
Abstract
This study was performed to evaluate the effectiveness of mesenchymal stem cells (MSCs) on bone healing and to assess the role of various chemical stimulants and mediators in healing. Forty female mice were randomly assigned to 4 groups (10 mice each) after the induction of fixed fractures: group I: received fixation only; group II: received phosphate-buffered saline (PBS); group III: received intralesion MSCs (IL-MSCs); and group IV: received intraperitoneal MSCs (IP-MSCs). Serum alkaline phosphatase (ALP) levels and the expression of the osteocalcin (OCN), bone morphogenetic protein-2 (BMP-2), and stromal-derived factor-1 (SDF-1) genes were measured. ALP reached baseline level only in IL-MSCs, whereas OCN reached baseline level in MSCs recipients (IL-MSCs and IP-MSCs). BMP-2 significantly increased in MSCs recipients 3 weeks postfracture and increased in all groups 8 weeks postfracture with significant increases in MSC recipients than the fixation and PBS groups. The highest BMP-2 expression was reached in IL-MSC group. MSCs either locally or systemically improves or accelerates the healing of bone fractures with better results obtained after local injection, as shown by biochemical, radiological, and histological findings. MSCs are effective candidates for bone regeneration.
Introduction
Bone has a characteristic ability to maintain viability under mechanical stress and external compression. This property decreases with advancing age (Mehta and others 2010). Fractures, one of the most frequent musculoskeletal injuries, have morbidity and mortality in all age groups. After injury, fibrotic scarring and incomplete tissue restoration occurs. However, fracture healing, a regenerative process, returns structure and composition to normal. During fracture healing, multiple cellular lineages are recruited and contribute to regeneration (Gerstenfeld and others 2003). Small damage can repair spontaneously but marked damage needs surgery. Autologous bone transplantation was the best for bone repair but its access is limited and it is associated with morbidity, pain, and infection at the donor site (Kang and others 2014).
Fracture healing involves an anabolic phase with new tissues formation, and a catabolic phase with tissue remodeling. Delayed healing and nonunion are common problems. Mechanical factors in the fracture area and multiple patient and fracture factors affect the healing outcome (Mirhadi and others 2013).
Mesenchymal stem cells (MSCs) are pluripotent, nonhematopoietic, and self-renewing cells. Many sources for MSCs were recognized like bone marrow (BM), adipose tissue, amniotic fluid, umbilical cord blood, and even breast milk (Shao and others 2015). MSCs are able to differentiate into mesodermal, endodermal, and ectodermal cells promoting structural and functional repairs (Wang and others 2013). MSCs are capable of secreting growth factors and immune-protective cytokines used in cell and organ transplantation. MSCs are safe and do not form teratoma. In the healing process, both local and circulating MSCs cooperate. Circulating MSCs are attracted to the site of injury to help tissue repair (Zhao and others 2016).
MSCs are responsible for the formation of osteoblasts, chondroblasts, and other stromal cells. These cells are important for the homeostasis of bone tissue and bone repair (Tong and Li 2006). The extracellular matrix (ECM) delivers cytokines secreted from different cells. Many cytokines are involved in the healing process of bone. Among them, stromal cell-derived factor-1 (SDF-1) and bone morphogenetic proteins (BMPs) participate in the recruitment of MSCs and induce their osteogenic differentiation (Bahney and others 2019).
BMPs are members of the transforming growth factor-beta (TGF-β) superfamily. TGF-β is a growth factor that can induce the proliferation and differentiation of pluripotent cells (Toosi and others 2019). Undifferentiated MSCs release BMPs to promote angiogenesis, chemotaxis, mitogenesis, and induce cell differentiation into osteoblasts and chondroblasts and promote osteoblast proliferation (Malizos and Papatheodorou 2005). They also stimulate SDF-1 that induces migration of MSCs to bone. BMP-2 binds with type I and type II serine-threonine kinase receptors and activates the Smad and mitogen-activated protein kinase (MAPK) pathways, inducing significant osteogenic effects (Katagiri and Watabe 2016). BMP-2 effects are mediated by upregulating the expression of runt-related transcription factor 2 (RUNX2).
The ECM contains collagen fibers and noncollagenous proteins, which include osteocalcin (OCN). OCN is a bone-related transcription factor that is involved in the formation of the ECM and collagen mineralization (Zhu and others 2021). The phosphate is up taken by stem cells to form adenosine triphosphate (ATP), which activates the intracellular signaling pathways through autocrine and paracrine mechanisms. This activates the expression of OCN and enhances osteogenic differentiation (Shih and others 2014).
Biochemical bone metabolic markers reflect the healing stages either as bone formation markers that indicate osteoblastic activity, or as bone absorption markers that indicate osteoclastic activity. Alkaline phosphatase (ALP) is responsible for mineralization of the ECM. Moreover, ALP mRNA and protein expression can be used to describe osteogenic differentiation progression (Maroon and others 2004). OCN is a commonly used marker of osteogenic differentiation, as well (Aubin 2001).
Thus, this study was performed to evaluate the effectiveness of MSCs on bone healing. Besides, we studied many chemical mediators that stimulate stem cell migration to fractured bone as SDF-1 and BMP-2 and bone formation and differentiation markers (ALP, OCN, and BMP-2).
Methods
This study was performed in the Molecular Biology and Biotechnology Unit, Medical Biochemistry Department, Faculty of Medicine, Benha University, Egypt and the Stem Cell and Clinical Chemistry Laboratory, Medical Biochemistry and Molecular Biology Department, Faculty of Medicine, Zagazig University, Zagazig, Egypt. This study received IRB approval from Research Ethics Committee, Faculty of Medicine, Benha University on 12/1/2015.
