
Abstract
Infection is the leading cause of admission and mortality in neonatal intensive care units. Immature immune function and antibiotic resistance make the treatment more difficult. However, there is no effective prevention for it. Recently, more and more researches are focusing on stem cell therapy, especially mesenchymal stem cells (MSCs); their potential paracrine effect confer MSCs with a major advantage to treat the immune and inflammatory disorders associated with neonatal infection. In this review, we summarize the basal properties and preclinical evidence of MSCs and explore the potential mechanisms of paracrine factors of MSCs for neonatal infection.
Introduction
Newborns, especially preterm infants, are at significant risk for multiple infections during antenatal period, in parturition and hospitalization. Intrauterine infection is the main mechanism associated with preterm birth (Helmo and others 2018), which can increase the incidence of intraventricular hemorrhage, bronchopulmonary dysplasia (BPD), sepsis, and other complications (Thomas and Speer, 2011). With the increasing successful rescue of high-risk neonates, the incidence of preterm birth is on the rise.
Preterm infants in NICU are susceptible to hospital-acquired infection due to their immature immune systems, need for invasive procedures, and prolonged hospitalization. They also have higher mortality, and experience neurodevelopmental and growth impairment compared to uninfected counterparts (Mobley and others 2017). Currently, there is no specific prevention for neonatal infection, so new treatment is extremely urgent.
Mesenchymal stem cells (MSCs), mesodermal stem cells with self-renewing and multidirectional differentiation abilities, which can differentiate into osteocytes, adipocytes, and chondrocytes, and into nonmesodermal-derived cells under specific conditions (Kariminekoo and others 2016). In addition, MSCs can secrete many cytokines and proteins by paracrine signaling, and exhibit anti-infection, antioxidation, immune regulation, neuromodulation, hormone regulation, metabolic regulation, and other functions (Caplan, 2017).
The properties that make MSCs attractive therapy for treatment of acute disease as follows:
First, MSCs are relatively nonimmunogenic due to their low expression of major histocompatibility complex type I and type II, so they can be used as an allogeneic transplantation without the need for immunosuppression. Second, MSCs may decrease injury or restore function in diverse organs, including the heart, liver, lungs, and kidneys (Keane and others 2017). Finally, MSCs have low tumorigenicity (Park and others 2016) and rapid proliferation.
Initial research focused on the ability of MSCs to migrate and engraft at sites of tissue injury and exert local reparative effects (Francesco and others 2015). However, with technical improvement, a growing number of studies have shown that MSCs have the effects of anti-inflammatory, antimicrobial, antiapoptotic, and restoring multiple organs function through a robust pathway—paracrine signaling (Walter and others 2014). MSCs secrete a variety of cytokines and proteins, which exhibit biological effects on adjacent target organs, tissues, or cells, including various growth factors (such as vascular endothelial growth factor, VEGF), cytokines (such as interleukin), proteases, and some biological active substances (such as exosomes). This review will present the paracrine effects of MSCs that modulate important pathobiological pathways in life-threatening infections (Figs. 1 –4). A summary of referenced literature is included in Table 1.

Potential paracrine mechanism of MSCs in pulmonary infection. MSCs modify important pathobiological pathways in pulmonary infection through the release of paracrine factors, which include anti-infection, antimicrobial, reduction of the permeability of the alveolar epithelium, attenuation of impaired alveolarization, and angiogenesis. ANGPTL-1, angiopoietin-like protein 1; AT I cell, alveolar epithelial cells type I; AT II cell, alveolar epithelial cells type II; BD2, β-defensin-2; GM-CSF, granulocyte-macrophage colony-stimulating factor; KGF, keratinocyte growth factor; MSC, mesenchymal stem cell; PGE2, prostaglandin E2; RBC, red blood cell; VEGF, vascular endothelial growth factor. Color images are available online.
