
Abstract
Cortisol is secreted in prolonged stress and has therapeutic effects in inflammatory diseases. Considering the immunomodulatory effects of mesenchymal stem cells, here we investigated the effect of hydrocortisone (HC) long-term treatment on immunomodulatory properties of human adipose-derived mesenchymal stromal/stem cells (ASCs). Isolated ASCs from healthy subjects were treated with different HC concentrations for 14 days. The effect of HC-treated ASCs on the proliferative response of peripheral blood mononuclear cells (PBMCs) was evaluated in ASCs/2-way mixed leukocyte reaction coculture using 2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide (XTT)-assay. HC-treated ASCs were further divided into interferon gamma (IFN-γ) stimulated and unstimulated groups. Transforming growth factor beta 1 (TGF-β1) and interleukin (IL)-6 levels were measured in culture supernatants by enzyme-linked immunosorbent assay. Relative expression of cyclooxygenase-2 (COX-2), hepatocyte growth factor, indoleamine dioxygenase, and programmed death-ligand 1 genes was assessed by real-time PCR. Levels of TGF-β1 and COX-2 expression were elevated in unstimulated ASCs, while exposure to high concentration of HC significantly increased TGF-β1 levels and reduced COX-2 expression. Unstimulated HC-5-μM-treated ASCs increased PBMC proliferation ratio on day 2 of coculture compared to the control group (P = 0.05). In IFN-γ stimulated condition, pretreatment with HC-5 μM resulted in a significantly increased IL-6 and significantly decreased COX-2 expression compared to the HC untreated control group. In conclusion, our results showed various alterations of ASC immunomodulatory related features as a result of long-term exposure of different concentrations of HC. It seems that HC at low concentration pushed the balance toward extended immune response in ASCs, while this observation wasn't persistent in ASCs treated with higher concentrations of HC.
Introduction
Cortisol is a major glucocorticoid hormone released by hypothalamus-pituitary-adrenal axis, which is produced by adrenal cortex in response to hypoxia and physiological and metabolic stresses (Koutmani and Karalis 2015). This hormone is involved in the maintenance of physiological functions of the thyroid gland, gastrointestinal tract, immune system, as well as metabolism of carbohydrates, lipids, and proteins (Tsigos and others 2016). Cortisol's roles in maintenance of tissue homeostasis are mediated by its engagement with glucocorticoid receptor (GR) or mineralocorticoid receptor, both of which are present in various tissues (Lee and others 2014; Jia and others 2017).
Endogenous or pharmacologic excessive level of cortisol impairs lipid and glucose homeostasis, leading to lipodystrophy and insulin resistance (Dalle and others 2019). Excessive levels of cortisol may affect the immune system function, so that if stress precedes inflammation, cortisol can boost production of pro-inflammatory cytokines and enhance immune responses, whereas in the presence of low amount of cortisol, production of pro-inflammatory cytokines can be reduced (Sorrells and others 2009).
Mesenchymal stem cells (MSCs) are multipotent stromal cells with self-renewal ability and multilineage differentiation capacity. These cells can be isolated from different tissues, including bone marrow, umbilical cord, and adipose tissue (Zhou and others 2011). MSCs have immunomodulatory properties and adopt different phenotypes under the influence of relevant microenvironment (Le Blanc and Davies 2015). For example, MSCs suppress immune cell function in inflammatory conditions with the help of anti-inflammatory factors such as transforming growth factor beta 1 (TGF-β1), indoleamine dioxygenase (IDO), human leukocyte antigen-G5 (HLA-G5), prostaglandin E2 (PGE2), and interleukin-10 (IL-10) (Kim and others 2018). In contrast, these cells have immune-enhancing properties, such as enhancement of neutrophils and monocyte migration toward infection site and preserving M1 macrophages against local infections through soluble factors such as IL-6, IL-8, and interferon beta (IFN-β) (Le Blanc and Davies 2015).
Glucocorticoids can affect MSCs in various ways; for instance, high amount of dexamethasone reduces the immunosuppressive function of MSCs (Ankrum and others 2014), while at low amount, dexamethasone decreases differentiation of MSCs and increases their proliferation (Xiao and others 2010). Budesonide increases IDO expression in IFN-γ stimulated MSCs (Ankrum and others 2014), and prednisone enhances the immunosuppressive properties of MSCs by increasing expression and activity of IDO and cyclooxygenase-2 (COX-2) in IFN-γ stimulated MSCs (Javorkova and others 2018). In addition, concentration of hydrocortisone (HC) seems to be playing a major role in its treatment outcome, in which short-term exposure to high concentration of HC can boost bone marrow MSCs' immunosuppressive features, while low concentration of it doesn't show the similar effect (Durand and others 2019).
Prolonged presence of cortisol in chronic stresses, as well as administration of glucocorticoids in a number of diseases such as chronic obstructive pulmonary disease (COPD), rheumatoid arthritis, and systemic lupus erythematosus (SLE), can affect immunological events like inflammation (Fan and Morand 2012). The majority of prior studies looked at the short-term effects of glucocorticoids on stem cells (Abdanipour and others 2015), and there is lack of data on glucocorticoids' long-term effects on these cells. In contrast, glucocorticoids' long-term effects on immune system mediators and responses have not been studied comprehensively.
Considering the various effects of different HC concentrations on the immune system (Dong and others 2018; Durand and others 2019), we aimed to investigate the long-term effect of HC at various concentrations on immunomodulatory properties of human adipose-derived mesenchymal stromal/stem cells (ASCs).
