
Abstract
Sinapic acid (SA) was reported to protect against inflammation in various types of diseases. However, the role of SA in rheumatoid arthritis remains unclear. This study was designed to investigate the role of SA on rheumatoid arthritis. Rheumatoid arthritis mouse model was established by collagen immunization [collagen-induced arthritis (CIA)]. Histological analysis of articular cartilage tissue was carried out by hematoxylin and eosin (H&E) staining. Serum concentrations of tumor necrosis factor alpha and interleukin 6 were determined through enzyme-linked immunosorbent assay (ELISA). Oxidative damage indexes such as superoxide dismutase activity, malondialdehyde detection, glutathione detection, and catalase were determined by biochemical analysis. The protein levels of related genes were determined using Western blot. In CIA model, SA treatment attenuated paw swelling and clinical score of arthritis, attenuated articular cartilage tissues edema and infiltration of inflammatory cells, suppressed inflammatory cytokines release, and attenuated oxidative damage indexes. Mechanically, SA suppressed immune responses through inhibiting the IκB kinase (IKKs). SA attenuates rheumatoid arthritis through reducing inflammation and oxidative stress by downregulating IKKs.
Introduction
Rheumatoid arthritis (RA) is commonly believed to be a degenerative disease in joint tissues and the incidence of rheumatoid arthritis is increasing (Guo and others 2018; Zhai and others 2018a, 2018b, 2019). More and more people regarding of different races, genders, and ages are suffering from rheumatoid arthritis. The characters of rheumatoid arthritis include stiffness, pain, and swelling in joint tissues (Smolen and others 2016). Moreover, the development of rheumatoid arthritis will lead to damages in skin, lung, eye, heart, and kidney. Accordingly, patients with rheumatoid arthritis have a higher risk for disability and death (Naz and Symmons 2007). Currently, the strategies for rheumatoid arthritis treatment include sulfasalazine, methotrexate, hydroxychloroquine, leflunomide, tumor necrosis factor (TNF) inhibitors, anti-interleukin (IL) 6 receptor antibodies, and Janus kinase inhibitors (Smolen and others 2016). Among these strategies, methotrexate is the most commonly used based on its activity in suppressing immune responses and anti-inflammation. However, the effect is not so satisfactory because of its toxicity, which was reported to be harmful to gastrointestinal, respiratory, neuronal, cardiovascular, urinary, and immune systems (Romão and others 2014). Therefore, it is urgently needed to search for the more effective and less toxic therapeutic agents for rheumatoid arthritis.
Inflammation is widely accepted to be crucial for rheumatoid arthritis development (Demoruelle and others 2014; Zhai and others 2018b, 2019) and this is the theoretical basis for the design for many antirheumatoid arthritis medicines (Quan and others 2008). Sinapic acid (SA) is a phenolic acid that is widely present in plants, and it is commonly obtained from vegetables, fruits, and rye (Andreasen and others 2001; Lu and others 2001). The biological functions of SA in anti-inflammation were previously reported in various types of diseases. In rat chondrocytes, SA suppressed prostaglandin E2 and nitric oxide productions, inhibited in cyclooxygenase-2 and inducible nitric oxide synthase protein levels, and attenuated the mitogen-activated protein kinase pathway activation that were induced by IL-1β in vitro (Huang and others 2018). In 2,4,6-trinitrobenzene sulfonic acid-induced colitis model, SA treatment enhanced colon length, attenuated the colonic damage, suppressed myeloperoxidase activity and TNF alpha (TNF-α) expression in colonic tissues (Lee 2018). In vitro experiments showed that SA-pretreated human chondrocytes had the lower levels of IL-1β-induced prostaglandin E2 (PGE2), IL-6, nitric oxide (NO), and TNF-α. In vivo, SA could delay osteoarthritis progress in mice models (Li and others 2019). In destabilization of the medial meniscus (DMM)-induced osteoarthritis model, oral SA administration significantly decreased the histopathologic scores of knee articular cartilage and decreased inflammatory cytokines [IL-6, IL-1β, TNF-α, matrix metalloproteinase-1 (MMP-1), MMP-3] levels in cartilage (Cai and others 2019). These reports clearly demonstrated the anti-inflammation effect of SA. However, in rheumatoid arthritis, the effect of SA remains unclear.
In this study, we investigated the role of SA in rheumatoid arthritis using collagen-induced arthritis (CIA) model.