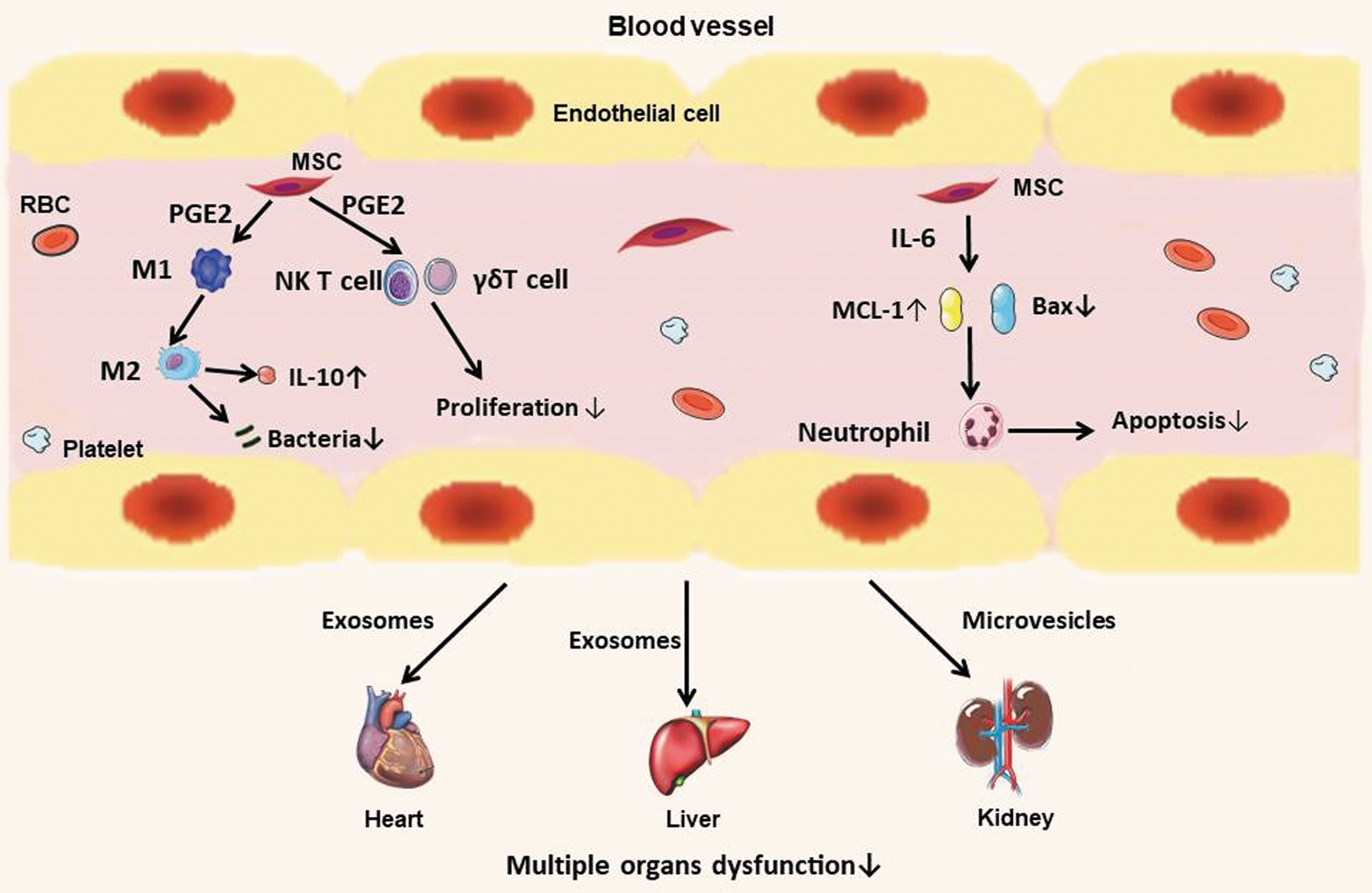
Potential paracrine mechanism of MSCs in systemic sepsis. MSCs exert immunomodulatory function by paracrine factors, control tissue inflammation and vascular integrity, increase microbe phagocytosis and clearance, and improve multiple organ dysfunction after sepsis. M1 and M2, macrophage; NK, natural killer. Color images are available online.
