CXCL8 promotes the proliferation of cancer cells
Since it has been reported that the CXCL8 promotes prostate cancer growth (Singh and Lokeshwar 2011), we examined the biological activity of our recombinant CXCL8 protein on the cell proliferation of prostate cancer cells. We found that our recombinant CXCL8 significantly increased the LNCaP cell proliferation (Fig. 4), suggesting that the recombinant CXCL8 protein produced in our system functions properly in vitro and has biological activity.
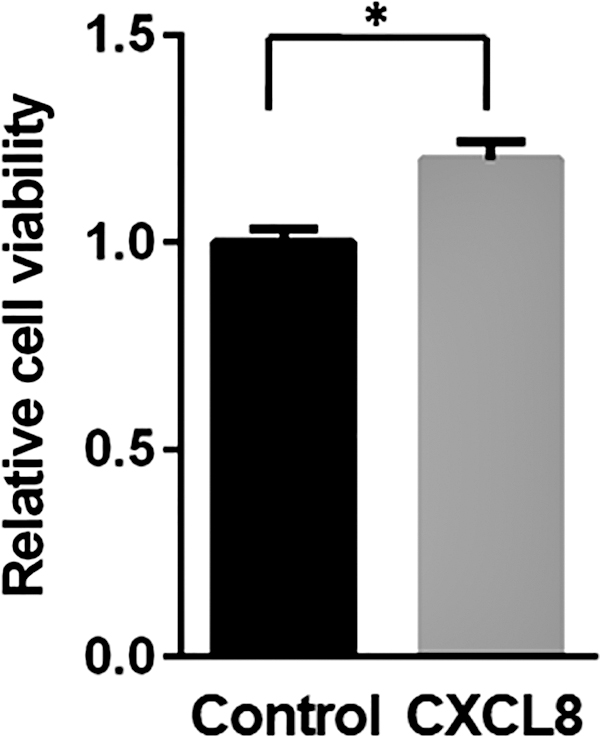
The effect of CXCL8 on the cell proliferation of LNCaP cells. LNCaP cells were treated with the CXCL8 (1 μg/mL) or control (phosphate-buffered saline) for 48 h. The cell proliferation was determined by using the MTT assay. Values represent the mean ± SD of triplicate experiments. *P < 0.01, statistically significant compared with the control. SD, standard deviation.
Discussion
CXCL8, a member of the CXC family, is a chemotactic factor secreted by macrophages, T cells, monocytes, fibroblasts, and epithelial cells (Hebert and Baker 1993; Xie 2001; Waugh and Wilson 2008; Liu and others 2016). As CXCL8 plays important roles in pro-inflammation responses, CXCL8 is considered one of the major potential therapeutic targets in autoimmune, inflammatory, and infectious diseases (Friedland 1996; Russo and others 2014; Lee and others 2017). The interaction of CXCL8 with its receptors is of considerable interest for drug discovery and immunology research.
To study the physiological function of CXCL8, large quantities of CXCL8 are required for in vitro or in vivo assays. To date, many efforts have been made to optimize the protocol and to improve the efficiency for the production of CXCL8; the strategies that efficiently improve the solubility of recombinant CXCL8 protein and simplify the purification procedures of CXCL8 are still lacking (Supplementary Table S2). For instance, Furuta and others partially purified CXCL8 as a water-soluble polypeptide by re-naturation from inclusion bodies of both the soluble and insoluble protein fractions to increase total yield amount (Furuta and others 1989). Wiese and Schmitz (2011) generated pelB-IL8 fusion protein by using pelB-leader peptide, cleaved off its N-terminus by an endogenous bacterial peptidase.
This provided wild-type CXCL8 with a serine at its N-terminus in the soluble fraction after lysis from the pellet. Cheng and others (2008) mutated CXCL8 at K11 and G31 positions, and synthesized as an analogue of CXCL8 in the E. coli expression system. This analogue had high affinity for both CXCR1 and CXCR2, which were able to block the functions of CXCL8 to activate or chemoattract neutrophils in vitro. Li and others (2009) employed the yeast expression system instead of the E. coli expression system, and they purified active CXCL8 from the fermentation supernatant by using Pichia pastoris with the AOX1 promoter. In all these reported CXCL8-producing systems, a number of different buffers and time-consuming dialysis steps are needed to obtain soluble and pure CXCL8 protein.
In this study, we show that CXCL8 protein can be successfully expressed and purified in the E. coli expression system. Soluble CXCL8 is secreted by the N-terminal OmpA signal sequence of pASK75 vector, and it is captured in the periplasm of E. coli JM83 without the formation of inclusion bodies. The CXCL8 protein in the periplasm is easily purified by 1 lysis step using periplasmic fraction buffer. Since this lysis step does not include an additional re-naturation step or precipitation step from the supernatant, our system is more efficient than previous systems. CXCL8 from our system is simply cloned and expressed without any fusion tag protein, including SUMO, MBP, and GST, or any mutation to synthesize an analogue of CXCL8, which provides a time-saving way by relieving additional cloning works.
In addition, high-purity CXCL8 protein (>95%) is required for in vitro or in vivo assays (Hou and others 2005), and high-quality CXCL8 protein is required for crystallography applications (Kim and others 2008). CXCL8 obtained from our system has biological activity and is successfully applied for crystallization, which indicates that the CXCL8 protein is soluble, folded, stable, and is chemically, conformationally, and functionally homogeneous, and free of critical contaminants. In conclusion, we have established a simple and efficient recombinant CXCL8-producing system, which can be easily operated and is suitable to those requiring a large quantity of CXCL8.
Footnotes
Acknowledgments
The authors thank Dr. Arne Skerra in Technische Universität München, Freising-Weihenstephan, Germany, who kindly provided them with the expression vector pASK75-strepII.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2020R1I1A1A01052072), and a scholarship from the Frenchman's Creek Women for Cancer Research to J.-H.J.
Supplementary Material
Supplementary Table S1
Supplementary Table S2