Methods
Cell culture
Rheumatoid arthritis fibroblast-like synoviocytes (RA-FLS) were first isolated from synovial tissues from RA patients. Written consent was obtained from the patients. The study was approved by Daqing Oilfield General Hospital (2019-035). Fresh synovial tissues were minced and digested in type I collagenase (Sigma-Aldrich, St. Louis). The cells were cultured with DMEM-Ham's F-12 (DMEM/F12) (Gibco, Life Technologies, China) and 20% fetal calf serum (FCS; Gibco, Life Technologies) in a 5% CO2 incubator. The morphology of FLS was confirmed under the light microscope and further characterized by flow cytometry using monoclonal antibodies CD90-PE and CD68-Alexa 647 (BD Biosciences, Franklin Lakes, NJ). Then, the cells were kept in DMEM (Invitrogen, Carlsbad) medium with 10% FCS (Gibco, Life Technologies), 10 mM HEPES (Thermo Fisher Scientific, Waltham), 2 mM
Collagen-induced arthritis
DBA/1 mice at the age of 8 weeks were purchased from Cyagen Biosciences (Suzhou, China). All mice were divided into 3 groups: control (Ctrl, treated with saline), CIA, and CIA+SA. CIA was performed as previously described (Pietrosimone and others 2015; Zhai and others 2019). DBA/1 mice were subcutaneously injected at the proximal tail using 100 μg bovine type II collagen (Gibco, Life Technologies) that was emulsified in Freund's complete adjuvant. On day 21, the mice were reinjected subcutaneously with 100 μg bovine type II collagen (Gibco, Life Technologies). For SA treatment, SA (Sigma-Aldrich; 100 mg/kg body weight) was orally gavaged on day 21 after immunization, followed by daily gavage for 5 weeks.
On day 21, the paw thickness was measured. The paws were scored following the characteristics: 0, normal, no inflammation or redness; 1, redness and swelling in 1 digit; 2, redness and swelling in >1 digit, or redness and swelling in 1 digit, ankle, wrist joint; and 3, redness and swelling in all digits and joints. Animal studies were approved by the Ethics Commitment of Daqing Oilfield General Hospital (jc-2019-21).
Histological analysis
Hematoxylin and eosin (H&E) staining was used for histological assessment. The joint tissues from different group mice were collected 49 days after immunization, then fixed using 10% neutral formalin for 24 h. Subsequently, the tissues were embedded in paraffin, and were sliced into 5-μm-thick sections. Then, H&E staining was performed following the standard protocols. The histological scores were assessed following the characteristics: 0, normal; 1, mild synovitis or slight cartilage erosion; 2, moderate synovitis and cartilage erosion; 3, erosion of the cartilage or bone destruction (Zhai and others 2016).
Enzyme-linked immunosorbent assay
On day 35 after immunization, the serums were collected from different group mice. The concentrations of TNF-α and IL-6 in serum were detected using the commercial enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis) following the manufacturer's protocols.
Biochemical analysis
On day 48 after immunization, synovial tissues from different groups were harvested and homogenized. The superoxide dismutase (SOD) activity, malondialdehyde (MDA) detection, glutathione (GSH) detection, and catalase (CAT) activity were detected following the published protocol (Zhai and others 2016; Erol and others 2019).
Luciferase assay
A total of 2 × 105 HEK293 cells/well were seeded into a 24-well plate. After 24 h, the cells were co-transfected with IL-6 promoter luciferase plasmid and IκB kinase (IKK) β or p65 using Lipofectamine® 2000 (Thermo Fisher Scientific). Cells were incubated with 5 or 10 μM SA, respectively, for 2 h before harvest. Forty-eight hours later, the Dual-Luciferase® Reporter Assay System (Promega, Madison) was used to detect luciferase activities.
Western blot
Western blot was carried out as described (Sun and others 2020a, 2020b). In brief, RA-FLS cells were collected and were lysed after stimulated with IL-1β for the different time periods with or without 10 μM SA. The lysed proteins were separated using 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). Then, the proteins were transferred to polyvinylidene fluoride (PVDF) membrane (Millipore, Burlington). Then, the PVDF membrane was blocked using 5% milk. Subsequently, the membrane was incubated using primary antibodies at 4° overnight: anti-p-IKK α/β (1:1,000; Abcam), anti-p-IκBα (1:1,000; Abcam), anti-IKKα (1:1,000; Abcam), anti-p65 (1:2,000; Cell Signaling Technology), anti-Lamin B (1:1,000; Abcam), and anti-Actin (1:2,000; Cell Signaling Technology). All the primary antibodies were from Cell Signaling Technology (Danvers). The membranes were incubated with horseradish peroxidase (HRP)-linked secondary antibody (1:3,000; Bioworld, ST. Louis Park, MN). Signals were detected using chemiluminescence.