Preparation of BM-derived MSCs
Mouse MSCs were isolated according to Pulavendran and others (2010). BM cells were collected by flushing the femurs and tibias of 6-week-old BALB/c male mice. Cells were cultured at a concentration of 5,000/cm2/0.2–0.3 mL of media and then incubated at 37°C in 5% humidified CO2 in a CO2 incubator (Heraeus, Langenselbold, Germany). Nonadherent cells were eliminated every 2 days, and the whole medium was replaced with fresh medium. The cells were grown for 2–3 weeks until 80%–90% confluence. The whole adherent cells were detached by trypsinization with 0.25% trypsin/ethylenediaminetetraacetic acid (EDTA) (trypsin, 1:250, EDTA, 1 mM; Lonza Bioproducts) and then were replated. Third-passage cells were used for experiments. Cell cultures were routinely assessed with an inverted microscope, and cell viability was determined by means of trypan blue staining.
Determination of surface markers of BM-MSCs was carried out through evaluation of the positive expression of nonhematopoietic origin (using monoclonal antibodies that recognize an epitope on cluster of differentiation 105 [CD105]) and the negative hematopoietic marker expression, CD34, which were analyzed by flow cytometer according to Calabro and others (2003).
MSC staining with PKH26 fluorescence was carried out according to Garnier and others (2012). MSCs were labeled with a fluorescence marker using PKH26 Fluorescent Cell Linker Kit (Sigma, Aldrich) before being injected. Then, detection of homing into tissues of injected groups was performed by using a fluorescence microscope (Leica).
Induction of bone fracture
The animals were deeply anesthetized; closed mid-diaphyseal unilateral fractures of the femur were performed. External fracture fixation was performed using splint (wooden sticks and adhesive plaster). Then, mice were allowed free movement and limb weight-bearing (Supplementary Fig. S1).
Animal model and experimental design
Forty healthy BALB/c female mice of matched age and weight were included in this study. Mice were randomly divided after induction of fixed fractures into 4 groups (10 animals each): group I, mice with fixation only; group II, mice received phosphate-buffered saline (PBS); group III, mice received intralesion MSCs (IL-MSCs); and group IV: mice received intraperitoneal MSCs (IP-MSCs).
Blood samples (0.5 mL) were collected from the periorbital vein of mice under complete aseptic conditions in EDTA containing tube, before induction of fracture (baseline data), at the first day (d), the third-week (3w) and the eighth-week (8w) postfracture. Blood was used for ALP determination and quantitation of gene expression by real-time PCR.
X-ray imaging of the fracture site in mice was performed 1 day, 3 weeks, and 8 weeks postfracture. The procedure was performed under anesthesia using diagnostic x-ray unit (Ecoray, Korea). Mice from each group were killed 3 and 8 weeks postfracture and the fractured femurs were dissected and processed for histology and hematoxylin and eosin (H&E) staining.
Injection of MSCs into mice
The third passage MSCs were washed twice with PBS (Lonza Bioproducts) and the cells were trypsinized with 0.25% trypsin in 1 mM EDTA for 5 min at 37°C. Fluorescent-labeled MSCs were injected in a dose of 1 × 106 cells locally in the fracture site in mice of group III and IP in group IV.
Quantitation of target gene mRNA expression by real-time PCR
Total RNA extraction: from 100 μL EDTA mouse blood using Direct-zol™ RNA MiniPrep (Zymo Research), with Trizol reagent and DNase 1 digestion following the manufacturer's instructions. Eluted RNA (50 μL) was quantified by Ultraviolet Nanodrop 2000 Spectrophotometer (Thermo Fisher Scientific, Wilmington, NC): absorbance at 260 and 280 nm was measured to give an estimate of RNA purity (Wilfinger and others 1997). Conversion of RNA into cDNA was performed using SensiFAST™ cDNA synthesis kit (Bioline Reagents Ltd, United Kingdom) in Veriti™ Thermal Cycler (Applied Biosystems, Singapore). The reaction included total RNA (5 μL), 5 × TransAmp buffer (4 μL), reverse transcriptase (1 μL), and up to 20 μL nuclease free water with the thermal program; 25°C for 10 min, 42°C for 15 min, and 85°C for 5 min.
Then quantitation of target gene expression was performed in Stepone Real-Time PCR System (Applied Biosystems). Singleplex reactions were performed. The primer sequences of the respective mice genes were as follows: OCN forward: 5′-CCGGGAGCAGTGTGAGCTTA-3′, reverse: 5′-AGGCGGTCTTCAAGCCATACT-3′ (Zhu and others 2011), BMP-2 forward: 5′-TGGAAGTGGCCCATTTAGAG-3′, reverse: 5′-TGACGCTTTTCTCGTTTGTG-3′) (Jensen and others 2010), SDF-1 forward: 5′-TTCCGCTTCTCACCTCTGTA-3′, reverse: 5′-TGGTTAATTCTAGGCATGTTCTC-3′ and β-actin forward: 5′-CTAAGGCCAACCGTGAAAAGA-3′, reverse: 5′-CCAGAGGCATACAGGGACAAC-3′) (Zhang and others 2014). A real-time PCR was performed with addition of 2× SensiFAST Syber Hi-Rox Kit (Bioline Reagents Ltd) (10 μL), forward primer (0.8 μL), reverse primer (0.8 μL), cDNA (2 μL), and up to 20 μL nuclease-free water. The thermal program was 95°C for 5 min (holding), cycling (40 cycles: denaturation; 95°C for 15 s, annealing; 56°C, 1 min for OCN, 60°C, 1 min for BMP-2, and 54°C for SDF-1 and extension; 72°C for 20 s). Melting curve analysis was performed in each run to confirm specificity of real-time PCR assay.
The relative quantities of the target genes are normalized against the relative quantities of the β-actin gene. Fold expression changes are calculated using the equation 2−ΔΔCT (Livak and Schmittgen 2001).
Histopathological analysis of fractured femurs
Mice were anesthetized with ether inhalation, and the femur of each mouse was dissected out carefully. Femur specimens were fixed in 10% formalin solution and processed to prepare 5 μm thick paraffin sections for H&E stains (Bancroft and Gamble 2002).
