
Abstract
Epidural fibrosis after lumbar laminectomy refers to a serious complication, and excessive proliferation of fibroblasts is considered the major factor. Interferon-alpha-2b (IFN-α-2b) can exert antiviral and antiproliferative effects, which has been suggested to effectively prevent several fibrotic diseases. However, the effect of IFN-α-2b on the prevention of epidural fibrosis (EF) and its possible mechanism remain unclear. In this study, in vitro and in vivo experiments were performed to examine the possible mechanism of IFN-α-2b for preventing EF. Cell counting kit-8 (CCK-8), cell cycle test, Edu incorporation, wound healing assay, transwell test, and Western blotting assay were performed to investigate the inhibitory effect of IFN-α-2b on the proliferation and migration of fibroblasts in vitro. As indicated from the results, IFN-α-2b was capable of inhibiting proliferation and migration of fibroblasts and inhibiting the activity of the transforming growth factor β (TGFβ)/Smad signaling pathway. In vivo, the effect of IFN-α-2b on the reduction of EF was determined by performing histological macroscopic evaluation and histological and immunohistochemical staining. As suggested from the results, IFN-α-2b significantly inhibited EF after laminectomy. As revealed from the mentioned results, IFN-α-2b may have a promising application for preventing EF in the future.
Introduction
Lumbar laminectomy is capable of effectively treating lumbar spondylolisthesis and lumbar spinal stenosis, whereas excessive epidural fibrosis (EF) after a lumbar surgery is recognized as a troublesome complication, thereby even causing various clinical symptoms and even reoperations after a spinal surgery once (Lee and others 2015). The back surgery failure syndrome (FBSS) refers to a type of serious complication, primarily manifested as persistent back or nerve root pain after surgeries (Lee and others 2014; Sun and others 2016). As indicated from the reports, ∼8%–40% of patients unfortunately have the FBSS, and 4%–9% require a second surgery (Guyer and others 2006). For this reason, surgeons should operate as gently as possible.
As reported in existing studies, extradural fibrosis, primarily attributed to an excessive proliferation of fibroblasts, can adhere to the dura mater or be extended to the nerve root canal, thereby generally causing leg and low -back pain (Tian and others 2015; Waszak and others 2016).
The authors have previously studied a large number of drugs for preventing EF (e.g., triptolide, artesunate, and tacrolimus) (Li and others 2017; Dai and others 2019; Wan and others 2019). However, as impacted by the side effects or the route of administration of the mentioned drugs, considerable limitations remain before their clinical trials. Accordingly, a reasonable, safe, and effective drug with less side effect is required to prevent EF.
Interferon (IFN) acts as a vital cytokine exhibiting antitumor, antiproliferative, and immunomodulatory activities. Currently, based on their nucleotide sequences, interactions with specific receptors, structures, and chromosomal locations, 3 types of IFNs were distinguished. Type I IFNs consist of α, β, ω, κ, ɛ, ζ, τ, δ, and ν. IFN-γ refers to the representative of type II IFN. The most recently classified type III IFNs comprise IFN-λ1, IFN-λ2, and IFN-λ3 (Asmana Ningrum 2014; Bandurska and others 2014). In clinical treatments, the subtype IFN-α2 [i.e., IFN-α-2a and interferon-alpha-2b (IFN-α-2b)] has been most extensively used since 1986 when it was initially approved as a therapeutic protein (López de Padilla and Niewold 2016).
However, compared with IFN-α-2b, patients administrated with IFN-α-2a will synthesize anti-IFN-α antibodies at a higher rate, thereby reducing the therapeutic effect (Butler and others 2012). As reported in existing studies, the use of IFN-α-2b is capable of treating some fibroproliferative diseases (e.g., hypertrophic scars and keloids). Moreover, it is involved in the treatment of fibroproliferative diseases (Berman and Flores 1997; Ghahary and others 2001), which has some advantages over IFN γ for treating some fibroproliferative diseases (e.g., hypertrophic scars) (Ghahary and others 1995). Thus, IFN-α-2b was selected for experimental exploration and research.