Materials and Methods
Isolation and culture of ASCs
After obtaining written informed consent, visceral adipose tissue samples were isolated from healthy individuals (5 female participants, age range: 25–40 years old) undergoing cosmetic surgery (IRB No. 1398.247).
Samples were transferred to the laboratory in Hanks basic salt solution buffer containing 300 U/mL penicillin, 300 μg/mL streptomycin, and 25 μg/mL amphotericin B. To isolate ASCs by a nonenzymatic method (Ghorbani and others 2014), the samples were cut into small pieces (∼5 mg) and placed in 6-well plates. The surface of each piece was covered with fetal bovine serum (FBS) (Life Technologies, United Kingdom), and the plate was incubated for 24 h at 37°C, 5% CO2, and 95% humidity. FBS was then replaced with low glucose Dulbecco's modified Eagle's medium (DMEM-LG) (Life Technologies) containing 100 μg/mL streptomycin, 100 IU/ml penicillin, and 15% FBS (complete culture media). Culture's media were changed every 2 days. Once fibroblast-like adhesive cells were observed fat fragments were removed.
Cells were then harvested using 0.25% trypsin/EDTA solution at 80% confluency and transferred into T25 flasks at 10,000 cells/cm2 density. Cultures were passaged for 4 times before further experiments.
Characterization of ASCs
Study of surface markers
Passage 4 of cultured cells was suspended in FACS buffer (PBS contains or with 0.1% BSA). The cells were incubated in a dark environment with CD105-FITC, CD45-FITC, CD34-PE, CD73-PE, and CD90-PerCP (BioLegend) (eBioscience) anti-human conjugated antibodies for 45 min at 4°C. Incubated cells with anti-mouse PE, FITC, and PerCP conjugated antibodies were used as isotype controls (eBioscience). Stained cells were washed 3 times with FACS buffer and fixed using 4% paraformaldehyde. Cell suspensions were evaluated for fluorescence intensity by flow cytometer (FACS Calibur; Becton Dickinson), and data were analyzed using FlowJo V.7.6 software (FLOWJO; LLC, OR).
Differentiation into adipogenic lineage
A suspension of 5,000 cells was seeded into a 24-well plate (SPL, Korea). After 24 h, the complete media were replaced with StemPro adipogenic differentiation medium (Life Technologies) and changed every 3 days. After 21 days, the cells were washed with PBS and fixed using 10% formalin. To assess differentiation into adipogenic lineage cells were stained with oil red O solution and examined for fat vacuoles using a light microscope (Olympus, Japan) at 200 × magnification.
Osteogenic differentiation
A suspension of 5,000 cells was seeded in 24-well plate (SPL, Korea). After 24 h, the complete media were replaced with StemPro osteogenic differential culture medium (Life Technology). The media were changed every 3 days, and after 21 days, the cells were washed by PBS and fixed with 10% formalin. The cells were subsequently stained with alizarin red S solution and examined by a light microscope at 200 × magnification to observe calcium precipitates.
Chondrogenic differentiation
Suspension of 1.6 × 107 cells/mL was prepared, and 5 μL droplets of it were seeded in the center of a 96-well plate and incubated for 2 h. StemPro chondrogenic differentiation medium (Life Technology) was added to the wells, and the media were changed every 3 days. After 14 days, the cells were washed using PBS, fixed by 10% formalin, and stained with 1% solution of alcian blue. Presence of proteoglycans was investigated by a light microscope at 100 × magnification.
Study design and cell treatment
To evaluate the effect of long-term cortisol treatment on immunomodulatory properties of ASCs in all test groups, cells from passage 4 were cultured separately for 14 days using 5, 25, and 100 μM concentrations of HC (Jaber Ebne Hayyan Pharmaceutical Company, Iran) in complete media, presented as the HC-5 μM, HC-25 μM, and HC-100 μM study groups, respectively. During this period, the cortisol containing media were changed every 48 h, and cells were passaged when needed. In addition, untreated ASCs were considered as control.
Due to the role of IFN-γ in MSCs priming and induction of their immunomodulatory properties (Ankrum and others 2014; Javorkova and others 2018), after 14 days of treatment ASCs in separate groups based on cortisol concentrations were stimulated by 100 ng/mL IFN-γ (Exir Pharmaceutical, Iran) for 24 h, and their immunomodulatory abilities and related features were also examined in this condition. Cortisol-free stimulated ASCs were considered as control. All the further experiments were performed in triplicate.
Investigation of gene expression
To measure the relative expression of immunomodulatory related genes, including COX-2, hepatocyte growth factor (HGF), IDO, and programmed death-ligand 1 (PD-L1) in cortisol-treated ASCs, total RNA was first extracted from stimulated and unstimulated groups, according to the kit protocol (SINACLON, Iran). cDNA synthesis was carried out using 1 μg of RNA based on manufacturer instruction (SINACLON, Iran). The expression of mentioned genes was investigated by real-time PCR using specific primers (Table 1) and SYBR green master mix (SINACLON, Iran) in Rotor-Gene™ 6000 sequence detection system (Corbett Life Science, Australia). The expression of target genes was calculated and reported relative to reference gene (β-Actin) using standard curves and 2−ΔCt formula (Schmittgen and Livak 2008). ΔCt was assessed as below:
Sequences, Product Size, and Annealing Temperatures of the Primers Designed to Assess the Expression of β-Actin, IDO, HGF, COX-2, and PD-L1 Genes Using Relative Real Time PCR
ASCs, human adipose-derived mesenchymal stem cells; COX-2, cyclooxygenase-2; IDO, indoleamine dioxygenase; PD-L1, programmed death-ligand 1.