Statistical analysis
Statistical analysis was performed using SPSS 17.0 software. All data were presented as mean ± standard deviation, as indicated in the figure legends. *P < 0.05, **P < 0.01, ns, not significant. One- and two-way analysis of variance (ANOVA) analysis with appropriate post hoc tests were used to determine the statistical significance. P < 0.05 was considered to be significant in all cases.
Results
SA attenuated paw swelling and clinical score of arthritis in CIA model
We first determined the role of SA in CIA mouse model. Supplementary data indicated that, when comparing with CIA group, administration of 50 mg/kg SA had moderate effects on degree of paw swelling (Fig. 1A) and clinical score of arthritis (Fig. 1B), but the effect was less significant than 100, or 150 mg/kg SA. Moreover, there was no significant difference of the effect between 100 and 150 mg/kg SA. We, therefore, chose 100 mg/kg SA for administration in this study. As shown in Fig. 1C, when comparing with control mice (Ctrl), the paw swelling degree of CIA mice significantly increased 4–8 weeks after immunization. In the established CIA model, SA treatment dramatically decreased the paw swelling degree 4–8 weeks after immunization. Consistently, the clinical score of arthritis analysis indicated that 5–8 weeks after SA treatment, the severity of arthritis in CIA model significantly decreased (Fig. 1D). These results demonstrated that SA attenuates paw swelling and clinical score of arthritis in CIA model.
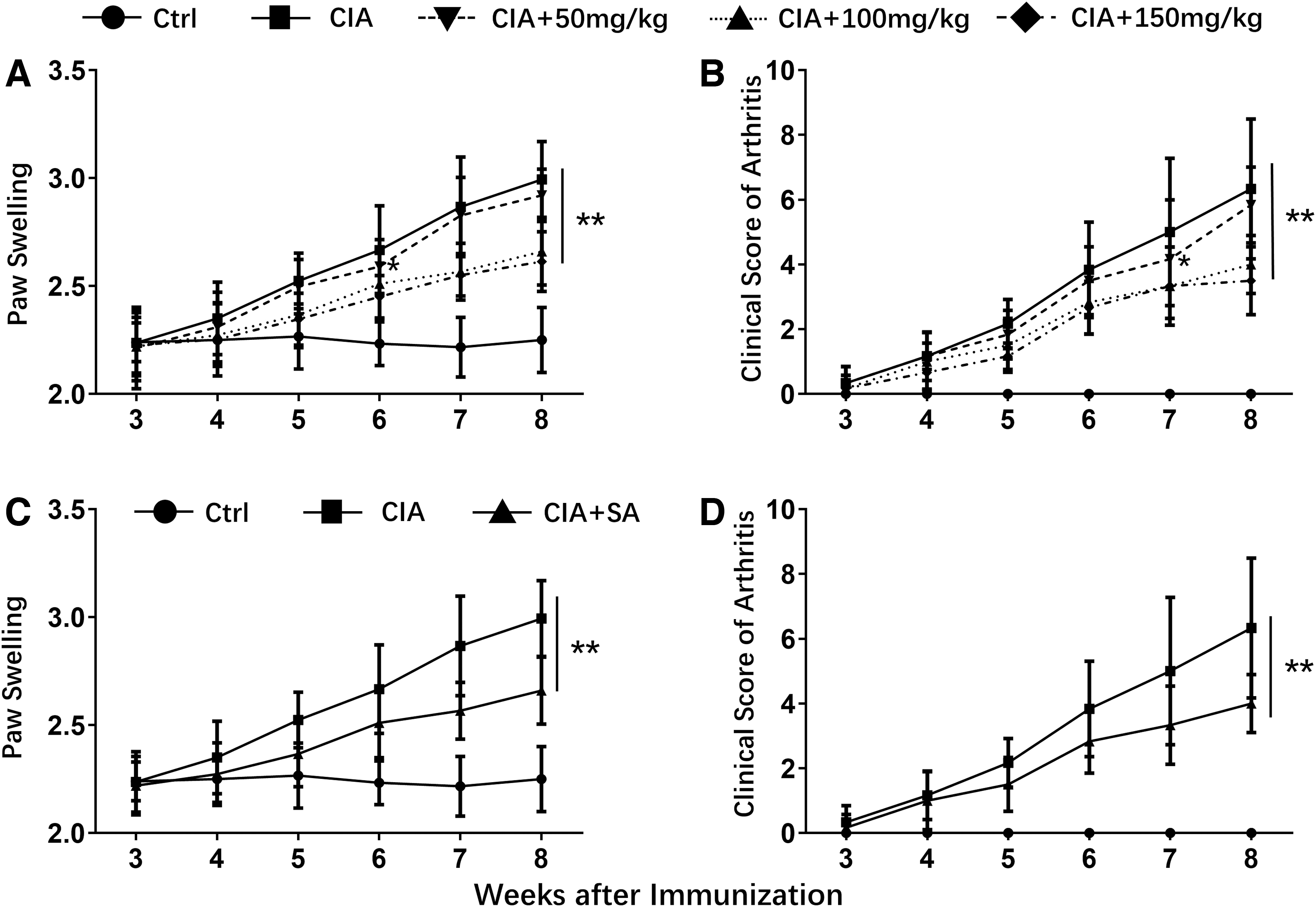
Effect of SA on degree of paw swelling
SA attenuated articular cartilage tissues edema and infiltration of inflammatory cells in CIA model
To further confirm the effect of SA on CIA, we performed the histological analysis using articular cartilage tissues from control, CIA, and SA-treated CIA mice. As shown in Fig. 2A, when comparing with control mice, articular cartilage tissues edema, increased infiltration of inflammatory cells, and