Statistical analysis
All statistical analysis was performed using the SPSS software (Statistical Package for the Social Sciences, version 20.0; SPSS, Inc., Chicago, IL). Quantitative variables were demonstrated as mean ± standard deviation. Comparison within the studied groups was performed by paired t-test, whereas between groups by means ANOVA with least significant difference. Values of P < 0.05 were statistically significant.
Results
Characterization of cultured MSCs
Morphologically, MSCs were characterized by their attachment to plastic and fusiform shape. These cells had a spindled, fibroblast appearance after expansion. Flow-cytometric analysis of cell surface markers in MSCs expressed CD105 but did not express CD34. The surface marker expression pattern corresponds to BM-MSCs (Fig. 1).

Flow cytometric analysis of cell surface markers in MSCs: stem cells were negative for CD34 and positive for CD105. This expression pattern corresponds to bone marrow MSCs. MSCs, mesenchymal stem cells. Color images are available online.
Detection of homing
Concerning the analysis of MSCs homing in the femurs after MSCs transplantation, detection of PKH26-labeled MSCs in the femur of recipient female mice was performed by means of a fluorescent microscope (Leica). MSC-treated tissues showed PKH26-labeled MSCs as bright dots in the blood vessels and the connective tissue. The highest percentage of these cells was in group III. However, complete absence of these bright dots was detected in non-MSC recipient control groups (Fig. 2).
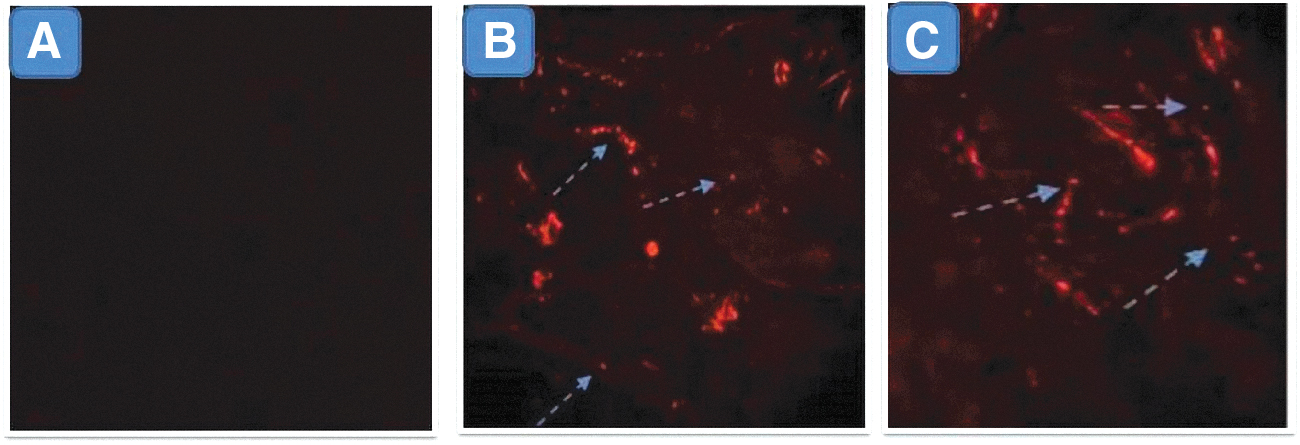
Tracking of transplanted MSCs in fractured bone:
ALP results
Three weeks postfracture, a significant increase in all groups was detected (P < 0.05), and IL-MSC group was significantly higher than other groups (P < 0.05), whereas there was nonsignificant difference between the other groups. Eight weeks after fracture, there were significant decreases compared with 3-week levels in all groups (P < 0.05) and group III (IL-MSCs) was the only one that reached the baseline level (Fig. 3A).

Gene expression results
Three weeks postfracture, there was a significant increase in OCN gene expression in all groups compared with baseline (P < 0.05) with nonsignificant difference between IL and IP-MSC groups. However, it was significantly higher in group III compared with groups I and II (P < 0.05). Eight weeks postfracture, nonsignificant difference was detected compared with baseline in MSC groups (III and IV) only and the baseline level had been reached in both groups, as well (Fig. 3B).
Compared with baseline, there was significant increased BMP-2 gene expression in MSC-recipient mice (groups III and IV) 3 weeks postfracture (P < 0.05 for each). Eight weeks postfracture, BMP-2 significantly increased in groups I and II (P < 0.05) compared with the 3-week level but still significantly lower than MSC-recipient mice (groups III and IV). Eight weeks postfracture, there was a significant decrease in group IV (P < 0.05) but nonsignificant difference in group III. The highest level of BMP-2 had been reached in group III (Fig. 3C).
Compared with baseline, there was a highly significant increase in SDF-1 gene expression as regard all groups at 1 day and 3 weeks postfracture (P < 0.05), whereas there was nonsignificant difference on comparing levels either 3 weeks or 1 day postfracture. There was no significant difference between MSC-recipient groups concerning 1-day levels (P = 0.91) and 3-week levels (P = 0.87), as well (Fig. 3D).
Correlation of serum ALP levels with other parameters
There were significant positive correlations between each of OCN and BMP-2 gene expression and serum ALP levels 3 and 8 weeks postfracture in the studied groups. Besides, a significant positive correlation between SDF-1 gene expression and serum ALP levels was found at 3 weeks postfracture in the studied groups (Figs. 4 and 5).