Evaluation of cytokine levels
Supernatants from cultures of all study groups were collected after desired treatment duration and stored at −70°C. TGF-β1 and IL-6 levels were evaluated by enzyme-linked immunosorbent assay (ELISA) according to the protocol provided by kit manufacturers (BioLegend), (MabTech, Inc., Sweden). For both of these cytokines, sample O.D.s were measured by Stat Fax 3200 microplate reader (Stat Fax) and converted into desired concentration units using standard curves.
Coculture and leukocyte proliferation assay
ASCs/2-way mixed leukocyte reaction (MLR) coculture was performed to study the effect of unstimulated cortisol-treated ASCs on proliferative response of 2 allogenic peripheral blood mononuclear cells (PBMCs) to each other. For this purpose, 5,000 ASCs from each group that were suspended in DMEM containing Mitomycin C (10 μg/mL) were seeded in 96-well plate. After 2 h, the cells were washed 5 times with PBS/FBS 10% solution and incubated overnight in complete media. PBMCs were subsequently isolated from 2 healthy volunteers by Ficoll concentration gradient (1.077 g/mL) (GE Healthcare, Life Sciences, United Kingdom), and 5 × 104 PBMCs from each subject were used to perform 2-way MLR in ASCs containing wells. Allogenic proliferative response was assessed using combination of PBMCs from both donors to perform 2-way MLR.
Considering the aim of this study which is comparison of ASCs' immunomodulatory properties with or without HC presence, coculture of untreated ASCs with 2-way MLR was used as the control group. The cocultures were incubated for 4 days at 37°C in 5% CO2 and 95% humidity. The proliferative response of allogeneic PBMCs to each other was examined using 2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide (XTT)-assay (PromoCell GmbH, Germany), and O.D.s were measured on day 0, 2, and 4 of experiment using the Stat Fax 3200 microplate reader (Stat Fax). The results were reported as normalized proliferation ratio:
Data analysis
Data were analyzed using SPSS V.16 software (IBM Analytics). Cell proliferation data were analyzed by Mann–Whitney U test. Gene expression and cytokine levels were examined by 1-way ANOVA. *P < 0.05, **P < 0.01, and ***P < 0.001 were considered significantly different, and ns denotes P > 0.05.
Results
Characteristics of ASCs
The isolated cells from human adipose tissue were plastic adherent and spindle shaped (Fig. 1A). In addition, flow cytometry results indicated the presence of CD90, CD73, and CD105 markers, but no sign of CD34 and CD45 markers was detected on cell surface (Fig. 1B). In addition, isolated cells' ability to differentiate into chondrogenic, osteogenic, and adipogenic lineages was confirmed through their culture in differentiation media and specific staining (Fig. 1C). The combination of these observations and investigations confirmed that our isolated cells were fit into the mesenchymal stromal/stem cell category, and considering their adipose tissue source they were identified as ASCs.

ASC characteristics.
Gene expression pattern of HC-treated ASCs
Real-time PCR was carried out to assess expression of the immunomodulatory related genes in HC-treated ASCs. The relative expressions of COX-2, HGF, IDO, and PD-L1 genes were calculated and reported based on 2−ΔCt formula. In unstimulated condition, the expression ratio of COX-2 gene was 0.061 ± 0.005, 0.17 ± 0.017, 0.065 ± 0.006, and 0.016 ± 0.01 in the control, HC-5, HC-25, and HC-100 μM groups, respectively. The expression ratio of this gene was higher in HC-5 μM group compared to the others (P < 0.05). COX-2 expression in HC-25 μM group was altered relative to the control (P > 0.05); however, for HC-100 μM group, COX-2 expression was lower than other groups of this condition (P < 0.05).
The HGF expression ratio was 0.014 ± 0.004, 0.013 ± 0.003, 0.008 ± 0.001, and 0.007 ± 0.001 in the control, HC-5, HC-25, and HC-100 μM groups, respectively. The expression of this gene was changed in HC-5 μM compared to the control (P > 0.05). Meanwhile, the HC-25 μM group showed a significantly lower expression of this gene relative to both the control and HC-5 μM groups (P < 0.05). Alteration in HGF expression was also observed in HC-100 μM group compared to the control (P > 0.05).
The expression ratio of IDO gene in the control, HC-5, HC-25, and HC-100 μM groups was 0.00006 ± 0.00005, 0.00010 ± 0.00003, 0.00004 ± 0.00002, and 0.0001 ± 0.0001, respectively. No significant difference was seen in the HC-treated groups relative to the control (P > 0.05). The expression ratio of PD-L1 gene in control, HC-5, HC-25, and HC-100 μM groups was 0.0004 ± 0.0001, 0.0004 ± 0.00007, 0.0001 ± 0.00001, and 0.0001 ± 0.00003, respectively, and no significant difference was observed between them (P > 0.05).
In the stimulated condition, expression ratio of the COX-2 gene was 0.077 ± 0.003, 0.136 ± 0.01, 0.082 ± 0.01, and 0.037 ± 0.01 in the control, HC-5, HC-25, and HC-100 μM groups, respectively. The expression of this gene increased in HC-5 μM group compared to the control (P < 0.001) and decreased in HC-100 μM in relative to the control (P = 0.006). Furthermore, this gene expression in HC-25 μM group was significantly lower than HC-5 μM (P = 0.001) and higher than HC-100 μM groups (P = 0.004). In addition, the HC-100 μM group had a significantly lower expression of COX-2 relative to the HC-5 μM group (P < 0.001).