enhanced angiogenesis in articular cartilage tissues were observed in CIA mice. SA treatment dramatically attenuated these phenotypes. The analysis of histological scores of the articular cartilage tissues from control, CIA, and SA-treated CIA mice demonstrated that SA treatment in CIA mice significantly decreased the histological scores (Fig. 2B). These results indicated that SA treatment significantly attenuated articular cartilage tissues edema and infiltration of inflammatory cells in articular cartilage tissue of CIA model.

Effect of SA on the articular cartilage tissue of CIA immunized mice (200 × magnification).
SA suppressed inflammatory cytokines release in CIA model
Inflammation is crucial for rheumatoid arthritis development (Choy and Panayi 2001). Accordingly, we determined the effect of SA on inflammatory cytokines release in the CIA model. CIA development led to significantly enhanced concentrations of TNF-α, IL-6, and IL-1β in serum as compared with control mice. SA treatment dramatically suppressed TNF-α, IL-6, and IL-1β in serum of CIA mice (Fig. 3A–C). These results demonstrated that SA suppressed inflammatory cytokines release in CIA model.
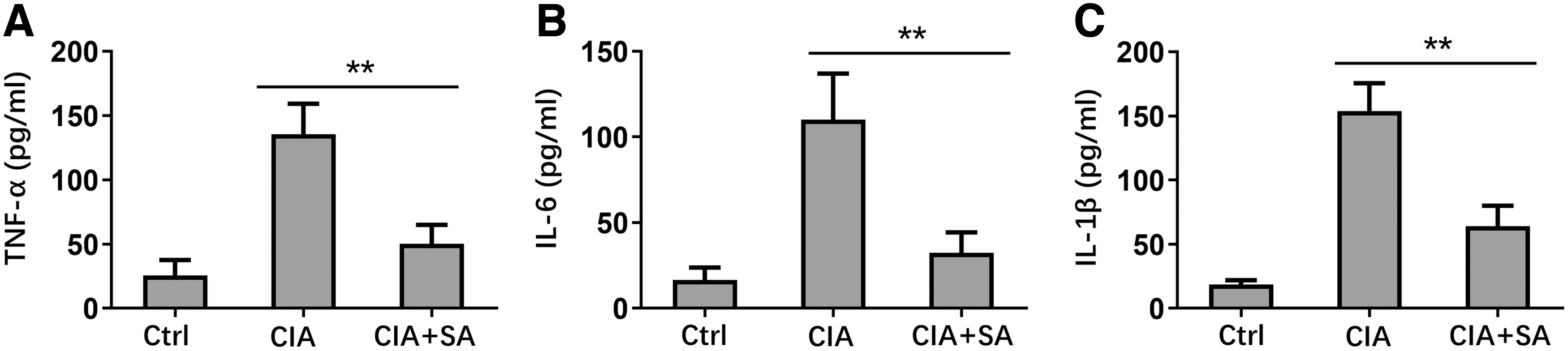
Effect of SA on decreasing
SA attenuated oxidative damage indexes in CIA mice
Oxidative damage is also crucial for rheumatoid arthritis development (Vasanthi and others 2009; da Fonseca and others 2019), and we determined the oxidative damage related indexes in synovial tissues of control, CIA, and SA-treated mice. CIA development significantly decreased the concentrations of SOD, GSH, and CAT in synovial tissues, whereas SA treatment significantly increased SOD, GSH, and CAT in CIA mice (Fig. 4A, C, D). MDA concentration significantly increased in synovial tissues of CIA mice, whereas SA treatment dramatically decreased MDA concentration in synovial tissues of CIA mice (Fig. 4B). These results demonstrated that SA treatment attenuated oxidative damage indexes in CIA mice.