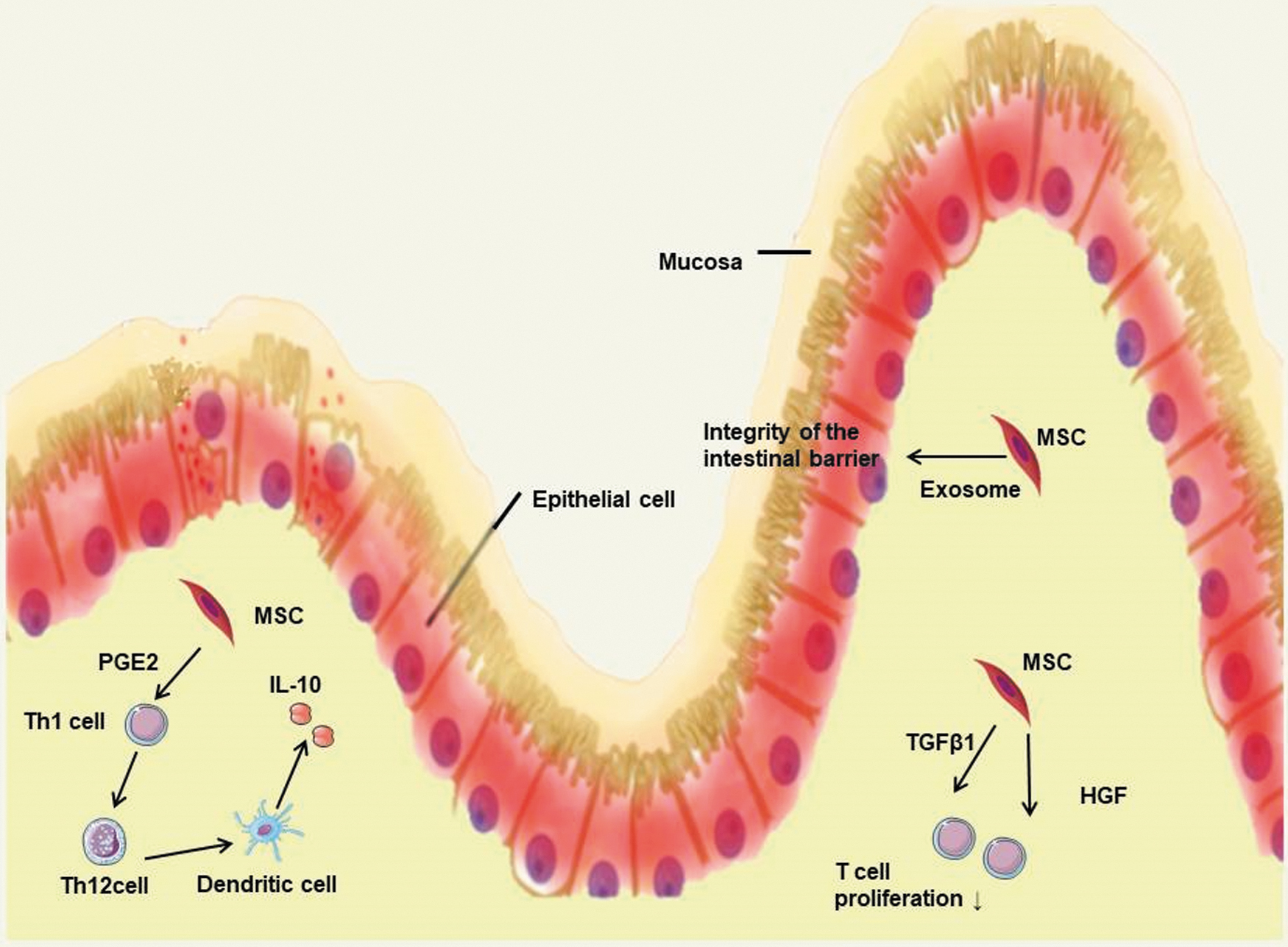
Potential paracrine mechanism of MSCs in intestinal infection. MSCs regulate intestinal immune and the integrity of the intestinal barrier by paracrine pathways. HGF, hepatocyte growth factor; TGF-β1, transforming growth factor-β1; Th, helper T cell. Color images are available online.

