
Abstract
The purpose of this experiment is to find out the function of Vitamin D (VD) in airway inflammation in asthmatic guinea pigs by regulating mammalian target of rapamycin (mTOR)-mediated autophagy. A total of 40 male guinea pigs were randomly assigned into the Con group, the ovalbumin (OVA)-sensitized group, the VD group, the VD + dimethyl sulfoxide group, and the VD + rapamycin (mTOR inhibitor) group. Then, serum from all groups was harvested for the measurement of immunoglobulin E (IgE), interleukin (IL)-4, and IL-5 levels. Next, bronchoalveolar lavage fluid was collected for cell counting. Moreover, lung tissues were extracted to assess levels of p-mTOR and autophagy factors (LC3B, Beclin1, Atg5, and P62). Compared with the Con group, the OVA group showed elevated levels of IgE, IL-4, and IL-5, increased contents of eosinophils, neutrophil, and lymphocytes, and declined monocytes. And the VD group improved inflammatory reactions in the guinea pigs. Besides, the OVA group showed lower levels of p-mTOR and P62 and higher autophagy levels than the Con group, while the VD group had opposite results. Rapamycin annulled the suppressive role of VD to airway inflammation in asthmatic guinea pigs. VD might inhibit OVA-induced airway inflammation by inducing mTOR activation and downregulating autophagy in asthmatic guinea pigs.
Color images are available online
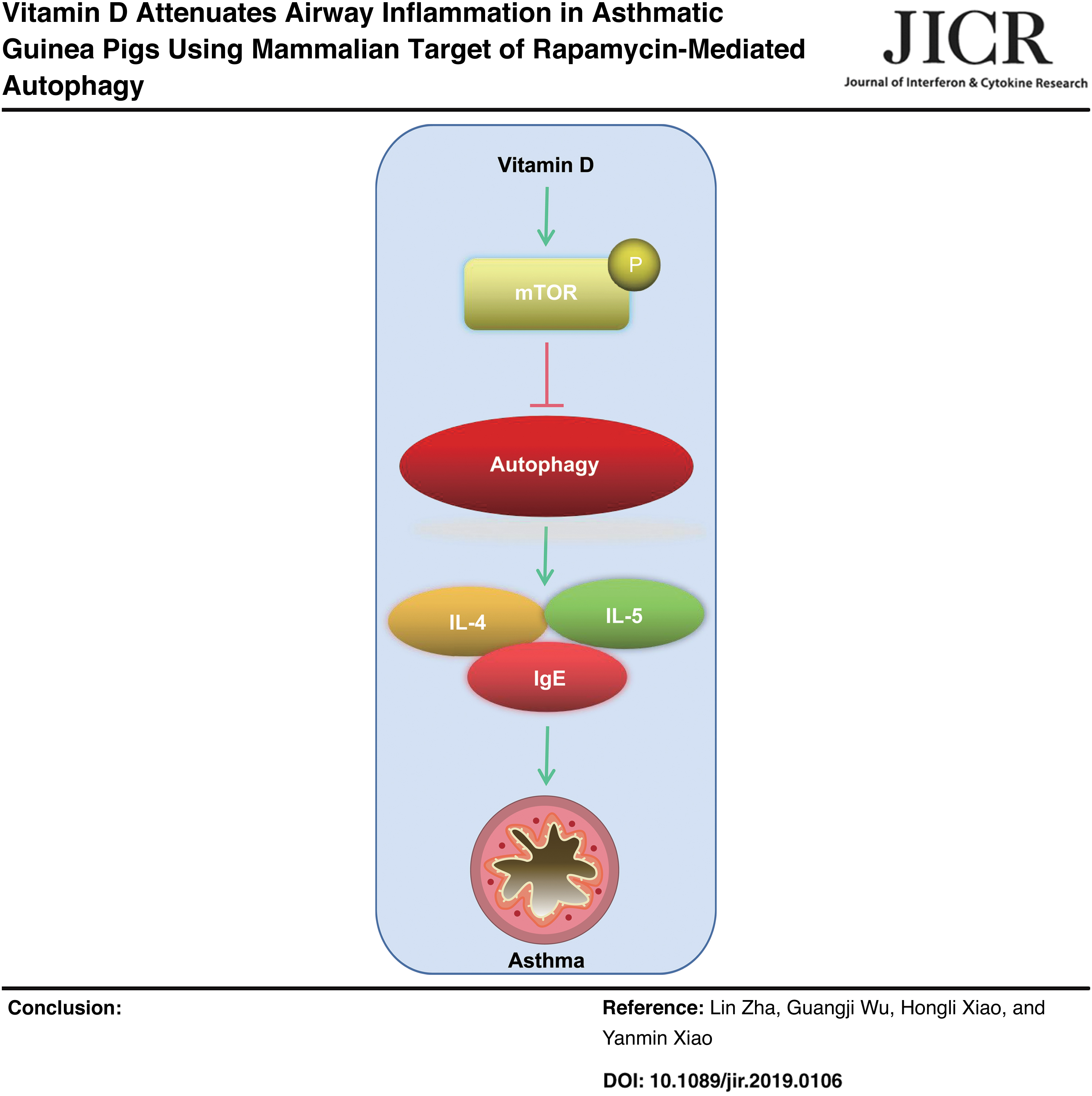
Introduction
Asthma represents a kind of complicated respiratory disease, which is featured by heterogeneity in the disorder risky factors and reactions of patients toward treatment (Robinson and others 2017). As a chronic inflammatory dilemma happening in the respiratory tract, asthma is responsible for the subsequent cough, chest distress, wheeze, and shortness of breath, and it is related to multiple complications, embracing gastroesophageal reflux disease, hormonal disorders, sinusitis, obstructive sleep apnea, and rhinitis (Boulet and Boulay 2011; Mims 2015). Prematurity, obesity, cigarette smoking, and pulmonary infection are perilous causes of asthma pathogenesis (Wu and others 2019b). Inflammatory responses in airways emerge as a marked indicator in severe asthma (Finkas and Martin 2016). Besides, abnormally modulated autophagy coupling with unleashed release of inflammatory cytokines is also a principal characteristic of asthma (Sachdeva and others 2019).
Therapies of asthma are classified according to the syndrome severity, physical exercise, peak expiratory flow rates, and the first of expiration (Kwah and Peters 2019). Curative treatment targeting asthma could reassess asthma, effectively regulate this disease, reduce negative consequence, and discourage inflammatory reactions (Wu and others 2019b). Despite this, asthma remains a severe disorder in both teenagers and the aged as it poses great threat to their happiness and welfare and thus renders disappointing clinical consequences and growing rates of incidence and mortality (Wu and others 2019a).