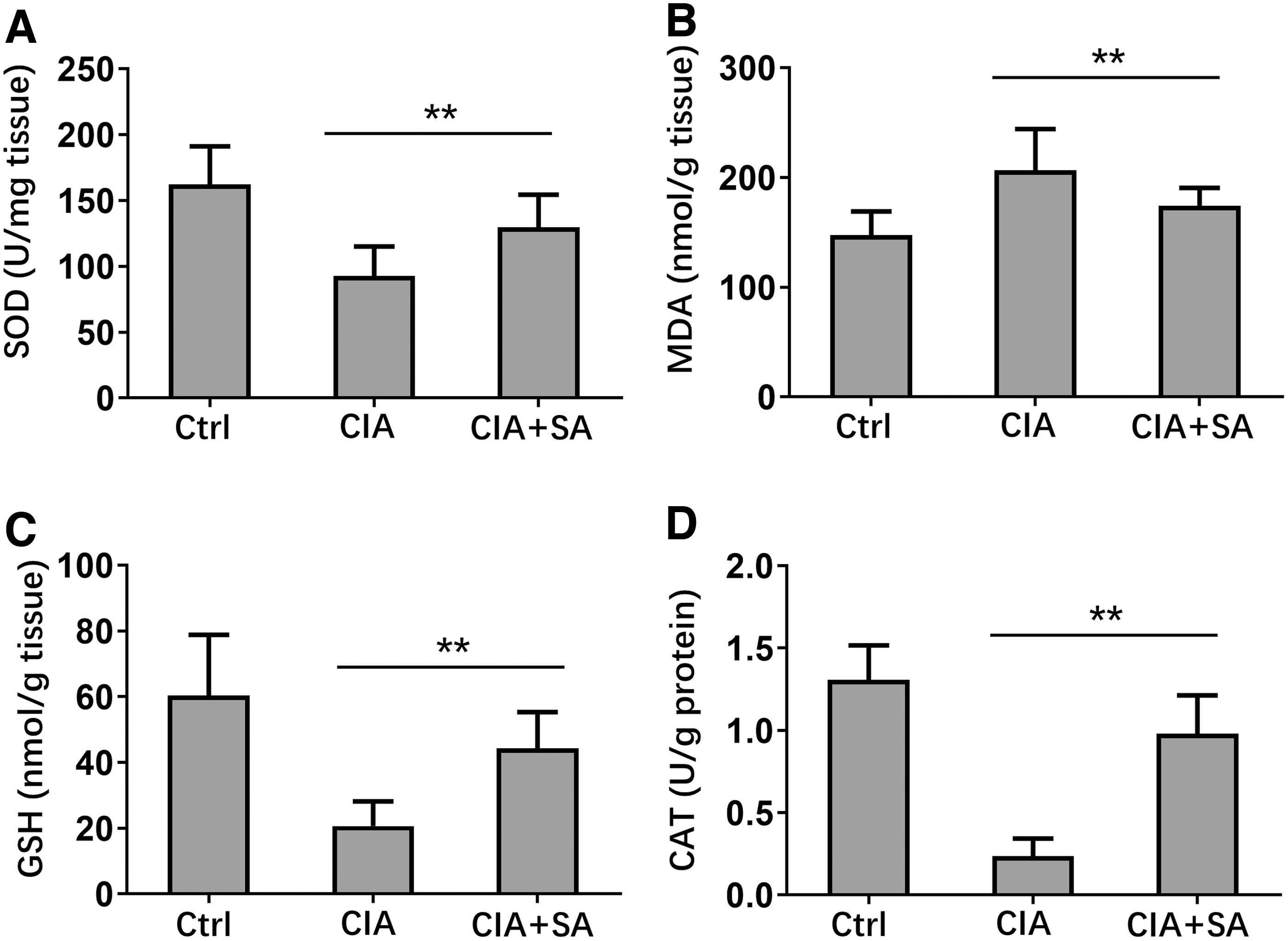
Effect of SA on inhibits oxidative stress markers in joint tissue. On day 48, the synovial tissues were collected from each group and made into tissue homogenate.