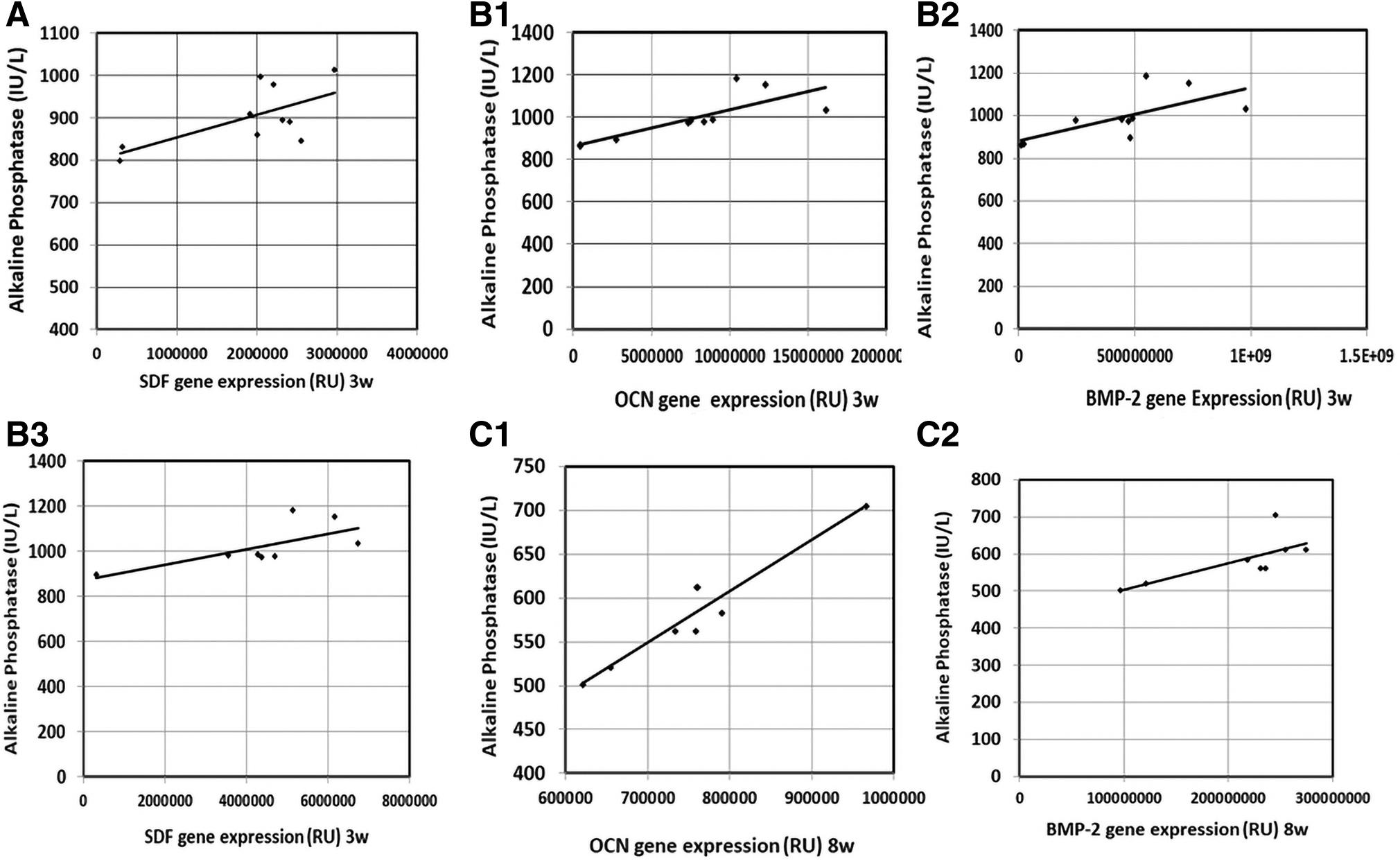
Correlation of serum ALP levels with gene expression of the studied parameters in the intra-lesional group.
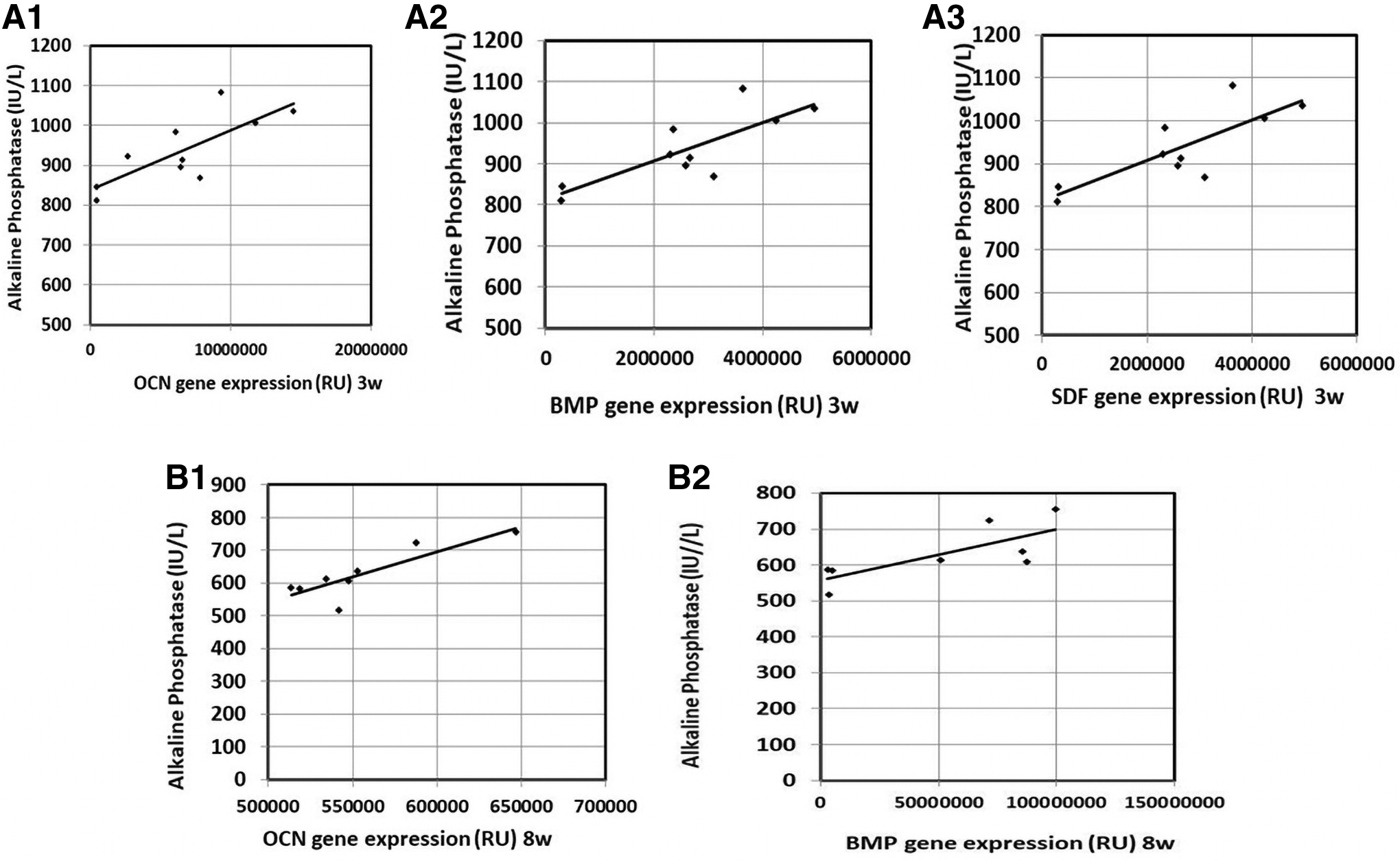
Correlation of serum ALP levels with gene expression of the studied parameters in the intra-peritoneal group:
Histopathology results
Three weeks postfracture, there were poorly organized fibrovascular tissue and few chondroid cells in groups I and II. More chondroid cells had appeared with the fibrovascular tissue