The cytokine transforming growth factor β1 (TGF-β1) has been extensively reported to be critical to numerous fibrotic diseases. In vitro, TGF-β1 can promote the proliferation of fibroblasts and produce more extracellular matrix (ECM) proteins (Park and others 2018; Wei and others 2018; Lin and others 2020). TGF-β1 first binds to TGF-β receptor II (TGFβRII) and then to TGFβRI to form catalytically active TGFβR. Subsequently, active TGFβR recruits and phosphorylates Smad2 and Smad3 in the cytoplasm. The TGFβ receptors lay the basis for TGF-β1 signal transduction and its biological effects, and the downstream drosophila mothers against decapentaplegic protein (Smad) are capable of directly transmitting the signal of the cell surface receptor to the nucleus (Massagué 2012). In the Smad proteins, Smads 2 and 3 critically impact the TGFβ/Smad signaling pathway (Derynck and Budi 2019).
In this study, whether IFN-α-2b can reduce EF formation and whether it can inhibit the proliferation and migration of fibroblasts by regulating the TGFβ/Smad pathway were investigated, which might supply a novel treatment option for preventing postoperative EF in the future.
Materials and Methods
Reagent
Recombinant human IFN-α-2b, molecular 19.2 kDa, purity >95%, originated from GenScript Biotechnology Co., Ltd. (Nanjing, China).
Fibroblast culture and IFN-α-2b treatments
The human fibroblast cell line was offered by Jenino Biotech Co., Ltd (Guangzhou, China) and then cultured in a medium supplemented with Dulbecco's modified Eagle's medium Beyotime (Gibco) based on 15% fetal bovine serum (FBS; Gemini), 1% penicillin/streptomycin (Beyotime, Shanghai, China), at 37°C with 5% CO2. Fibroblasts between passages 3–5 were selected to perform the subsequent experiments. Specific to the experiments in vitro, fibroblasts were cultured, with the density reaching 60%–70%, after being washed with phosphate buffered saline (PBS) and administrated with IFN-α-2b at different concentrations.
The cell viability assay
The cells were administrated with IFN-α-2b at different concentrations in different periods, and the effect of IFN-α-2b on cell viability was examined by the cell counting kit-8 (CCK-8; Dojindo, Tokyo, Japan). In brief, 100 μL cell suspension was planted in 96-well plates under the density of ∼5 × 103 cells/well. Under the density of cells reaching ∼60%–70%, IFN-α-2b at different concentrations was treated for 24, 48, and 72 h. Subsequently, 10 μL CCK-8 solution was added to the respective test well. The plate was incubated for another 2 h at 37°C. The absorbance at 450 nm was determined with a microplate absorbance reader (Tecan, M1000, Switzerland). The cell viability was calculated by the formula in the reagent manual.
Cell cycle analysis
The Cell Cycle Testing Kit (Beyotime, Shanghai, China) was adopted to analyze the cell cycle. In brief, the fibroblasts were harvested after 24 and 48 h in the control and the 5,000 U/mL IFN-α-2b treatment group and then fixed in 70% ice-cold ethanol at 4°C overnight. Next, the cells were collected through centrifugation, resuspended in 0.5 mL propidium iodide, and then incubated for 30 min at ambient temperature in the dark. Afterward, the flow cytometry test was performed, and ModFit LT software was finally used to calculate the distribution of cells in different periods of G0/G1, S, and G2/M.
EDU incorporation assay
The Cell-Light KFluor555 Edu Kit (KeyGEN, Nanjing, China) was employed to assess the fibroblast proliferation. In brief, the fibroblasts were planted in 6-well plates at a density of 1 × 105 cells per well. The cells were cultured at 37°C for 24 h, treated with IFN-α-2b at different concentrations for 48 h, and then incubated in the medium supplemented with 10 μM 5-ethynyl-2′-deoxyuridine (EdU) for 2 h. Next, they were fixed in 4% paraformaldehyde for 15 min and then infiltrated with 0.5% Triton x-100 for 20 min. Furthermore, the mentioned cells were stained with Hoechst 33342, incubated at ambient temperature in the dark for 30 min. Lastly, the positive staining was observed under an inverted fluorescence microscope.