Potential paracrine mechanism of MSCs in central nervous system infection. MSCs suppress inflammatory injury and ameliorate the destruction of the blood–brain barrier by their paracrine factors, drive vascular anastomosis of injury lesions. TSG-6, tumor necrosis factor α-stimulated gene 6. Color images are available online.
Preclinical Studies Supporting the Paracrine Mechanisms of Mesenchymal Stem Cells for Infection
Ad, adipose derived; ALI, acute lung injury; AKI, acute kidney injury; ATII, alveolar epithelial cells type II; CLP, cecal ligation and puncture; hA-MSC, human allogeneic mesenchymal stem cells; hBM, human bone marrow; hUCB, human umbilical cord blood; HSCs, hematopoietic stem cells; hUC-MSC, human umbilical cord-derived MSC; IP, intraperitoneal; IRPA, imipenem-resistant Pseudomonas aeruginosa; IT, intratracheal; IV, intravenous; LPS, lipopolysaccharide; MBM, murine bone marrow; MCAO, intraluminal middle cerebral artery occlusion; NEC, necrotizing enterocolitis; NFI, neuroinflammatory injury; PGE2, prostaglandin E2; SBM, swine bone marrow; TGF-β, transforming growth factor-β; TSG-6, tumor necrosis factor α-stimulated gene 6.
Paracrine Mechanism of MSCs in Pulmonary Infection
Acute lung injury (ALI) induced by bacterial pneumonia is an important cause of morbidity and mortality in critically ill patients. Several preclinical studies have demonstrated the potential of MSCs to decrease injury after pulmonary infection by their paracrine signaling rather than cell differentiation. Research has shown that in Escherichia coli pneumonia model, Toll-like receptor-4, which is distributed on the surface of macrophages, activates MSCs to secrete β-defensin-2 (BD2), downmodulating the inflammatory responses and enhancing bacterial clearance (Sung and others 2016). In addition to the E. coli model, in a model of cystic fibrosis, lung infection induced by Pseudomonas aeruginosa and Staphylococcus aureus, MSCs can produce an antimicrobial peptide—LL-37, functions as directly killing microorganisms, regulating inflammation, angiogenesis, and antiapoptosis (Chieosilapatham and others 2018), which impacts the rate of bacterial growth and transition into colony-forming units in vitro, and finally impacts the MSC antimicrobial potency and efficacy (Sutton and others 2016).
Similarly, our group separated imipenem-resistant P. aeruginosa (IRPA) from neonates and incubated it with human umbilical cord mesenchymal stem cells (HUCMSCs); we found that the growth of IRPA was inhibited by MSC-secreted products—LL-37, BD2 (Ren and others 2020). So the above studies together confirmed the antibacterial properties of paracrine factors of MSCs.
As we all know, BPD is also an inflammatory process, and stem cell is a promising treatment for pulmonary injury caused by infection. Recently, some studies have shown that hyperoxia-induced lung injury is alleviated with alveolar and vascular remodeling (Aslam and others 2009; Van Haaften and others 2009; Michael and others 2018). Moreover, it is also shown that the paracrine actions of MSCs are effective in BPD treatment (Hansmann and others 2012; Fung and Thebaud, 2014).
In a model of hyperoxia-induced lung injury (Chang and others 2014), intratracheal transplantation of HUCMSCs with or without VEGF knockdown, it could be found that VEGF knockdown abolished the protective effects of MSCs in hyperoxic lung injury, such as the attenuation of impaired alveolarization and angiogenesis, and downregulation of proinflammatory cytokine levels; at the same time, the increased expression of inflammatory cytokines was attenuated in cells that were co-cultured with nontransfected or scrambled siRNA-transfected MSCs in vitro, VEGF levels also increased, and oxidative stress-induced cell death was significantly lower. So VEGF secreted by MSCs is the key paracrine factor in protection against lung injury induced by hyperoxia, it may contribute to alveolar and vascular remodeling.