in MSC groups (III and IV). Large areas of cartilage-like tissue with gradual maturation into bone were observed in group III. Eight weeks postfracture, new bone formation showed a similar trend to the new bone formation observed in 3-week sections. The highest amounts of new bone had formed in group III followed by group IV. This implies that MSCs initiated faster bone regeneration (Figs. 6 –8).
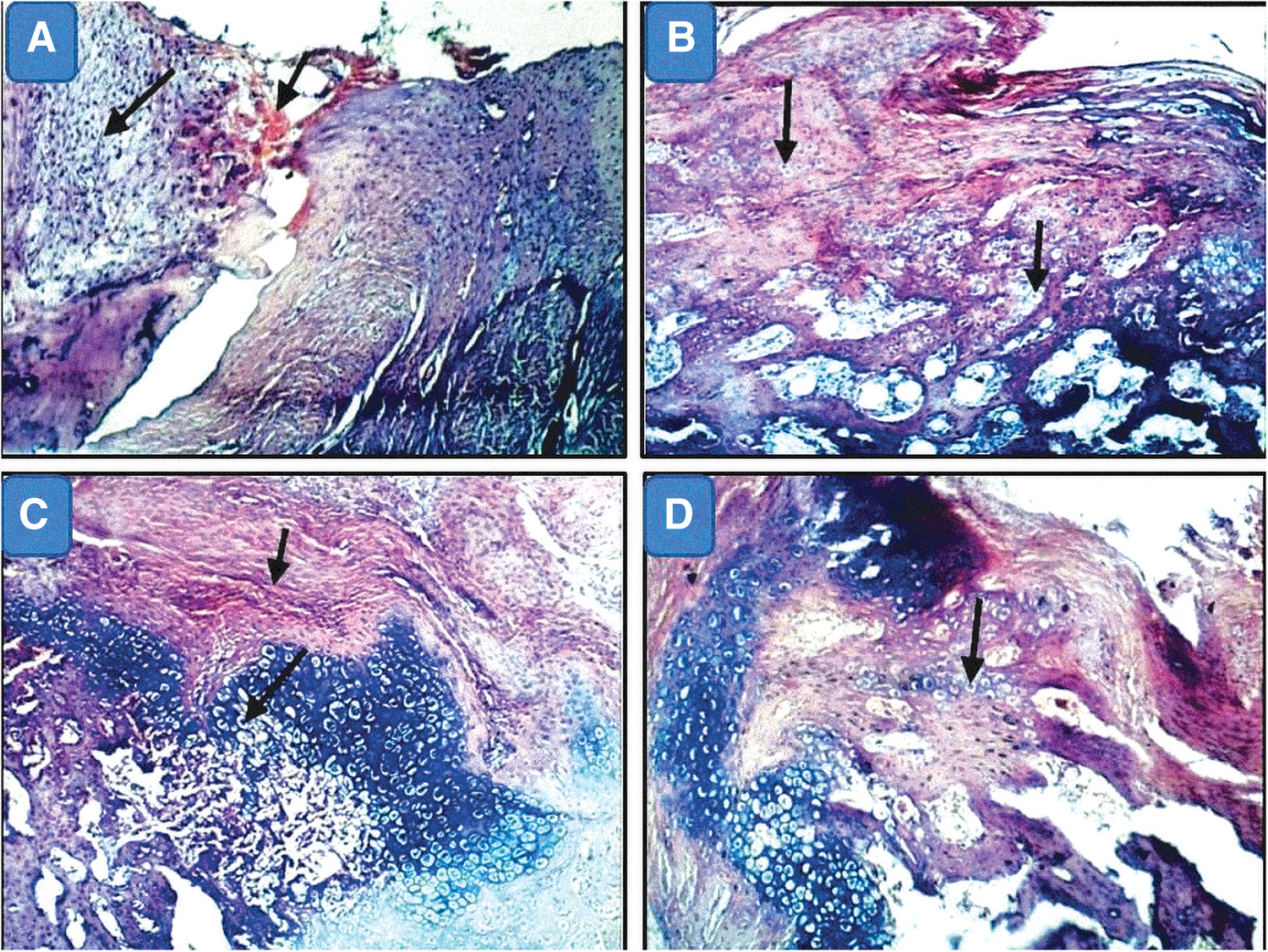
H&E-stained sections of femur specimens from studied groups.