Cell migration assay
The wound healing assay and the transwell migration test were performed to assess the cell migration behavior. In brief, the culture inserts in each well were transferred to a 12-well plate, each compartment of the insert was filled with 70 μL of the cell suspension, and the plate was incubated at 37°C for 24 h. It was ensured that a monolayer of fused cells was formed first, then the inserts were gently removed. The rinsing process was conducted to remove dead cells, and then the serum-free medium and test reagents were added, and f Zeiss inverted microscope was finally employed to capture the images at the designed times (0, 12, 24, and 48 h). ImageJ software was adopted to process the images and calculate the wound healing rate.
The fibroblasts (5 × 104 cells) were transferred to the transwell inserts, and a range of media were used in 24-well plates. After 24 h, the transwell insert was washed gently with PBS, fixed with 4% paraformaldehyde, and then stained with crystal violet. The cells not penetrating the transmembrane were gently wiped with a cotton swab. Under the Zeiss inverted microscope, the migrated cells were observed. The number of the migrated cells was counted and processed with ImageJ software.
Western blotting analysis
The fibroblasts were collected after the treatment with IFN-α-2b at different concentrations. The total protein of the cells was extracted by complying with the instructions of radio-immunoprecipitation assay (RIPA) lysate (Beyotimi, Shanghai, China). Identical amounts of total protein (60 μg/lane) were electrophoresed on 10% or 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and then the protein was transferred onto the polyvinylidene difluoride membranes (Millipore, Bedford, MA) at low temperatures. After being blocked in 5% skimmed milk or 3% bovine serum albumin at ambient temperatures for 2 h, the cells were incubated with the primary antibody and the secondary antibody in sequence. Lastly, the enhanced chemiluminescence detection (ECL-Plus kit; Beyotime) was employed to detect the protein bands.
The mouse monoclonal antibodies against proliferating cell nuclear antigen (PCNA) (no. 2586), the rabbit monoclonal antibodies cyclin E (no. 20808), horseradish peroxidase-conjugated goat antirabbit (no. 7074), and goat antimouse (no. 7056) secondary antibodies originated from Cell Signaling Technology (CST, Beverly, MA.). The antibodies of rabbit anti-TGFβRII polyclonal antibody (Item number:abs135929), phospho-TGFβRII (Tyr284) rabbit polyclonal antibody (item number:abs140020), rabbit anti-Smad2 polyclonal antibody (item number:abs131983), rabbit anti-Smad3 polyclonal antibody (item number:abs131872), phospho-Smad3 (Ser423+Ser425) rabbit polyclonal antibody (item number:abs140144), and phospho-Smad2 (Ser465+Ser467) rabbit polyclonal antibody (item number:abs140143) were offered by Absin Biotechnology Co., Ltd (Shanghai, China).
Animals
This animal study was approved by the Animal Research Committee of Yangzhou University (Yangzhou, China). In total, 48 Sprague–Dawley male rats weighing from 250 to 300 g randomly fell into 4 groups (12 in the respective group), and each rat received a strict care.
Laminectomy modeling
The experimental model of laminectomy complied with the existing study (Dai and others 2019). In brief, after intraperitoneal injection of 1% pentobarbital sodium (40 mg/kg) to achieve the satisfactory anesthetic effect, the lumbar 1 position was located, the median skin incision was selected on the back, and the paraspinal muscles were carefully separated. Subsequently, the L1 vertebral lamina was removed, and ∼5 × 3 mm defect area of dura mater spinalis was exposed. After thorough hemostasis, the operative areas were washed with saline, and neurological sponges were fully saturated with saline, 1, 2, and 5 MU/mL of IFN-α-2b. Next, the samples were placed on the surgical area for ∼10 min. After the neurological sponges were removed, the surgical site was washed again with saline, and then the wound was sutured. Lastly, each rat was injected with penicillin (intramuscularly, 50 mg/kg,) to prevent the infection at the incision. Three times a week after the surgery, 50 μL of normal saline or corresponding concentration of IFN-α-2b was injected locally close to the operation area (Quiroga and others 2020).