For HGF gene, the expression ratio in control, HC-5, HC-25, and HC-100 μM groups was 0.006 ± 0.002, 0.006 ± 0.002, 0.008 ± 0.0001, and 0.004 ± 0.002, respectively. No significant variation was observed in comparison of the cortisol-treated groups with each other, as well as in comparison with the control group. For IDO gene, expression ratio in the control, HC-5, HC-25, and HC-100 μM groups was 0.159 ± 0.02, 0.182 ± 0.049, 0.222 ± 0.029, and 0.205 ± 0.01, respectively. The expression of this gene was significantly increased in HC-25 μM group compared to the control (P = 0.035). However, no significant difference was observed in comparison of other HC-treated groups with control (P > 0.05). Besides, there was no significant difference in IDO expression between HC-treated groups.
Finally, PD-L1 expression ratio was 0.002 ± 0.0006, 0.005 ± 0.0003, 0.005 ± 0.002, and 0.007 ± 0.0008 in control, HC-5, HC-25, and HC-100 μM groups, respectively. The only significant difference was related to the increment of this gene expression in the HC-25 μM group compared to the control (P = 0.02) (Fig. 2).
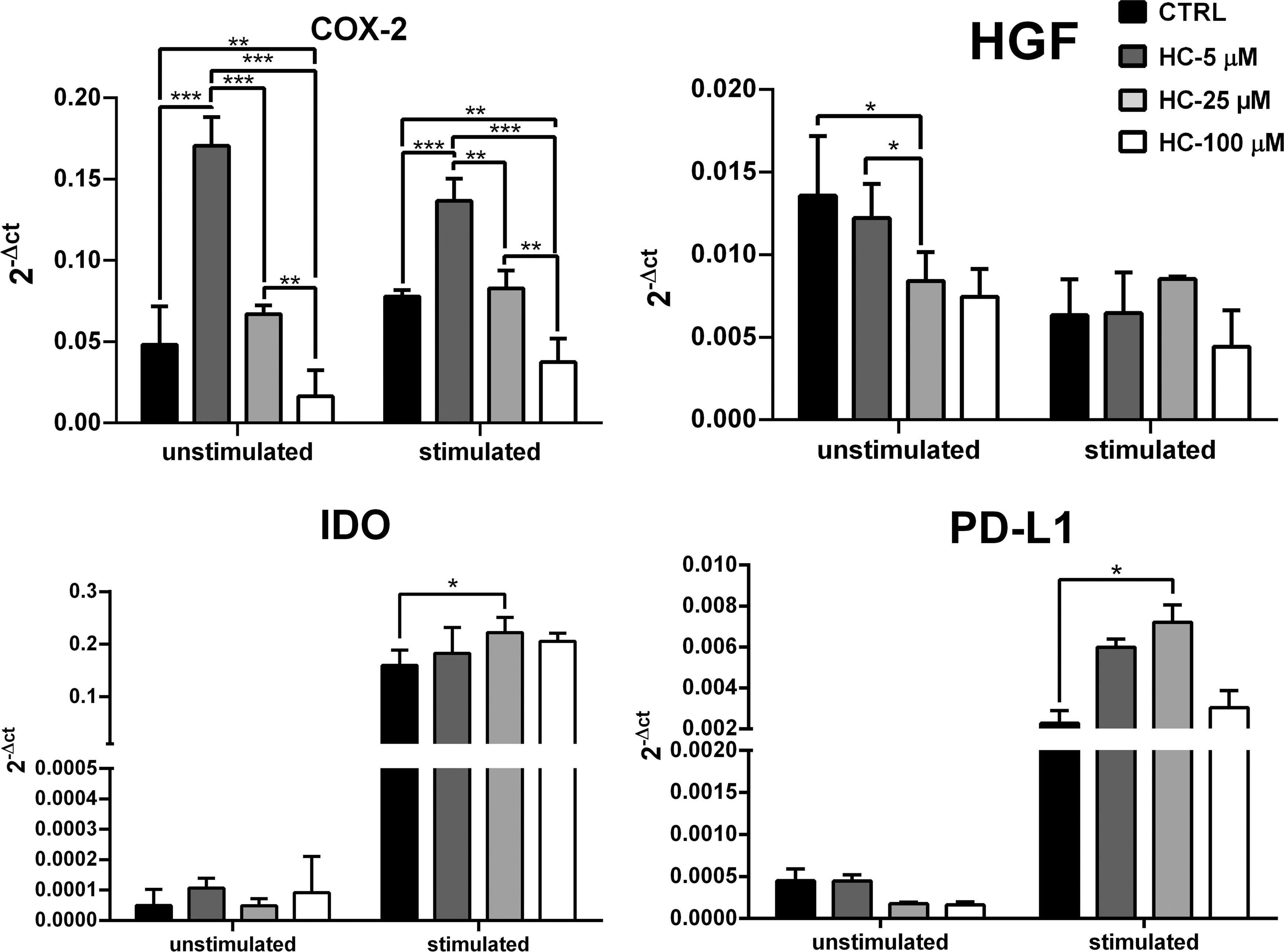
Evaluation of COX-2, HGF, IDO, and PD-L1 gene relative expression by real-time PCR. The expression ratio is presented as mean ± SD of 2−ΔCT. β-Actin was considered as reference gene. ***Indicates significance level of below 0.001, **indicates significance level of below 0.01, and *indicates a significance level of below 0.05. COX-2, cyclooxygenase-2; HGF, hepatocyte growth factor; IDO, indoleamine dioxygenase; PD-L1, programmed death-ligand 1.