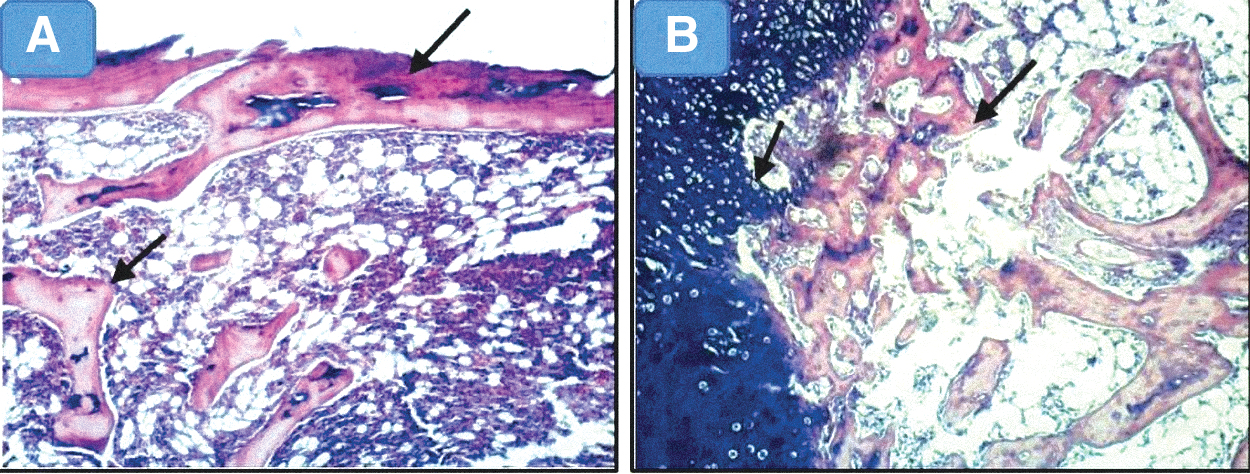
H&E-stained sections of femur specimens from studied groups.
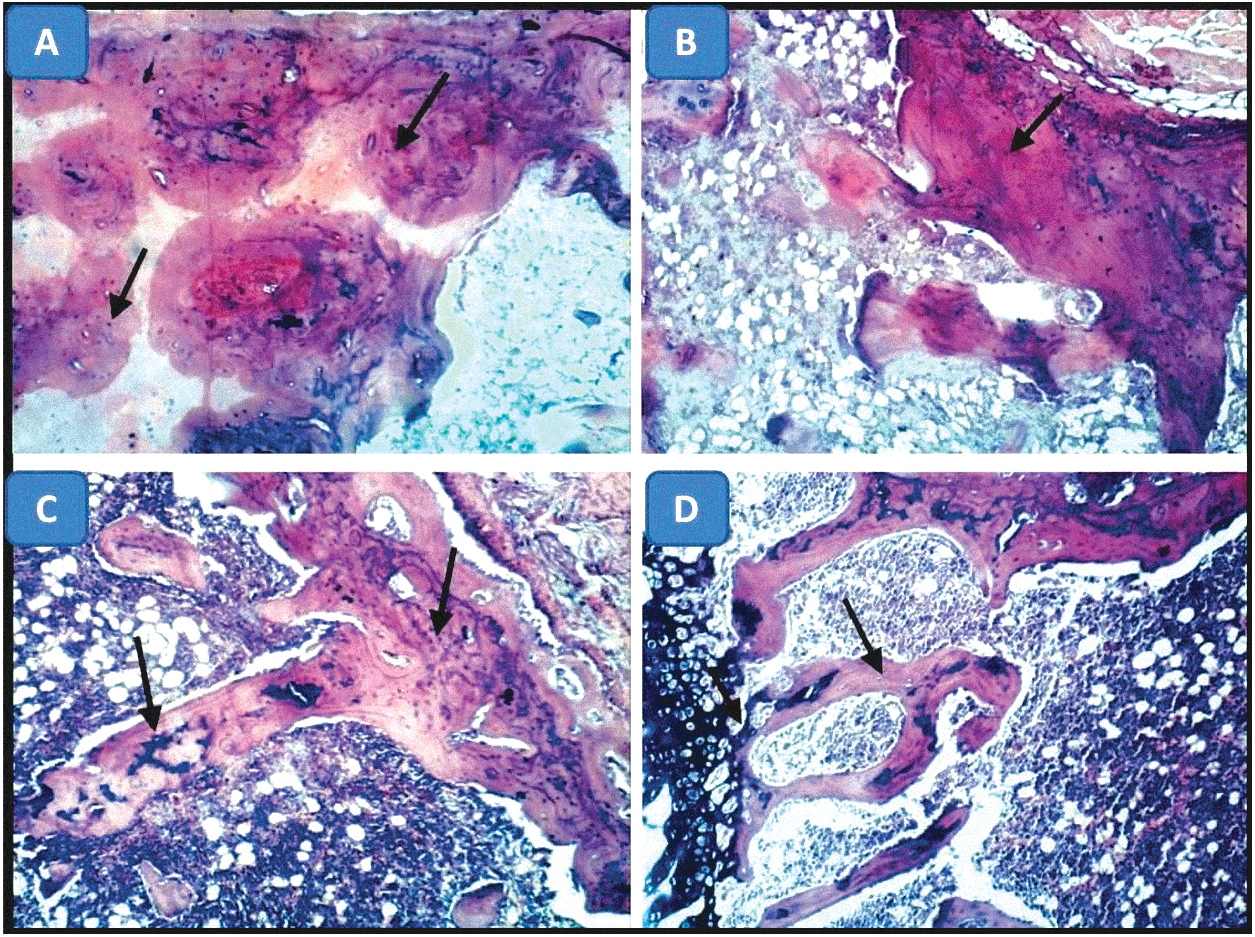
Photomicrographs of bone of rats 8 weeks after fracture (H&E × 100).