Macroscopic assessment of EF
After 4 weeks, 6 rats in the respective group were randomly selected to conduct the macroscopic evaluation, as judged based on an existing study (Rydell 1970). Given the difficulty of anatomy, the adhesion degree fell to 4 levels, that is, from 0 to 3, corresponding to from no adhesion to firm adhesion, respectively.
Picric-sirius red polarized light method
The sections were deparaffinized in water and stained with sirius red for 1 h, rinsed fully with water, and stained with the Mayer hematoxylin staining solution for 8–10 min, and then washed with tap water for 10 min. Lastly, the gradient alcohol dehydration and xylene sealing tablets were used. The polarized light microscopy was employed to distinguish between type I and type III collagen.
Histological analysis
The other 6 rats in each group were selected to conduct the histological analysis. The rats were anesthetized successfully and then perfused with 4% paraformaldehyde through the intracardiac injection. The entire L1 vertebral body (e.g., adjacent paravertebral muscle and epidural fibrous tissue) was subsequently excised and overall soaked in 4% paraformaldehyde. After approximately 1 month of tissue decalcification, the specimens were embedded in paraffin. Besides, the vertebral body was cross-sectioned continuously at 4 μm, and hematoxylin and eosin staining was performed to observe the EF and the fibroblast count of the epidural defect. Furthermore, the collagen tissue content was stained with Masson's trichrome stain.
Immunohistochemical staining
After the sections were dewaxed and rehydrated with gradient alcohol, the antigenicity was activated for 15 min in 95°C sodium citrate solution, placed in 3% H2O2 to inhibit the activity of peroxidase, and then washed 3 times with PBS. Next, the primary antibody (e.g., diluted anti-PCNA, anticollagen I, anticollagen III, antiphospho-Smad2, and anti-TGFβ1) was incubated overnight at 4°C. Subsequently, the sections were incubated with antirat IgG for 1 h at ambient temperature, and the antibody binding was examined with the DAB kit. Lastly, hematoxylin staining was performed, and the results were observed under a light microscope.
Statistical analysis
In this study, SPSS 22.0 statistical software was adopted to analyze the data, and the data are expressed as mean ± standard deviation. The values of P < 0.05 were statistically significant in all analyses.
Results
IFN-α-2b inhibited cell proliferation in fibroblasts
The CCK-8 test was performed to assess the inhibitory effect of IFN-α-2b on cell viability. The treatment with IFN-α-2b at different concentrations in different periods was performed. As suggested from the results, IFN-α-2b could inhibit the activity of fibroblasts in a concentration- and time-dependent manner. IFN-α-2b gradually showed its inhibitory effect when treating fibroblasts for 24 h, and the cell survival rate declined significantly after 24 h (Fig. 1A). According to the cell cycle analysis, IFN-α-2b induced a significant accumulation of cells in the S phase, and then the distribution of GO/G1 and G2/M phases decreased (Fig. 1B, C). In addition, the inhibition effect of IFN-α-2b on the proliferation of fibroblasts was demonstrated with the Edu incorporation method (Fig. 1D, E). The results of the Western blotting assay presented that the expression levels of PCNA and cyclin E, 2 important cell proliferation markers, decreased in the IFN-α-2b treated group (Fig. 1F, G). As revealed from all the mentioned results, IFN-α-2b could inhibit the proliferation of fibroblast.

The inhibitory effect of IFN-α-2b on the growth and proliferation of fibroblasts.
IFN-α-2b on fibroblasts migration
The wound healing rate of the IFN-α-2b group obviously decreased as compared with the control (Fig. 2A, B). Moreover, the transwell migration assay proved that the number of cell migrations in the IFN-α-2b group was significantly lower than that in the control (Fig. 2C, D). Moreover, the TGFβ1 stimulation group could elevate the wound healing rate and the number of migrating cells, whereas IFN-α-2b could also restrain the promotion effect of TGFβ1 (Fig. 3A–D). As indicated from the mentioned findings, IFN-α-2b could inhibit cell migration.