Cytokine profile of HC-treated ASCs
The level of IL-6 and TGF-β1 in supernatants of all study groups was measured by ELISA and presented in pg/ml. In unstimulated condition, IL-6 level in control, HC-5, HC-25, and HC-100 μM groups was 5.382 ± 2.13, 2241.5 ± 915.105, 1082.750 ± 248.05, and 667 ± 339.26, respectively. Level of this cytokine increased in all of the HC-treated groups compared to the control. However, none of these differences was significant (P > 0.05). A downward concentration-dependent trend was detected in IL-6 level of HC-treated groups, but observed alterations were not significant among these groups (P > 0.05).
Levels of the TGF-β1 level in control, HC-5, HC-25, and HC-100 μM groups were 9.66 ± 0.8, 705.33 ± 204.3, 1710.93 ± 294.49, and 2060.53 ± 91.02, respectively. In general, concomitant with the increment in HC concentration, this cytokine level was elevated, which was significant for HC-5 (P = 0.001), HC-25, and HC-100 μM (P < 0.001) groups compared to the control. Besides, the increase of TGF-β1 level in HC-25 and HC-100 groups was significantly relative to the HC-5 μM group (P < 0.001).
In IFN-γ stimulated condition, IL-6 level in control, HC-5, HC-25, and HC-100 μM groups was 2786.25 ± 526.64, 23851.25 ± 3595.31, 35032.50 ± 733.50, and 12289.5 ± 4081.807, respectively. The level of this cytokine showed a significant increment in all 3 HC-treated groups compared to the control group (P < 0.001). However, IL-6 level was lower in HC-100 compared to HC-5 and HC-25 μM groups (P < 0.001).
The level of TGF-β1 in control, HC-5, and HC-25 μM groups was 802.93 ± 48.16, 910.4 ± 166.17, and 853.33 ± 34.8, respectively, and the increase of these 2 HC-treated groups was not significant compared to the control (P > 0.05). Nevertheless, the level of this cytokine in HC-100 μM group was 672.26 ± 46.18, which was significantly lower than the control (P = 0.046). There was also a significant decrease in TGF-β1 level at HC-100 μM group compared to HC-5 and HC-25 μM groups (P < 0.05) (Fig. 3).

Assessment of IL-6 and TGF-β1 cytokines. Level of these cytokines was measured by ELISA in culture supernatant and reported using pg/mL. Results represent the mean ± SD. ***Indicates significance level of below 0.001 and **indicates significance level of below 0.01. ELISA, enzyme-linked immunosorbent assay; IL, interleukin; TGF-β1, transforming growth factor beta 1.
HC-treated ASC effect on the 2-way MLR proliferative response
Proliferative response of allogenic PBMCs was measured using XTT on day 0, 2, and 4 of the ASCs/2-way MLR cocultures and reported as normalized proliferation ratio. On day 2 of coculture, the proliferation ratio of 2-way MLR significantly increased in the HC-5 μM group (0.48656 ± 0.089) relative to the control group's proliferation on the same day (0.34358 ± 0.023), (P = 0.046). However, the proliferation ratio of HC-25 (0.33615 ± 0.043) and HC-100 μM (0.28765 ± 0.010) groups was similar and decreased compared to the control on this day (P > 0.05). On day 4 of coculture, proliferation ratios of HC-5 (0.81539 ± 0.016) and HC-100 μM (0.83334 ± 0.013) groups decreased compared to the control (0.91139 ± 0.013) on the same day (P = 0.05), while on the same day, the HC-25 μM group (0.92230 ± 0.037) showed no difference relative to its control (P > 0.05) (Fig. 4).
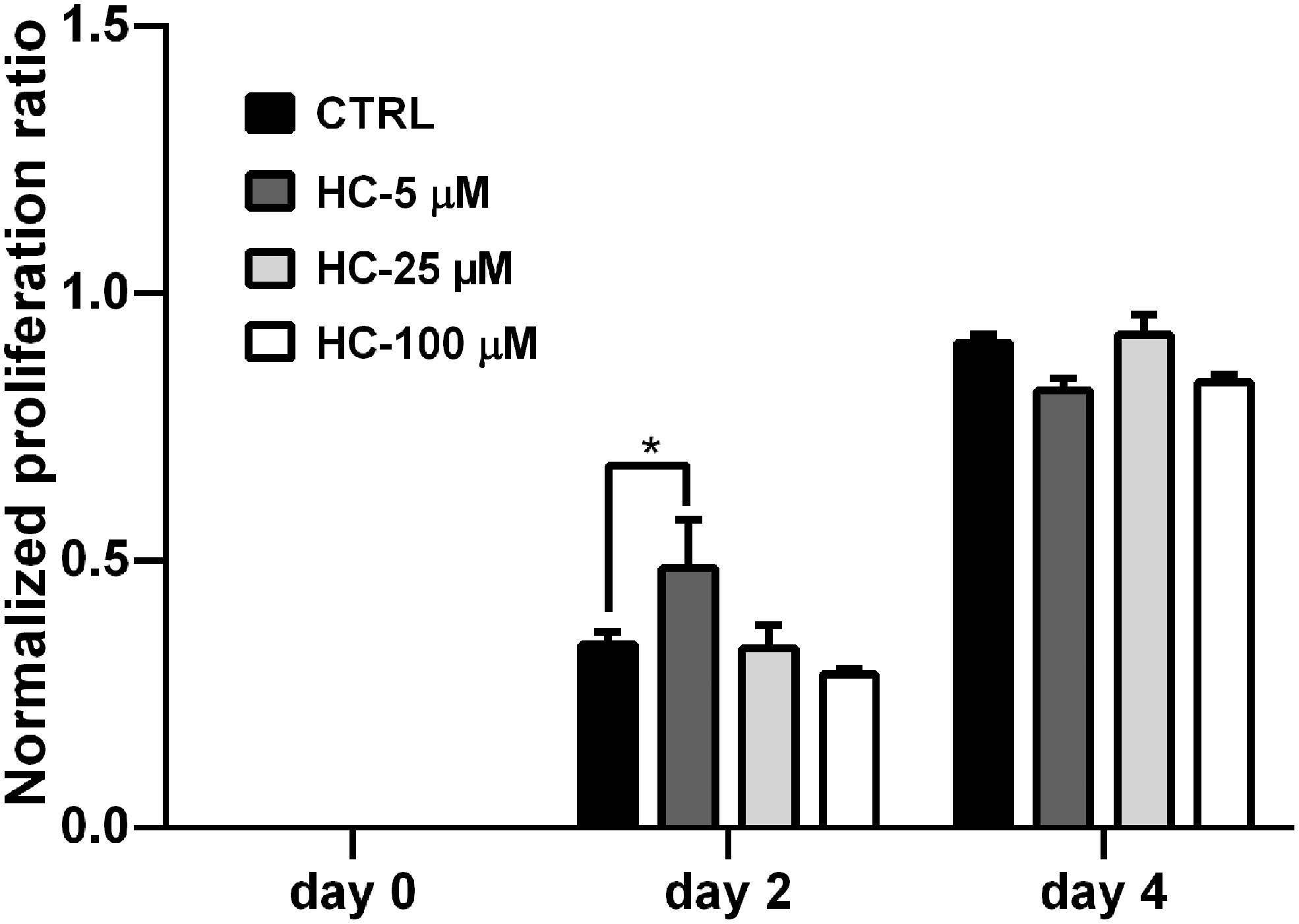
Proliferation rate of allogenic PBMCs in the ASCs/2-way MLR coculture. Effect of cortisol-treated ASCs on PBMC proliferation response in 2-way MLR was evaluated on days 0, 2, and 4 of the coculture using XTT and presented as normalized proliferation ratio. Results represent the mean ± SD. *Indicates a significance level below 0.05. MLR, mixed leukocyte reaction; PBMCs, peripheral blood mononuclear cells; XTT, 2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide.