SA suppressed immune responses through inhibiting IKKs
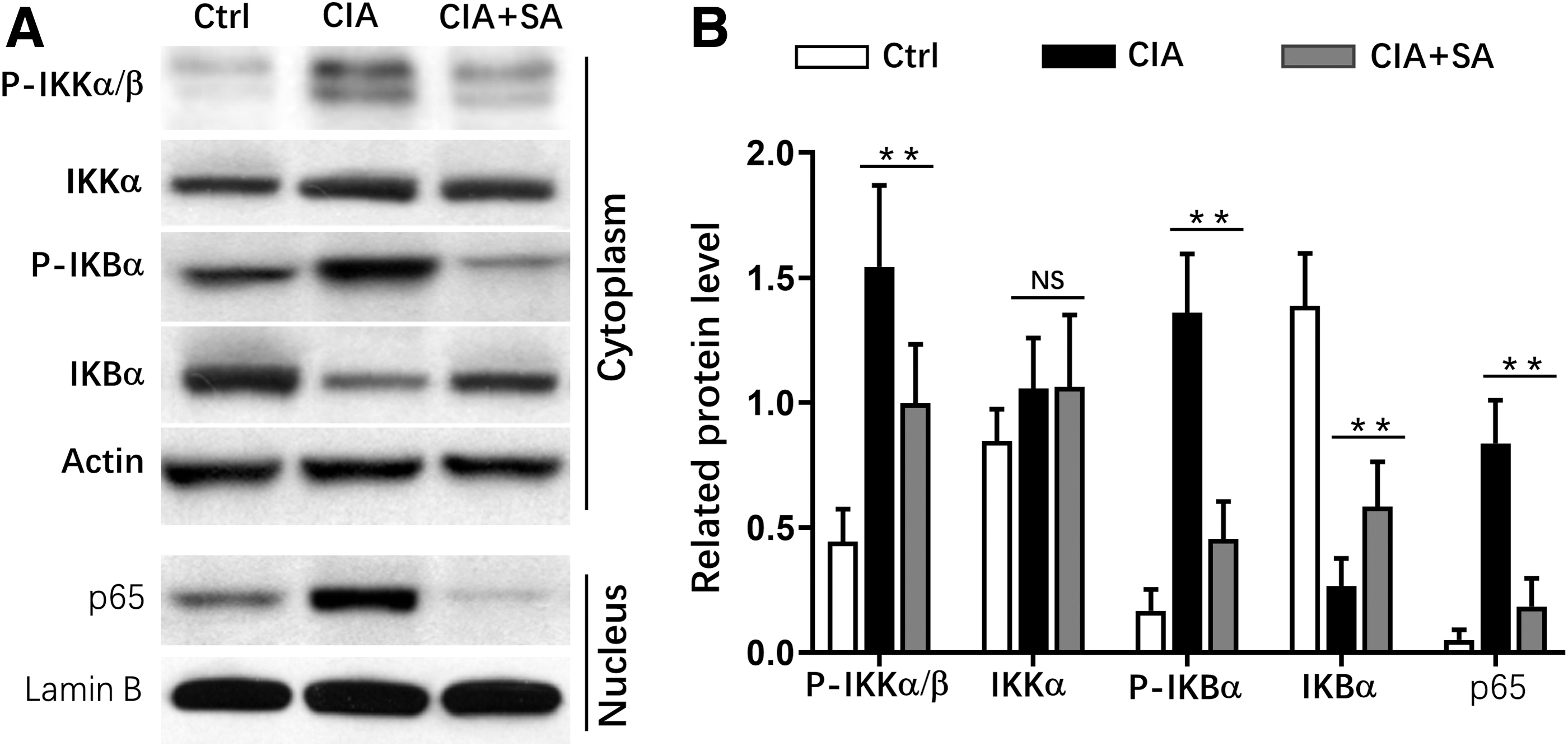
We further explored the mechanism of how SA suppressed immune responses in CIA mice. RA-FLS cells were stimulated by IL-1β, and this stimulation led to significantly increased expression levels of p-IKKα/β, p-IKBα, and p65 in RA-FLS at both 15 and 30 min. SA treatment dramatically suppressed the expression levels of p-IKK α/β and p-IKB α in cytoplasm of RA-FLS, and also decreased p65 expression in nucleus (Fig. 5A–E). To confirm which the target of SA was, we did luciferase assay. The results indicated that SA treatment significantly suppressed IKKβ activity in a dose-dependent manner. However, SA treatment showed no influence on p65 activity (Fig. 5F). These results demonstrated that SA suppressed immune responses through inhibiting IKKs.