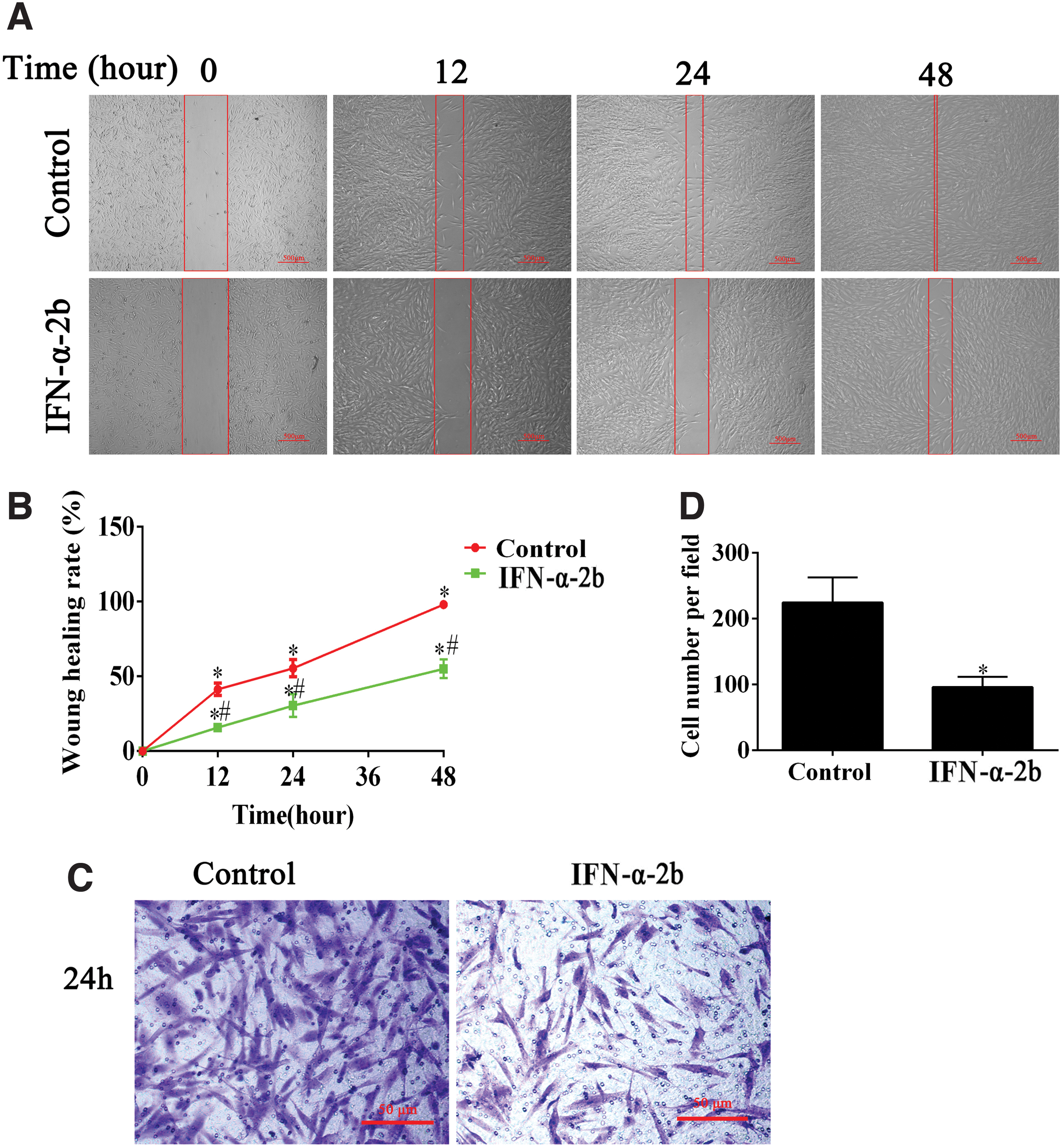
The inhibitory effect of IFN-α-2b on migration of fibroblasts.