Acute respiratory distress syndrome (ARDS) is caused by direct lung injury or systemic inflammatory response with increased permeability of the capillary endothelium and alveolar epithelium. To study the effects of MSCs and its paracrine factors on the alveolar epithelial cell type II (ATII), a research had found that allogeneic human MSCs restored epithelial protein permeability after cytomix-induced injury, which was mediated by the paracrine soluble factor—angiopoietin-like protein 1 (ANGPTL-1), effected on cytoskeletal re-organization of both actin and claudin18, and suppression of NF-kB activity (Fang and others 2010). Therefore, paracrine mechanisms of MSCs will be a potential therapy for pulmonary injury induced by infection, and may be a prospect for ARDS and BPD prevention.
Besides bacteria, viral infection is also the most common cause of pulmonary injury. Coronavirus disease 2019 (COVID-19) is an acute respiratory infectious disease caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). The current COVID-19 epidemic has rapidly grown expanding to become a global major concern. Now, there is no specific therapy for the disease, and most treatments are limited to supportive care. The main pathologic features of severe COVID-19 were consistent with ALI/ARDS, characterized by cellular fibromyxoid exudates, extensive pulmonary inflammation, pulmonary edema, and hyaline membrane formation (Rubenfeld and others 2005).
Stem cell therapy has been proved to be a potential treatment for refractory pulmonary illnesses, and has shown some valuable results in preclinical studies. MSCs have been identified to efficiently cure ALI/ARDS, mediated mainly by paracrine mechanisms based on the released extracellular vesicles (EVs), like microvesicles and exosomes (Fujita and others 2018). MSC exosomes can promote the regeneration of alveolar epithelial cells, exert antialveolar inflammation, prevent endothelial cell apoptosis, inhibit early epithelial-mesenchymal transdifferentiation, and prevent myofibroblast growth, thereby alleviating pulmonary fibrosis (Wang and others 2013).
In addition, MSCs can also release prostaglandin E2 (PGE2), granulocyte-macrophage colony-stimulating factor, keratinocyte growth factor, IL-6, and IL-13 to promote the phagocytosis and alternative activation of alveolar macrophage, alter the cytokine secretion profile of dendritic cell subsets, and decrease the release of interferon γ from natural killer cells (Liu and others 2020), which might be effective on pulmonary injury caused by COVID-19.
As of yet, MSC therapy has shown an improvement in the treatment of COVID-19, reducing the cytokine storm, enhancing alveolar fluid clearance, and promoting epithelial and endothelial recovery (Sara and Abdelalim, 2020). A cohort study addressed the safety and efficacy of exosomes derived from allogeneic bone marrow MSCs as treatment for severe COVID-19; exosomes have capacity to restore oxygenation, downregulate cytokine storm, and reconstitute immunity, and are a potential therapeutic candidate for severe COVID-19 (Sengupta and others 2020). Therefore, MSCs have been proposed as an emerging therapeutic option for the COVID-19 infection.
Paracrine Mechanism of MSCs in Systemic Sepsis
MSCs demonstrated the efficacy in several preclinical systemic sepsis models; the paracrine effects of MSCs play an important role in sepsis (Baudry and others 2019). Prockop and his team found that MSCs secreted an arachidonic acid metabolite—PGE2 in a model of cecal ligation and puncture-induced sepsis, which promoted the conversion of macrophages to the anti-inflammatory phenotype (M2), increased the secretion of IL-10, and reduced the pathogen load of inflammatory tissues and organs (Prockop, 2013). Besides the above mechanism, the immunomodulatory function of MSCs is also important in sepsis. Recently, Prigione and others (2009). discovered the inhibitory effect of MSCs on the proliferation of invariant natural killer T cells and γδT cell in the peripheral blood, which was mediated by secreting PGE2, increasing the possibility in MSC-based therapy of autoimmune diseases.