Discussion
Cortisol is one of the most important hormones released into the bloodstream during stress, affecting the metabolism of lipids and carbohydrates and leading to the accumulation of oxidative stress products at the cellular level (Myint and others 2017; Gouda and others 2018). Adipose tissue is a target for cortisol, causing changes in adipogenesis, metabolism, and secretion of adipokines (Lee and others 2014). Meanwhile, ASCs are important cells located in adipose tissue that can play a role in various functions such as cell repair and differentiation, through their immunomodulatory features (Bobis and others 2006).
The physiological dose of cortisol hormone in plasma is about 1 μM, which is believed to vary in circadian rhythms. In stress conditions, depending on the type of stress (acute or chronic), the stress may increase cortisol concentration up to 5 times (Gopi and Rattan 2019). Besides, this hormone is lipophilic, so it can easily transit across the cell membrane (Flück and Miller 2008); thus, the actual accessibility to this hormone (or HC) can be far more than its plasma level.
Due to the observation of continually upregulated levels of cortisol in stressed individuals, also, considering the application of high HC concentrations to control symptoms of several chronic diseases, including COPD, rheumatoid arthritis, and SLE (Fan and Morand 2012), we have decided to include different range of HC in our study.
To evaluate the long-term effects of HC on ASCs' immunomodulatory related features, we treated ASCs with 5, 25, and 100 μM concentrations of this hormone for 14 days. Genes and cytokines involved in immunomodulatory properties of HC-treated ASCs were evaluated in IFN-γ stimulated and unstimulated conditions. Some immunomodulatory related features of ASCs were examined by measuring the expressions of COX-2, HGF, IDO, and PD-L1 genes, as well as the levels of TGF-β1 and IL-6 cytokines. In addition, ASCs/2-way MLR cocultures were performed to investigate allogenic PBMCs' proliferative response in presence of HC-treated ASCs.
The ability of ASCs to exert immunomodulatory effects is partly related to their secretory factors. IL-6 is a pleiotropic cytokine with broad effects on immune responses. This cytokine is secreted by several cells in the body, including immune and stromal cells, and is involved in activation and proliferation of B and T lymphocytes (Jordan and others 2017). MSCs also secrete considerable amounts of IL-6, a feature that may be implicated in tissue regeneration by these cells (Gu and others 2016). Studies indicated that IL-6 production increases following the activation of GR and forming a complex with STAT3. The presence of IFN-γ also enhances this mechanism (Takeda and others 1998; Campia and others 2015). Dong and others (2018) reported that cortisol decreased macrophage IL-6 production in a concentration-dependent manner, through reduction of NF-κB.
Our results showed that long-term exposure to HC generally increased IL-6 production from ASCs in both stimulated and unstimulated conditions. Previous studies showed that cortisol removes one of the IL-6 production barriers through inhibition of mTOR (mammalian target of Rapamycin) and increases the level of this cytokine (Liao and others 2017). mTOR is a protein kinase involved in metabolism and cellular functions such as reception of messages related to cellular stress. mTOR signaling can be effective in reducing IL-6 gene expression. Glucocorticoids (cortisol and dexamethasone) have been shown to exert an effect similar to mTOR inhibitor (rapamycin) (De Martino and others 2012; Wang and others 2017). Consistent with these studies, our results indicated the increment of IL-6 production and the possible inhibition of mTOR signaling pathway by HC in ASCs.