Inverse effect of IFN-α-2b and TGF-β 1 on fibroblast migration.
IFN-α-2b inhibited the fibroblast proliferation through the TGFβ/Smad pathway
To verify whether the TGFβ/Smad pathway was involved in the regulation of the inhibitory effect of IFN-α-2b on fibroblasts, fibroblasts were treated with IFN-α-2b at different concentrations for 48 h, and then the expression levels of the related proteins were detected by performing the Western blotting assay. As indicated from the results, IFN-α-2b downregulated the expression of p-TGFβRII in a concentration-dependent manner. Moreover, IFN-α-2b downregulated the expression levels of p-Smad3 and p-Smad2 downstream of TGFβRII (Fig. 4A, B). To further verify the mechanism of IFN-α-2b inhibiting fibroblast proliferation, TGFβ1 was adopted to stimulate the cells. As indicated from the results of the Western blotting assay, TGFβ1 could promote cell proliferation and activate the TGFβ/Smad signaling pathway. It is noteworthy that the activation of TGFβ1 was reversed in the IFN-α-2b–treated group (Fig. 4C, D). In addition, the Edu incorporation method suggested that IFN-α-2b and TGFβ1 exerted the opposed effects on the DNA replication activity of fibroblasts (Fig. 4E, F). As revealed from all the mentioned results, IFN-α-2b could inhibit fibroblast proliferation by negatively regulating the TGFβ/Smad signaling pathway.

IFN-α-2b inhibits the activation of TGFβ/Smad signaling pathway, and the inhibition of this pathway may affect the proliferation of fibroblasts.
Macroscopic assessment of EF
The analysis suggested that except 1 rat in the control, the rest of the rats were grade 3, whereas grades 0, 1, and 2 were identified in the art treatment group. According to the Rydell's classification, the degree of EF was observed macroscopically. As reported from the analysis, except for 1 rat in the control, the rest of the rats were grade 3, whereas grades 0, 1, and 2 were reported in the art treatment group (Table 1).
The Grade of Epidural Scar Adhesion Through Macroscopic Assessment According to the Rydell Classification Standard
IFN-α-2b, interferon-alpha-2b.
IFN-α-2b reduces EF and prevents fibroblast proliferation
According to the images of HE staining, the local application of IFN-α-2b could decrease the degree of EF, and the loose fibrous tissue was not attached to the dura mater in the laminectomy area (Fig. 5A). It is noteworthy that the effect trend complied with the dosage. As indicated from the results, the number of fibroblasts in the scar tissue reduced with the increase in the IFN-α-2b concentration (Fig. 5A, B). The results of Masson staining revealed that the density of collagen decreased with the increase in the IFN-α-2b concentration, complying with the fibroblast counting (Fig. 5A, C). As reported from the immunohistochemical staining of type I and type III collagen in the fibrotic tissue, IFN-α-2b reduced the content of type I and type III collagen in the EF tissue (Fig. 5A, D, E). Moreover, the staining of epidural scar tissue was observed under a polarization microscope; as indicated from the results, the expression levels of type I collagen and type III collagen declined with the increase in the concentration of IFN-α-2b (Fig. 5A). According to the mentioned results, IFN-α-2b could reduce the synthesis of collagen in the epidural fibrous tissue, as well as the formation of EF.

Histological evaluation images of the laminectomy operation area treated with 1, 2, and 5 MU/mL IFN-α-2b and saline.
IFN-α-2b inhibits the EF through the TGFβ/Smad pathway
The expression levels of PCNA, p-Smad2, and TGF-β1 were further assessed through immunohistochemistry. The results of immunohistochemical analysis are presented in Fig. 6. The number of positive cells in the IFN-α-2b–treated group was significantly smaller than that in the control. The mentioned results further confirmed that the application of IFN-α-2b could inhibit the proliferation of fibroblasts, and the TGFβ/Smad signaling pathway might be involved.