Neutrophils are the first cells arriving to the inflammatory tissue; MSCs may show its immunomodulatory effects by interacting with neutrophils. A foreign study had shown that after co-cultivation of bone marrow-derived MSCs with neutrophils, MSCs could inhibit neutrophil apoptosis, and when co-cultivated with neutrophils stimulated by inflammation, the antiapoptotic effects still existed (Rafffaghello and others 2008). It was further proven by Transwell experiment and antibody blocking experiment that this antiapoptotic effect was mediated by IL-6, a soluble factor secreted by MSCs, thus causing Bax (proapoptotic protein) downregulation and MCL-1 (antiapoptotic protein) upregulation.
Sepsis induces multiple organ dysfunction due to overexpression of the inflammatory response, increasing the associated morbidity and mortality (Capcha and others 2020). There was a study that showed MSC-derived exosomes, an EV subtype (40–100 nm), which contain various components such as proteins, nucleic acids, and lipids derived from parental cells, have similar functions with MSCs such as repairing tissue damage, suppressing inflammatory responses, and regulating the immune system (Zhang and others 2019), which played a role in sepsis caused by a variety of bacteria; by transferring antiapoptotic miRNA-223 to cardiomyocytes and macrophages, the expression of the proinflammatory factors Sema3A and Stat3 was reduced, and thereby alleviated inflammatory response and reduced myocardial cell death (Wang and others 2015).
In addition to improving heart function, human umbilical cord blood (hUCB)-MSC-derived exosomes induced prosurvival effects in ischemia/reperfusion-induced hepatic dysfunction in mice, hUCB-MSCs released exosomes that transferred miR-1246 to hepatocytes and subsequently produced protective effect by regulating Th17/Treg imbalance, which was mediated through the IL-6-gp130-STAT3 pathway (Xie and others 2019). Another bioactive substance secreted by MSCs—microvesicles (50–1,000 nm), which played an important role in improving acute kidney injury induced by cisplatin, promoted the expression of antiapoptotic genes such as Bcl-xL, Bcl2 and BIRC8”,downregulated proapoptotic genes such as Casp1, Casp8 and LTA, and enhanced functional recovery (Bruno and others 2012). Therefore, these paracrine factors provide a potential effect in multiple organ dysfunction caused by sepsis.
Paracrine Mechanism of MSCs in Intestinal Infection
Preterm infants have an immature digestive system and are prone to various gastrointestinal emergencies, such as necrotizing enterocolitis (NEC). Some preclinical researches have proved the potential of MSCs in inflammatory bowel disease (IBD). MSC-derived PGE2, transforming growth factor-β1 (TGF-β1), and hepatocyte growth factor (HGF) can attribute to the therapy of IBD by their immunomodulatory effects (Mao and others 2017). PGE2 promoted T cell polarization from Th1 to Th2 subtype, inducing dendritic cells to produce more IL-10. TGF-β1 and HGF were responsible for the MSC-mediated reduction in T cell proliferation. All of these paracrine mechanisms have potential benefits in intestinal infection.
Furthermore, Rager and others (2016) found that intraperitoneal injection of bone marrow MSC-derived exosomes reduced the incidence and severity of NEC and maintained the integrity of the intestinal barrier, which might be caused by the antiapoptotic miRNAs (such as miR-15b and miR-34c), and growth factor receptors in exosomes interacted with mRNAs associated with wound healing (Yu and others 2015). Amniotic fluid-derived exosomes, neural stem cell-derived exosomes, and neonatal intestinal neural stem cell-derived exosomes can also reduce the incidence and severity of NEC as effectively as the stem cells from which they are derived, supporting the potential of exosomes for NEC (Mcculloh and others 2018).