That is to say, to figure out effective interventions is an essential part in alleviating asthma. Toward this, we noticed Vitamin D (VD), the key factor in the processes of bone metabolism and calcium homeostasis, which plays major role in different carcinomas and autoimmune disorders through suppressing cellular dissemination, prompting cell differentiation, regulation of immune system, and hormonal systems (Dusso and others 2005). Since VD depletion is emerging as a health burden around the world, incidence rate of autoimmune disorders is accordingly rising (Bizzaro and others 2017).
Jolliffe and others (2017) unmasked that administration of VD quenches the danger of asthma severity, whose treatment requires systemic corticosteroids (Jolliffe and others 2017). According to a recent report, VD could render an inhibitory effect on asthma through the suppression of inflammation by positively controlling immunologic axis and cellular activities of antigen-presenting cells, mast cells, and structural cells, as well as the biological behaviors of lymphocytes (Pfeffer and Hawrylowicz 2018). Furthermore, VD is also a modulator in the performance of autophagy and antimicrobial peptides (Jin and others 2015). On the other hand, VD protects against lipopolysaccharide-induced acute lung injury by balancing level of rennin-angiotensin system (Xu and others 2017). Robustly expressed VD in serum predicts a dampened hazard of lung cancer (Zhang and others 2015).
Moreover, VD administration strengthens pulmonary functions and contributes to the recovery of lung injuries after cystic fibrosis (Chesdachai and Tangpricha 2016). Thence, we attempt to investigate the potential function of VD in asthma. It is recently reported that VD is able to alleviate inflammatory reaction and autophagy by upregulating mammalian target of rapamycin (mTOR) (Kong and others 2020). Moreover, mTOR downregulation is responsible for the unremitting inflammatory symptoms and autophagy in airway of asthmatic mice (Li and others 2020). Generally speaking, it may be promising to speculate that VD could participate in asthma progression by mediating inflammation and autophagy through mTOR activation.
Materials and Methods
Ethics statement
This study was approved and supervised by the ethics committee of Puren Hospital Affiliated to Wuhan University of Science and Technology. The protocol was also approved by the Institutional Animal Care and Use Committee of Puren Hospital Affiliated to Wuhan University of Science and Technology. Significant efforts were made to minimize both the number of animals used as well as their respective suffering.