Immunohistochemical staining images in EF tissues.
Discussion
Fibrosis refers to the formation of excessive fibrous connective tissue in organs or tissues during repair or reaction. After laminectomy, to repair local defects, inflammatory cytokines and growth factors can activate fibroblasts to secrete collagen and an extracellular matrix, which lead to excessive fibrosis and thus the formation of EF (Sun and others 2015; Song and others 2016). Accordingly, the drug inhibiting the activity of fibroblasts can effectively reduce EF.
Human IFN-α-2b acts as a glycoprotein capable of regulating the activity of numerous antiviral and antiproliferative genes, and the antiproliferative effect of IFN-α-2b can be exerted in 2 manners, that is, directly and indirectly. To be specific, the direct activity inhibits cell growth by blocking cell cycle, thereby inducing apoptosis or differentiation. Besides, the indirect activity is generated by activating immune cells, immunosuppressive cells, and natural killer cells, thereby inhibiting vascularized antiangiogenetic and inducing cytokines (Asmana Ningrum 2014). Existing studies have reported that TGF-β1 and IFN-γ exert opposed influences on multiple cellular functions (Sobral and others 2007; Pappritz and others 2018). Compared with IFN-γ, whose inhibitory effect against fibrosis has been extensively studied, the mechanism causing the antifibrosis effect of IFN-α-2b remains unclear. It was reported that IFN-α-2b could antagonize TGFβ/Smad-dependent hepatic fibrosis in rat by competitive binding of Stat1 and Smad3 to p300, thereby laying a molecular basis for the antifibrosis effect of IFN-α-2b (Inagaki and others 2003).
TGF-β is recognized as a multifunctional mediator, involved in regulating the proliferation, differentiation, apoptosis, and migration functions of numerous biological systems.
Three TGF-β isoforms (i.e., TGF-β1, TGF-β2, and TGF-β3) have been isolated and identified in mammals. The vital mechanism of cell response to cytokines refers to the regulation of cell surface receptor expression, and TGFβRII is critical to the receptor activation and the subsequent signal propagation (Xu and others 2012; Hu and others 2018). Smad2 and Smad3, the direct substrates of TGF-β receptor kinase, bind to Smad4 after phosphorylation, and the complex can then be transported to the nucleus for gene transcription. It is generally known that TGF-β/Smad signals are the main pathways for the occurrence of fibers such as renal fibrosis, liver fibrosis, and lung fibrosis (Katz and others 2016; Chen and others 2018; Eser and Jänne 2018). Recently, some studies have shown that the TGFβ/Smad pathway is also involved in the process of EF. The mentioned reports show that inhibiting the TGFβ/Smad signaling pathway can prevent or ameliorate fibrosis (Yang and others 2016; Huang and others 2018). However, as far as we know, our study is the first to describe that IFN-α-2b inhibits EF through this pathway.
In this study, we found that IFN-α-2b inhibits fibroblast proliferation and reduces EF after laminectomy in rats, possibly by inhibiting the TGFβ/Smad signaling pathway. In vitro, CCK-8 and Edu incorporation showed that IFN-α-2b inhibited proliferation and activity of fibroblasts in a concentration-dependent manner. The expression of cyclin E and PCNA was also decreased after treatment with IFN-α-2b at different concentrations, and flow cytometry indicated that IFN-α-2b induced cell accumulation in the S phase. The mentioned results demonstrated that IFN-α-2b could inhibit proliferation of fibroblasts. Besides, IFN-α-2b could inhibit the TGFβ/Smad signaling pathway and reduce the effect of TGF-β1 on fibroblast proliferation. In in vivo experiments, rats after laminectomy were treated with different doses of IFN-α-2b. Histological observation, fibroblast count, immunohistochemical staining, and collagen density analysis were employed to assess the effect of IFN-α-2b in reducing EF. Compared with the control, the fibrotic tissue in the IFN-α-2b-treated group was looser and the collagen density was also lower. Moreover, the number of fibroblasts in the laminectomy site decreased after treatment with IFN-α-2b. As indicated from the mentioned results, the IFN-α-2b could inhibit fibroblast proliferation and correspondingly reduce EF in rats. Immunohistochemical analysis also showed that TGF-β1 and p-Smad2 were downregulated in the IFN-α-2b–treated group. As indicated from the mentioned result, the TGFβ/Smad signaling pathway is involved in the inhibitory effect exerted by IFN-α-2b on EF.