Paracrine Mechanism of MSCs in Central Nervous System Infection
MSCs have the capacity to migrate to the injury regions, exerting tissue regeneration and inflammatory modulation. It has been reported that MSCs can upregulate the expression of brain-derived neurotrophic factor and some neuroprotective factors, and can improve neurological recovery in many central nervous system diseases (Martino and others 2010; Song and others 2015). In the neuroinflammatory injury mice model caused by Japanese encephalitis virus (Bian and others 2017), MSCs significantly suppressed inflammatory lesions by expression of the cytokines TGF-β and tumor necrosis factor α-stimulated gene 6 (TSG-6), inhibited the destruction of the blood–brain barrier, and ameliorated tissue injury.
Studies abroad have found that some meningitis can present with small vessel vasculitis causing infarct (Mishra and others 2018). In cerebral infarct of rats, VEGF expression increased after tail vein injection of rat-derived bone marrow stromal cells, enhanced the proliferation of vascular endothelial cells, which subsequently derived vascular anastomosis, and benefited to regeneration in the infarction area and neurological recovery (Zhihua and others 2015).
Limitations and Challenges of MSCs
Despite the enormous progress in the therapeutic potential of MSC therapy for infectious diseases in the preterm infant, it was still hampered by a huge gap of knowledge, which include the following: (1) the optimal timing, dose, and route of administration and (2) the donor age, source, culture methods, stem cell type, quality control, and stem cell preconditioning. MSCs can be obtained from neonatal tissues (umbilical cord and placenta) and various adult (peripheral blood, bone marrow, and adipose tissue) sources. However, isolation of MSCs from adult sources involves invasive and painful procedures with possible donor site morbidity (Hass and others 2011), and the amounts of MSCs obtained by these isolations vary enormously. MSCs obtained from neonatal tissues avoid invasive procedures and ethical problems, and have a higher expansion and engraftment capacity (Barlow and others 2008). A study has shown that MSCs from the umbilical cord blood constitute the therapeutic option closest to clinical use, and suggests that the early preventive application is superior to late administration to the injured lung and brain (Gronbach and others 2018).
One of key challenges is the difficulty in sufficient quality control and assurance of MSCs that were derived from different donors with variance of batch-to-batch. Recently, functional MSCs from human induced pluripotent stem cells (iPSCs) have been successfully differentiated, which overcome many disadvantages of batch-to-batch variations in MSC quality, stem cell senescence, and limited proliferative potency. iPSC-MSCs have superior survival and engraftment following transplantation to induce vascular and muscle regeneration than bone marrow MSCs (Lian and others 2010), and display higher proliferative potential and stronger immunomodulation (Zhang and others 2012).
Therefore, iPSC-MSCs may provide another putative cellular source to overcome many limitations of adult MSCs. In a phase 1, open-label clinical trial (Bloor and others 2020), CYP-001 (iPSC-derived MSCs) has been used in refractory graft-versus-host-disease and proved the safety and efficacy of iPSC-derived MSCs; the therapeutic prospects of iPSC-derived MSCs would be explored in diverse inflammatory and immune-mediated diseases.
The route of MSC transplantation is an important consideration. We have proved the safety of autologous cord blood cell infusion in preterm neonates, and found it could reduce respiratory support duration and potentially preterm complications (Zhuxiao and others 2020). Yun Sil and his team demonstrated that local intratracheal MSC transplantation is more effective than systemic intraperitoneal administration in protecting against hyperoxic lung injury (Chang and others 2009), and had a lower burden of unexpected side effects compared with intravenous administration.
It is the principle of MSC therapy to infuse the smallest dose of stem cells, but obtain the greatest efficacy. Various studies have explored the dose selection of cord blood mononuclear cell infusion, and the results show that mononuclear cell infusion of 1–5 × 107/kg is safe and feasible, and there are no adverse reactions and liver toxicity (Cotten and others 2014; Kourembanas, 2014; Rosenkranz and others 2012). On this basis, some studies have found that 1 × 107/kg mononuclear cell infusion has the lowest therapeutic effect, while 5 × 107/kg mononuclear cell infusion has the greatest efficacy (De and others 2012; Peng and others 2019).