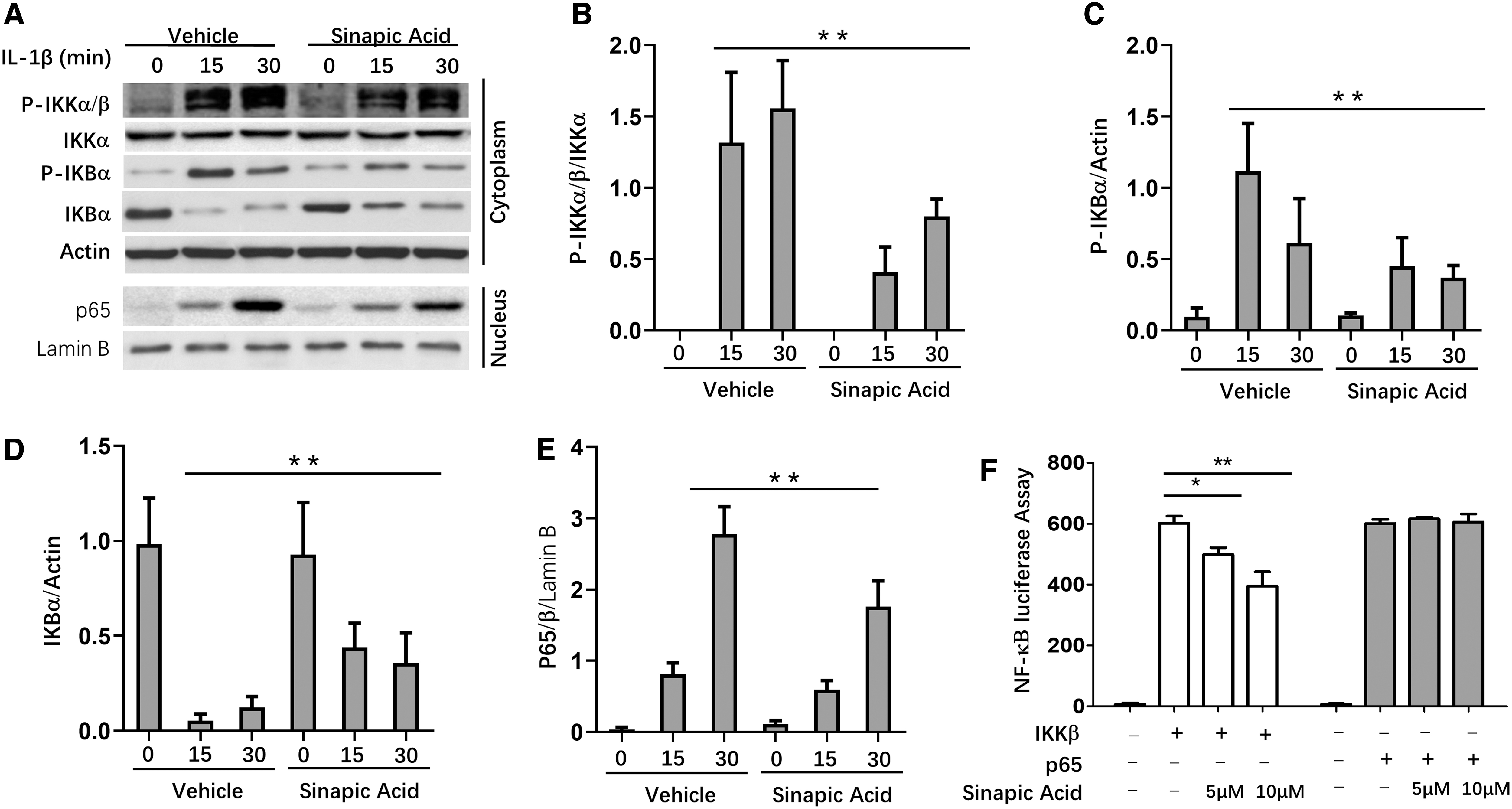
Activation of NF-κB is impaired in RA-FLS in response to SA treatment.
SA inhibited nuclear factor kappa B activation
We finally detected the effect of SA on nuclear factor kappa B (NF-κB) activation in synovial tissues of CIA model. CIA development significantly enhanced the expression levels of p-IKKα/β, p-IKBα, and p65, and decreased the expression of IKBα in synovial tissues. SA treatment dramatically suppressed the expression levels of p-IKKα/β, p-IKBα, and p65, and enhanced IKBα expression in synovial tissues (Fig. 6A, B). These results demonstrated that SA inhibited NF-κB activation in synovial tissues of CIA model.
Activation of NF-κB is impaired in synovial tissues in response to SA treatment.