Given the existing reports, IFN-α-2b has been explored as early as 1989 for treating keloid, and it has achieved good results (Berman and Duncan, 1989). As demonstrated from the subsequent studies, IFN-α-2b could be injected locally to treat keloid and burn scars. According to the existing results, IFN-α-2b could decrease the number of fibroblasts and the production of collagen, whereas it can inhibit the secretion of TGF-β1, which is considered a fibrogenic factor (Tredget and others 2000; Wang and others 2007; Lee and others 2008). Our experiment results showed that IFN-α-2b can also inhibit the proliferation of fibroblasts, reduce the content of TGF-β1, and reduce collagen production in the area of EF. Moreover, the experiments here confirmed that IFN-α-2b could inhibit fibroblast proliferation mainly in the S phase of the cell cycle, which was consistent with its effect on some tumor cells (Blank and others 2007; Katayama and others 2007).
However, it has been reported that IFN-α-2b treated rat palatal wound fibroblasts for 19 h. The results revealed that IFN-α-2b did not obviously reduce the proliferation of palatal fibroblasts. The reasons for such a result were analyzed to be probably the short time of drug treatment (Cornelissen and others 1999). The results of CCK8 in this study might provide some clues to explain the mentioned problem, and it was speculated that the effect of short-term application on inhibiting proliferation is not obvious. Thus, this study decided to treat for a longer time and should test before the cells reach fusion, since IFN-α-2b had an insignificant effect on near-confluent cells (Tredget and others 2000).
Existing studies have shown that IFN-α-2b is well tolerated and can be obtained even at high doses (Prata and others 1990), although in our treatment process, there were no side effects (e.g., delayed wound healing, epidermal necrosis, surgical incision infection, and death). However, whether there were any differences in the optimal concentration of local application, the frequency of application, and the distance between the injection area and the scar in the animal model, and whether the wound healing would be delayed when the concentration increased to a certain extent were not carefully explored, and there might be some potential side effects in other tissues around the injection site, side effects of the metabolism of the liver and kidney tissues. Even systemic toxicity and other issues are worthy of our consideration and in-depth study. In addition, the formation mechanism of fibrosis is significantly complex, including the proliferation and migration of fibroblasts. Whether there was apoptosis of fibroblasts at the identical concentration was not explored. Indeed, apoptosis can cause decline of cell viability. Accordingly, some deficiencies remained in this study, so further research is required before clinical application of IFN-α-2b.
Conclusion
In conclusion, this study preliminarily verified that IFN-α-2b can alleviate EF by inhibiting fibroblast proliferation. Moreover, the mentioned effects can be achieved by inhibiting the TGFβ/Smad signaling pathway. Our study provides a new therapeutic option for reducing EF after laminectomy.
Footnotes
Acknowledgments
We show great appreciation to all of our teachers for their generous help.
Ethical Approval
This experimental study was approved by the ethics committee and research committee of Clinical Medical College of Yangzhou University. The animal experiment was approved by the animal ethics committee of Yangzhou University, and all the rats received humanitarian care.
Author Disclosure Statement
We declare that we have no financial and personal relationships with other people or organizations that can inappropriately influence our study, there is no professional or other personal interest of any nature or kind in any product, service, and/or company that could be construed as influencing the position presented in, or the review of, the article entitled.
Funding Information
This study was supported by the National Natural Science Foundation of China (Grant No. 81772332). This study was also supported by Jiangsu Provincial Medical Innovation Team (Grant no. CXTDB2017004). Postgraduate Research and Innovation Program of Jiangsu Province (KYCX21_3285).