TGF-β1 is a multifunctional cytokine with broad spectrum of cellular activities such as proliferation, differentiation, migration, repair, and immunomodulatory properties (Xu and others 2018). TGF-β1 plays an important role in the regulation of immune responses and is able to inhibit T lymphocytes during inflammation (Kyurkchiev and others 2014). Yang and others (2014) showed that cortisol through reducing TGF-β1 in inflammatory milieu disrupts the healing process of damaged mouse liver tissue. This cytokine can be produced by MSCs (Shi and others 2012). Wang and others (2017) study revealed that exposure of rapamycin-treated MSCs to inflammatory condition (IFN-γ+TNF-α) reduces their expression of TGF-β1 gene.
In the unstimulated condition of our study, HC-treated ASCs increased TGF-β1 in a concentration-dependent manner. In addition, in the same condition, the increment in HC concentration caused an inverse pattern between TGF-β1 and IL-6 secretion, so in this process, increasing HC concentration rises TGF-β1 level while the IL-6 level reduces. It seems that the outcome of high HC concentration in unstimulated condition is likely to direct ASCs toward an anti-inflammatory and regenerative profile, while the trend of these 2 cytokines in ASCs treated with lower concentration of HC may point to development of the pro-inflammatory profile in ASCs. In addition, Inoue-Mochita and others (2018) demonstrated that high concentrations of IL-6 inhibit trans-signaling pathways associated with the production of TGF-β1 from ASCs.
In our study, secretion of TGF-β1 from HC-treated ASCs decreased in the IFN-γ-stimulated condition, which may be due to an excessive increase of IL-6 in these groups. Furthermore, it seems that the tendency of TGF-β1 changes in this condition is due to presence of IFN-γ in ASC microenvironment, which directed them toward a pro-inflammatory phenotype. As we have shown in our previous research, IFN-γ increases the pro-inflammatory function of ASCs by priming these cells (Aliakbari and others 2019). Therefore, concentration and duration of IFN-γ stimulation in the present study might be accountable for alteration in ASC cytokine secretion profile.
Regarding gene expression, in general we found that HC affects target gene expression in lower and higher concentration differently and also IFN-γ stimulation can modify this process. In detail, we found that stimulation with IFN-γ increased the expression of PD-L1 and IDO genes. Other studies have also shown higher expression of these molecules in the presence of IFN-γ (Ankrum and others 2014; Javorkova and others 2018). PD-L1, which is expressed on antigen presenting cells and tumor cells, interacts with its receptor (PD-1) on the surface of T and B lymphocytes, as well as myeloid cells, reducing the proliferation of immune cells and inhibiting their function (Wu and others 2019). MSC expression of PD-L1 and its role in inhibition of immune cells have been confirmed previously (Davies and others 2017).
IDO carries out the first step of tryptophan catabolism in cells, and its production is induced by IFN-γ (Meisel and others 2004). This enzyme is produced by MSCs as an immunosuppressive factor, arrests T cell proliferation, and induces Treg cells (François and others 2012). Earlier studies have demonstrated the similarity of IL-6 and PD-L1 expression patterns (Takeda and others 1998; Ding and others 2019). Increased IL-6 level in the presence of IFN-γ appears to augment IDO and PD-L1 levels by generating a positive feedback (Litzenburger and others 2014; Cheng and others 2016; Ding and others 2019).
Similar to these studies, our findings showed increasing PD-L1 expression along with the increment of IL-6 production. Glucocorticoid effect on MSC expression of IDO and PD-L1 has been shown. For instance, budesonide and dexamethasone increase IDO expression from MSCs in the presence of IFN-γ (Ankrum and others 2014; Javorkova and others 2018). However, Javorkova and others (2018) reported that short-term treatment with dexamethasone in the presence of IFN-γ reduces PD-L1 expression in ASCs.
In the IFN-γ-stimulated condition of our research, a higher expression of PD-L1 and IDO was observed. Lower concentrations of HC increased PD-L1, while higher HC concentration was not able to do this. However, in the unstimulated condition, HC did not alter the expression of this gene. Deng and others (2019) reported that the inhibition of mTOR signaling increases PD-L1 expression on the surface of nonsmall cell lung carcinoma cell line. Therefore, further expression of IDO and PD-L1 in the stimulated condition is probably due to the inhibition of mTOR signaling by HC that indirectly affects the expression of these genes.
COX-2 gene expression is induced by inflammatory stimuli, hormones, and growth factors and is a major factor in the production of prostanoids in inflammation. This enzyme has a dual role in regulating immune responses, so that the overexpression of this molecule leads to increment of inflammatory mediators and enhancement of cell proliferation. In contrast, COX-2 function leads to PGE2 production, which is known as an immune response inhibitor (Gandhi and others 2017).
Results of our research revealed that ASCs treated with lower HC concentrations in both stimulated and unstimulated conditions showed higher expression of COX-2, while ASCs treated with 100 μM of HC demonstrated lower expression of this gene. It has been shown that raised IL-6 level has also a positive effect on the production of COX-2 and PGE2 and indirectly increases the immunosuppressive function of MSCs (Bouffi and others 2010). In line with this study, we observed an increment in COX-2 expression along with elevated IL-6 level in ASCs treated with low concentration of HC.
The dual behavior of cortisol in altering COX-2 expression has been previously demonstrated. Yang and others (2014) examined the expression of COX-2 in cortisol-treated human fetal lung fibroblasts (HFL-1) and human amnion fibroblasts. Their result showed that high concentration of cortisol decreased the function of NF-κB, thus reducing the expression of COX-2 from HFL-1 cells. However, through phosphorylation of COX-2 promoter (CREB), cortisol increased the expression of this enzyme in human amnion fibroblasts cells (Zhu and others 2009).