Discussion
The effect of SA in rheumatoid arthritis was not investigated, so we initiated this study to answer the important question. We used DBA/1 mice to successfully established CIA model, which is the most used model for rheumatoid arthritis (Trentham and others 1977; Williams 2004). In this CIA model, we found that the paw swelling degree and the arthritis significantly clinical score increased as compared with control mice. Histological analysis using H&E staining showed that CIA development results in articular cartilage tissues edema, enhanced angiogenesis, and increased infiltration of inflammatory cells into articular cartilage. The inflammatory cytokines concentrations in serum dramatically increased in CIA mice. Moreover, CIA development led to oxidative stress in joint tissue. SA treatment by oral gavage significantly reversed the phenotypes in CIA mice. These observations suggested that SA treatment protected CIA mice by suppressing inflammation and attenuating oxidative stress.
Rheumatoid arthritis is also widely believed to be a chronic inflammatory joint disease. Therefore, inflammation is definitely crucial for rheumatoid arthritis development. In CIA model, there are obvious inflammation, which was detected by histological analysis and ELISA. Previous studies already indicated the importance of inflammation in rheumatoid arthritis (Choy and Panayi 2001), and accordingly many medicines for rheumatoid arthritis treatment were based on anti-inflammation. Our study clearly showed that SA treatment significantly attenuated the infiltration of inflammatory cells into articular cartilage tissue, inhibited inflammatory cytokines release in the serum, and finally decreased the degree of paw swelling in CIA mice. Consistent with observations, previous studies demonstrated that SA administration significantly suppressed inflammation in animal models of various diseases and human peripheral blood mononuclear cells (Raish and others 2018; Yang and others 2019; Alaofi 2020; Bin Jardan and others 2020; Olszewska and others 2020). In streptozotocin-induced diabetic nephropathy model, SA pretreatment (20 and 40 mg/kg bodyweight) both prevented the inflammatory cytokines release, and this effect is in a dose-dependent manner (Alaofi 2020). In acute doxorubicin-induced cardiotoxicity animal model, daily SA administration dramatically suppressed proinflammatory cytokines levels, including TNF-α, endothelin-1, IL-1β, and NF-Κb (Bin Jardan and others 2020). In lipopolysaccharides (LPS)-stimulated human peripheral blood mononuclear cells, SA treatment significantly decreased the proinflammatory cytokines release, and increased the anti-inflammatory cytokine (IL-10) release (Olszewska and others 2020).
Oxidative stress is another important accelerator for rheumatoid arthritis development (Balogh and others 2018; da Fonseca and others 2019). When determining the oxidative damage markers in the peripheral blood from rheumatoid arthritis patients, García-González and others (2015) found that oxidative damage of rheumatoid arthritis patients significantly increased as comparing with healthy control. Moreover, the effect of oxidative stress in rheumatoid arthritis was also explored. In human rheumatoid arthritis synovial fibroblast cells and synovial tissues from rheumatoid arthritis patients, oxidative stress induced by 4-hydroxy-2-nonenal damaged the energy metabolism, and also contribute to inflammation (Balogh and others 2018). These data indicated the association between oxidative stress and rheumatoid arthritis. In contrast, the strategies of inhibiting oxidative stress were also reported to be effective to attenuate rheumatoid arthritis. Dexamethasone is a widely used medicine for rheumatoid arthritis treatment based on its role in anti-inflammation. In CIA Wistar rat model, intraperitoneal injection of dexamethasone suppressed reactive oxygen species generation and attenuated arthritis intensity (Wang and others 2020), suggesting that the effect of dexamethasone in CIA was achieved by suppressing oxidative stress. Consistently, our study demonstrated that in the established CIA model, SA treatment significantly suppressed the oxidative stress, and attenuated rheumatoid arthritis, indicating the important role of SA-medicated oxidative stress reduction in rheumatoid arthritis.
Mechanically, we found that SA treatment dramatically suppressed the expression levels of p-IκK α/β and p-IκB α in RA-FLS. Moreover, luciferase assay indicated that SA treatment significantly suppressed IKKβ activity. These results indicated that SA suppressed immune responses through inhibiting IKKs. Consistently, in LPS-stimulated RAW 264.7 macrophages, SA treatment inhibited inflammatory cytokines expression at mRNA and protein levels through suppressing the p-IκB expression level and the nuclear translocation of NF-κB subunits (Yun and others 2008). All the data indicated that IKKs signal pathway was crucial for the protecting effect of SA in rheumatoid arthritis.
It is important to note the limitations of this study. Only CIA-induced rheumatoid arthritis mouse model was used in this study. Another rheumatoid arthritis animal model would be useful to confirm the effect of SA in attenuating rheumatoid arthritis.
In summary, we found that in the established CIA model, SA suppressed paw swelling degree, inhibited inflammatory cells infiltration, and attenuated inflammatory cytokines release. Mechanically, the effects of SA in CIA model were achieved through downregulation of IKKs.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.