Laboratory animals and grouping
Forty SPF male guinea pigs (250–300 g) (Hunan SJA Laboratory Animal Co., Ltd., Changsha, Hunan, China) were allowed to acclimate for 1 week with relative humidity (55%) and 22°C–24°C temperature for a 12-h light/12-h dark cycle with free access to food and water. Then, the guinea pigs were allotted into the control (Con) group (guinea pigs were subjected to 1.0 mL normal saline through intraperitoneal injection), the ovalbumin (OVA) group (guinea pigs were sensitized by OVA followed by an intraperitoneal injection of 1.0 mL normal saline), the VD group (guinea pigs were sensitized by OVA followed by a treatment with VD), the VD + dimethyl sulfoxide (DMSO) group (guinea pigs were sensitized by OVA followed by a treatment with VD and DMSO), and the VD + rapamycin [mTOR) inhibitor group (guinea pigs were sensitized by OVA followed by a treatment with VD and rapamycin) (n = 8).
Both the OVA group and the VD group were treated with 10% OVA (grade V; Sigma-Aldrich, Merck KGaA, Darmstadt, Germany) and 1.0 mL normal saline through intraperitoneal injection. After 10 days, guinea pigs were placed in atomizing gas with a mixture of 1% OVA and normal saline to challenge asthma every other day for 15 times altogether (10 min each time). The Con group was injected with the same amount of normal saline intraperitoneally, and 10 days later, guinea pigs in the Con group were placed in atomizing gas with normal saline every other day for 15 times altogether (10 min each time) (Ma and others 2021). Two hours before every sensitization, 1.0 mL sterile normal saline was intraperitoneally injected into the Con group and the OVA group, while the VD group was intraperitoneally injected with the mixture of 5 × 105 U/kg VD and 1.0 mL normal saline.
To elaborately clarify the role of mTOR activation in the process of VD affecting allergic airway inflammation, rapamycin (Sigma) was prepared in normal saline containing 20% DMSO and then injected intraperitoneally 30 min before each administration of VD, with the injection of an equal amount of DMSO as the control (Zou and others 2019). The protocol of treatment of guinea pigs in each group is listed on Fig. 1.
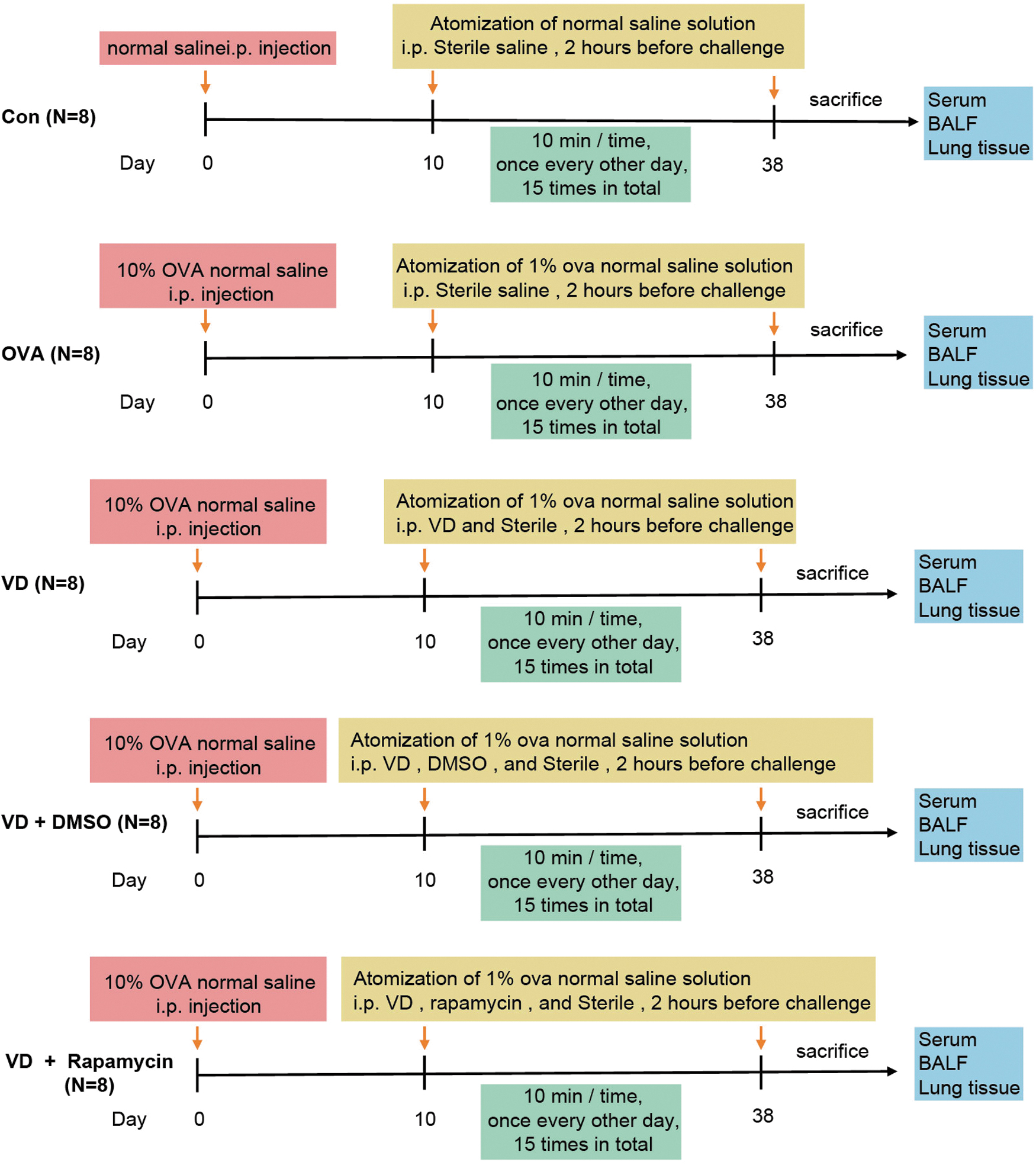
The protocol of treatment of guinea pigs in each group.
Sample collection
After the last atomizing gas sensitization, guinea pigs were euthanatized with excessive pentobarbital sodium (100 mg/kg) through intraperitoneal injection. Lung tissues, bronchoalveolar lavage fluid (BALF), and serum sample were extracted. Guinea pig abdominal aortic blood was harvested to extract serum, which was then stored under −80°C for the following enzyme-linked immunosorbent assay (ELISA). BALF was harvested as previously reported (Bolcas and others 2019; Ye and others 2019) and for the calculation of cell number. Lobus dexter was fixed by 10% formalin for histological analysis of lung; while left lobe was placed under −80°C for the following Western blot analysis.