Evaluation of femur fracture in terms of callus formation and fracture lines visibility in the X-ray images showed superior fracture healing in group III (IL-MSCs) at 8 weeks (Fig. 9).
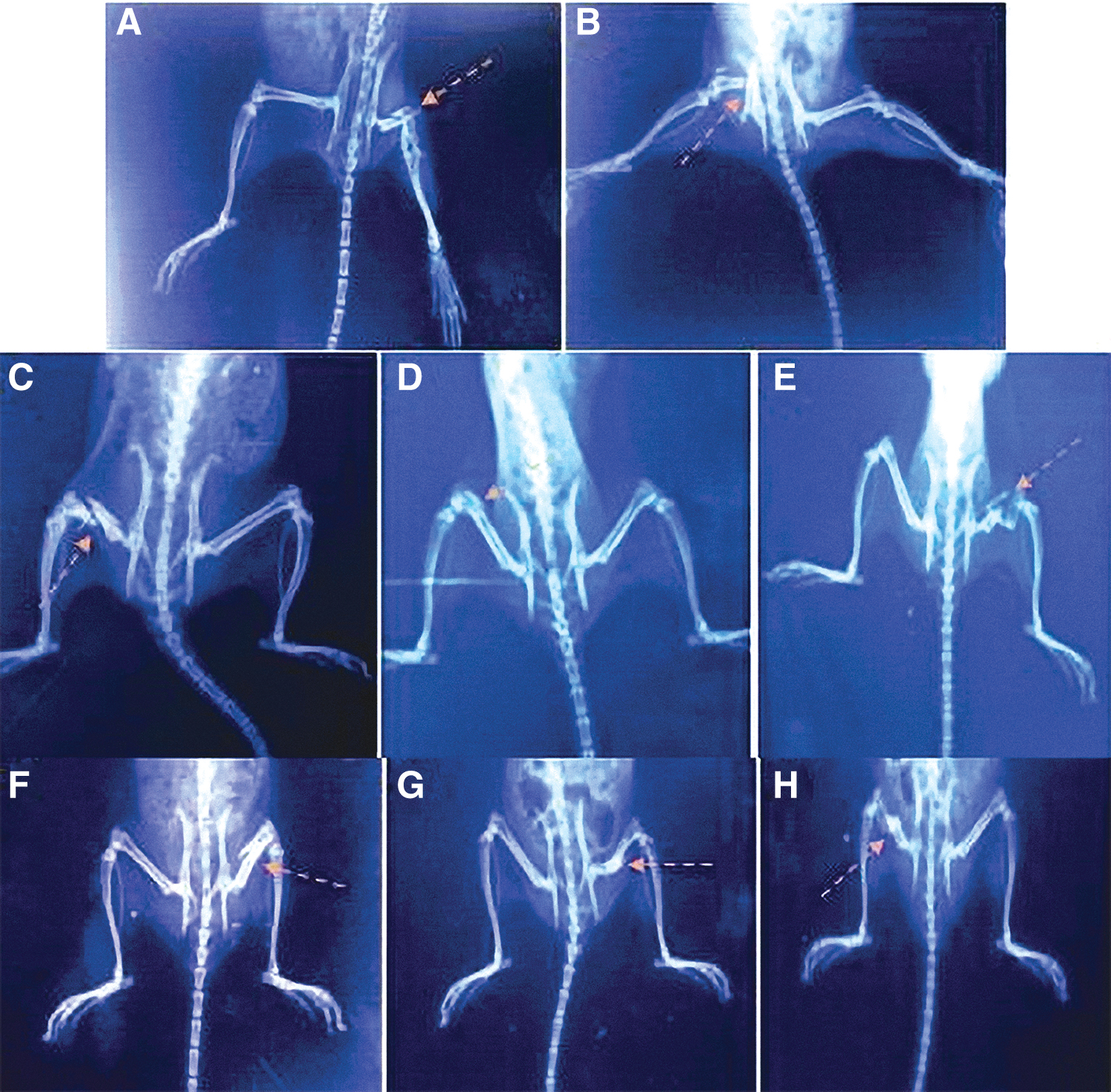
X-ray radiographs of fractured femurs in the studied groups.
Discussion
Small bone damage can repair spontaneously. Extensive damage needs reconstructive surgery and bone transplantation, which is a difficult line of treatment and faces donor problem or risk of autologous bone graft surgery. MSCs are multipotent cells capable of self-renewal and differentiation into several cell types forming new tissues under the influence of micro-environmental factors. So, they are attractive therapeutic candidates (Khakoo and others 2006).
This study aimed to evaluate the impact of MSCs on bone fracture of the femur shaft in mice through IL and IP routes and evaluation of stage biochemical mediators of healing. This study was performed on 4 groups (10 mice each with fixed fractures); group I: fixation, group II: received PBS (fixation+PBS), group III: received IL-MSCs (fixation+IL-MSCs), and group IV: received IP-MSCs (fixation+IP-MSCs).
This study was performed on mice as animal models because of the physiologic and genetic similarities between rodents and humans and the breeding capacity (Garcia and others 2013).
In this study, fluorescent-labeled MSCs were transplanted to mice in MSC-recipient groups (III and IV) to allow detection of the incorporated MSCs into the recipient bone tissue. There was a high number of fluorescent cells detected in IL group.
By histopathological examination of fractured mice femurs in the studied groups 3 and 8 weeks postfracture, new bone had been formed in MSC groups (III and IV). This result agreed with that of Zou and others (2016) who reported stable bone union in BM-MSCs, whereas a more fibrous union of fracture occurred in the control group. Radiological finding by X-ray supported the histological results. Previous studies have suggested that transplanted BM-MSCs induced bone regeneration by self-renewal and differentiation and enhancing the migration of host stem cells into injury sites in an endocrine manner (Khosla and others 2010 and Kumagai and others 2008).