Furthermore, MSC paracrine function is regulated by telomerase-associated protein/inflammatory pathways, including RAP1/NF-kB signaling pathway. The NF-kB transcription factor family provides a mechanism for cells to respond to a variety of inflammation stimuli (Bruno and others 2018), and inhibition of NF-kB activation has an anti-inflammatory effect. Rap1, a telomeric repeat-binding factor 2-interacting protein 1, is a novel modulator and known to regulate inflammation through NF-kB signaling (Teo and others 2010). Some studies find that the absence of Rap1 reduces the efficacy of MSC paracrine by inhibiting the NF-kB signal pathway, resulting i the decrease of cytokines like IL-6, IL-10, and IFN-γ, which decrease the immunosuppressive potency of MSCs (Ding and others 2018).
MSCs are also vulnerable to ischemic or hypoxic environment and undergo stem cell senesces post-transplantation. Liang and others (2015) recently revealed that introducing ERBB4 into MSCs ameliorated MSC survival potential by effective homing and activation of the PI3K/Akt pathway.
Recent studies showed MSC-mediated mitochondrial transfer is another important mechanism potentiating MSC tissue repair and immunomodulation. Mitochondrial dysfunction is common in many diseases. High efficiency of mitochondrial transfer from MSCs to tissue cells may be beneficial for alleviation of injury. There is a study that found human iPSC-MSC transplantation reduced T helper 2 cytokines, attenuated the mitochondrial dysfunction of epithelial cells, and relieved asthma inflammation, mitochondrial transfer from iPSC-MSCs to epithelial cells by tunneling nanotubes (TNTs) regulated by connexin 43 (Yao and others 2018). In addition, MSC functional mitochondria were also effectively transferred to cardiomyocytes (Zhang and others 2016), eye cells (Jiang and others 2016), and islet cells (Rackham and others 2020).
Recent series of evidence indicates that MSCs can communicate with their microenvironment through bidirectional exchanges of mitochondria. Mahrouf-Yorgov and others (2017) used a co-culture system consisting of MSCs and distressed somatic cells and found that foreign somatic-derived mitochondria were engulfed and degraded by MSCs, leading to induction of the cytoprotective enzyme heme oxygenase-1 (HO-1) and stimulation of mitochondrial biogenesis, finally promoting the MSCs to donate their mitochondria to injured cells to combat injury. Similarly, inflammatory cytokine TNF-α produced by MSCs can stimulate the formation of TNTs between MSCs and cardiomyocytes regulated through the TNF-a/NF-kB/TNFaIP2 signaling pathway (Zhang and others 2016).
Conclusion and Future Perspectives
The preclinical studies of MSCs have demonstrated the therapeutic potential for infection, which has been described as likely to change the practice of medicine (Caplan and Correa, 2011). Although several clinical trials have been recently registered to examine the safety and efficacy of MSCs as a therapeutic option for some diseases, fewer studies have achieved significant results. The most effective delivery route for MSCs with minimum toxic effects remains to be explored. Elucidating the molecular mechanisms of MSCs during organs injury is crucial to understand their role in infection. A series of experimental studies and trials will further our understanding of how MSCs interact with host tissue.
Nowadays, MSC therapy has been widely used in many clinical diseases, in which its paracrine factors play an important role. The mechanisms of paracrine signaling are increasingly well characterized, including modulation of the immune response and reduction of host injury by augmenting bacterial clearance and tissue repairment. Therefore, many researchers suggest that future studies should focus on developing genetically modified MSCs, generating large numbers of EVs that could safely transfer different effective therapeutic factors (Park and others 2019), which would help us to fully harness the therapeutic promise of MSCs for our patients with infection.
Footnotes
Authors' Contributions
J.Y. made substantial contributions to conception and design, J.M. wrote the text, and Z.R., Z.Z., X.X., J.W., and L.Y. all revised the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (81873847), Guangdong Nature Science Foundation (2021A1515012121), and Guang Zhou Science and Technology Grant under Grant (202102080104).