Another study showed that cortisol, in a concentration-dependent manner, inhibited the expression of COX-2 mRNA in macrophage cell line of RAW264.7. This study suggests that high cortisol concentration reduces the function of NF-κB by increasing its inhibitor molecule (iκB) and eventually diminishes the expression of COX-2 (Dong and others 2018). Given the similarities between our results with these studies, it is possible that both of described pathways have played a role in shaping the trend of COX-2 gene expression in our study.
HGF is a growth factor secreted by various cells, including MSCs, and is involved in their migration, proliferation, and immunomodulatory and regenerative functions (Madrigal and others 2014). ASCs basically have a high expression of HGF, which could be due to the role of these cells in tissue regeneration and repair (Rehman and others 2004). Our findings also showed high basal expression of HGF in untreated ASCs. Wang and others (2017) reported that MSCs treated with mTOR inhibitor expressed lower HGF levels when stimulated with IFN-γ and TNF-α.
Our results also genuinely revealed decreasing of HGF expression in the presence of different concentrations of HC. Earlier studies proposed that cortisol can destabilize mRNA of HGF and hence reduces production of this protein, which impairs HGF associated healing processes (Blanquaert and others 2000). Consistent with our results, a study has shown the downward pattern of HGF expression from Dexamethasone-treated ASCs in IFN-γ-stimulated condition (Javorkova and others 2018).
We also investigated the effect of HC-treated ASCs on the proliferative response of allogeneic PBMCs to each other in ASCs/2-way MLR coculture. Overall, untreated ASCs/2-way MLR cocultures showed less proliferative allogenic response compared to 2-way MLR; also cocultures of ASCs treated with different concentrations of HC did not suppress the proliferation of PBMCs.
On day 2 of this experiment and at the beginning of allogeneic response, ASCs treated with HC-5 and HC-100 μM concentrations had an inverse effect on PBMC proliferation in 2-way MLR. The HC-100 μM treated ASCs suppressed proliferation (P > 0.05), while the HC-5 μM treated ASCs enhanced proliferation on this day. Chan and others (2006) showed that BM-MSCs reacted differently to various amounts of IFN-γ, so that MSCs act as APCs in lower amounts of this cytokine, but with the increment of IFN-γ level, MHC-II was reduced on MSC surface and therefore they contribute in suppression of PBMC proliferation. Given that IFN-γ increases at the beginning of MLR response (Aliakbari and others 2019), we may say that there is a similar cytokine balance between day 2 of ASCs/2-way MLR coculture and the stimulated condition of our study.
Treatment of ASCs treated with low HC concentration appears to benefit the priming of these cells, while HC-100 μM reduced this process. In contrast, given the cytokine balance proposed on day 2 of experiment, further proliferation of PBMCs in coculture with HC-5 μM treated ASCs could be due to presumed association between HC and IFN-γ, which might have elevated IL-6 levels.
According to a study, IL-6 shifts T lymphocyte differentiation toward T helpers, which leads to expansion and activation of B cells (Eddahri and others 2009; Najar and others 2009). Therefore, the increment of PBMC proliferation in our study might be due to elevated level of IL-6 in coculture media of day 2. As ASCs/2-way MLR coculture continued on day 4, HC-5 and HC-100 μM treated ASC showed the same ability to suppress the proliferation of PBMCs (P > 0.05). Consequently, HC-treated ASCs reduced the proliferative response of allogeneic PBMCs at day 4 of ASCs/2-way MLR coculture.
Conclusion
MSCs have a special role in homeostasis, as well as in tissue repair and control of inflammation. Considering the cortisol impact on physiologic and metabolic stresses and its pharmaceutical use in chronic inflammatory diseases, the present study examined the HC long-term treatment (14 days) effect on ASCs' immunomodulatory properties. The result of this study revealed that high and low concentrations of HC affected ASCs' immunomodulatory properties differently. It seems that low concentration of HC prepares ASCs to take part in inflammatory response through variations in IL-6 and TGF-β1 levels along with COX-2 elevated expression in unstimulated condition. However, in the same condition, the excess of TGF-β1 over IL-6 and the dramatic reduction of COX-2 at high concentration of HC can ultimately push the ASCs toward modulation of immune responses.
Considering the role of IFN-γ in priming ASCs, in stimulated condition of our study, IFN-γ presence may be responsible for extensive changes in factors like IL-6 and IDO, although treatment with different concentration of HC also played a role in these alterations. Similar to the unstimulated condition, the changes observed in IL-6 level and COX-2 expression at HC low concentration in stimulated condition imply a pro-inflammatory phenotype, and alteration of these 2 and IDO gene at high concentration can be interpreted as the signs of anti-inflammatory phenotype in HC-treated ASCs.
In conclusion our results suggest that prolonged exposure with low concentration of HC may lead to a progressive inflammatory response through ASCs, while exposure to high concentrations of HC can alter ASCs' immunomodulatory properties in favor of immunosuppression. Our findings provide an insight on HC effects on ASCs and could be considered in management of lasted physiologic and pathologic stresses, also patients undergoing prolonged HC treatments. However, these results' confirmation and understanding HC interactions with ASCs need further investigation.
Footnotes
Authors' Contributions
Study Design: M.R.R. Data Collection: S.S., R.K., M.M., S.A. Statistical Analysis: S.S., M.M., A.A.A. Data Interpretation: M.M., M.R.R., S.S., A.A.A. Article Preparation: S.S., M.M., S.A., R.K. Literature Search: S.S. Visual Abstract: M.M. Funds Collection: M.R.R.
Acknowledgment
The authors thank Dr. Mahbubeh Sabokdast for helping them in providing human adipose tissue samples.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Research Council of Kurdistan University of Medical Sciences [1398.247].