Concerning ALP, total ALP rose to its peak value 3 weeks postfracture and then gradually decreases until 8 weeks. The IL-MSCs group was the only group that reached ALP baseline level. The MSC-recipient groups were the only groups that showed nonsignificant difference in OCN level compared with baseline level, as well. This could be explained by the fact that both ALP and OCN are products of osteoblasts and represent good markers for osteogenic maturation (Granéli and others 2014).
Measuring ALP and OCN at least 3 times (baseline preoperative, 3 weeks and 6 or 8 weeks postsurgery) in association with radiological findings was useful for monitoring the bone union process. Their usual pattern may help judge bone union (Nakagawa and others 2006). Based on the previous data, it became obvious that biochemical monitoring of the fracture-healing process would ideally allow early prediction of normal or delayed union, thus providing a basis for optimal management decisions.
Regarding OCN level, high OCN level indicated better bone formation, which was already confirmed histopathologically and biochemically in MSC-recipient groups and this reinforces the previous suggestion that MSCs would be useful for bone regeneration. Hosogane and others (2010) showed that stimulation with BMP-2 significantly increased ALP activity and OCN synthesis over control cells maintained in culture medium. In addition, BMP-2 induced osteogenic differentiation of multipotential mesenchymal cells into osteoblasts (Kim and others 2012). So, BMP-2 may be the control point of overall process as its level showed a significant decrease in 8 weeks than 3 weeks in IL-MSC group that showed accelerated healing. On the contrary, van Baardewijk and others (2013) reported that BMP-2 were not significantly associated with fracture-healing time.
This study found that SDF-1 gene expression was significantly increased in all groups 1 day and 3 weeks postfracture, and this result agreed with Ma and others (2005) and Togel and others (2005) who demonstrated an increase in SDF-1 expression within 24 h postinjury. Increased expression of SDF-1 mRNA was observed on days 2 and 3 in another study conducted by Kitaori and others (2009). However, its level showed no significant difference between 1 day and 3 weeks postfracture. The discrepancy in the peak point of SDF-1 expression may be related to the type of injuries. Many of the reported animal models were vascular injury types, in which oxygen tension changes rapidly (Ma and others 2005; Togel and others 2005). It occurred also in our study as trauma was the major cause of vascular injury that occurred during induction of fracture. SDF-1 is regulated by a hypoxia-specific transcriptional factor (hypoxia-inducible factor 1). So, the expression of SDF-1 may increase rapidly after the blood supply is impaired in those models (Ceradini and others 2004).
In a study performed by Ho and others (2015), MSCs transfected with SDF-1 gene secreted greater SDF-1 protein leading to increased MSC migration and retention in the fracture site and mobilization of nontransfected cells into this site with enhanced fracture healing. Our data showed positive correlation between SDF-1 and BMP-2 in IL-MSC group. This finding may explain enhancement of healing in IL-MSC group. The role of MSCs in bone regeneration can be explained also by the large number of cells produced together with their pluripotency, which allows the availability of different cell types that promote regeneration, with their immune inhibitory effect making transplantation much safer (Qin and others 2014).
In our study IL-MSC group showed better results than IP-MSCs. This can be explained by previous results with local rather than systemic injection because most infused MSCs are trapped in the lungs; only few migrate to the site of injury (Schrepfer and others 2007).
In addition, SDF-1 could also enhance the early osteogenic differentiation of stem cells, mediated by a BMP signaling pathway. A study by Hosogane and others (2010) showed that blocking of the SDF-1/C-X-C motif chemokine receptor (CXCR4) signal axis or adding SDF-1 protein to MSCs significantly affected BMP-2-induced ALP activity and OCN synthesis. Bone nodule mineralization decreased as well when the SDF-1 signaling was disrupted. They also showed that blocking the SDF-1 signaling inhibited RUNX2 and osterix expression, 2 important regulators of osteogenesis.
A study by Zhu and others (2011) in mice reported a significant impaired osteoblast development and bone formation when CXCR4 was inactivated in osteoprecursor cells, concluding that CXCR4 signaling is necessary for normal bone formation. Moreover, the osteogenic differentiation of mesenchymal and marrow stromal cells in response to BMP-2 was suppressed when CXCL12/CXCR4 signaling was inhibited (Zhu and others 2011). So, we used early passage of MSCs as late cell passage loses CXCR4. From previous studies, we can explain the positive correlation between serum ALP and SDF-1, OCN, and BMP-2.
Granero-Molto and others (2009) reported that mice transplanted with IL-MSCs displayed significant increase in total volume, total bone, soft tissue, new bone and callus volumes and mineralization compared with controls. MSC transplant improved the fracture healing by increasing the material toughness of the callus to be less brittle. Transplanted MSCs had systemic anti-inflammatory effects on cytokines released postfracture, significantly reducing IL-6 levels at day 3 and tumor necrosis factor α and IL-1β levels at 1 and 3 days postfracture. This process may limit tissue injury and prevent fibrosis development to promote rapid regeneration.
The effectiveness of the direct MSC injection was supported by Hernigou and Beaujean (2002), who found a positive correlation between the quantity of BM aspirate concentrate and mineralized bone formation to treat nonunion of the tibia and reported good results in 60 patients.
In conclusion, administration of MSCs either local or systemic improves and accelerates healing of bone fracture with better results in local injection as shown by radiological and histological findings with return of biochemical parameters to their normal level at 8 weeks in IL-MSC-injected groups. The high number of MSCs in IL-injected group that consequently affected the expression of biochemical mediators is the main cause of difference of IL and IP-injected groups.
Footnotes
Authors' Contributions
M.A.E., S.H.A. designed and directed the project. O.A., S.A.M., O.Y.H., E.M.A. and S.H. performed the experiments, analyzed the data and wrote the article that was revised by all co-authors.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Research Involving Animals
All animals were cared in accordance with the guidelines for animal research issued by the National Institute of Health, which was approved by Animal Ethics Committee, Zagazig University.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1