Enzyme-linked immunosorbent assay
After the last atomizing gas sensitization, serum of guinea pigs was collected and examined for levels of immunoglobulin E (IgE), interleukin (IL)-4, and IL-5 in the guinea pigs in each group, determined in strict accordance with the instructions of the kits (BioLegend, San Diego, CA) following the previous literature (Zhang and others 2018). Eventually, optical density (OD) at 450 nm was measured using a microplate reader.
BALF cell counting
BALF was collected, centrifuged, and then counted for the total number of cells by a blood cell counter (Thermo Fisher Scientific, Inc., Waltham, MA) (Bolcas and others 2019). When cells were transferred onto a slide, they were stained by a HEMA3 staining apparatus (Fisher Scientific). After the refreshment of the cover on the slide, the slide was identified. Subsequently, 200 cells were counted and the total cells in BALF were analyzed to calculate the percentage of neutrophils, eosinophils, lymphocytes, and macrophages.
Hematoxylin and eosin staining
Lung tissues were fixed in 4% paraformaldehyde and phosphate-buffered saline (PBS), paraffin embedded, sliced, stained with hematoxylin for 5 min, rinsed by PBS, differentiated by ethanol hydrochloride for 3 s, stained with eosin for 2 min, and fixed by neutral resins. In addition, pathological changes of the right bronchoalveolar tissues were observed by an optical microscope (Olympus Optical Co., Ltd, Tokyo, Japan). Pulmonary inflammation was assessed by peribronchovascular inflammatory cell infiltration (0, none; 1, mild; 2, moderate; 3 obvious; and 4, severe) (Huang and others 2019).
Western blot analysis
Bronchoalveolar tissues from guinea pigs were lysed on ice and centrifuged using radio-immunoprecipitation assay buffer consisting of protease inhibitor (Ma and others 2021). Then, protein concentration was measured by a protein bicinchoninic acid kit. Afterward, the proteins were transferred onto the polyvinylidene fluoride membranes after being separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis. Subsequently, the membranes were incubated with the following primary antibodies: nuclear factor mTOR (1: 1,000, #2972; Cell Signaling Technology (CST), Beverly, MA), p-mTOR (1:1,000, #2971; CST), light chain (LC) 3B (1:1,000, #3868; CST), Beclin1 (1:1,000, #3738; CST), Atg5 (1:2,000, ab108327; Abcam, Inc., Cambridge, MA), p62 (1:1,000, #5114; CST), and glyceraldehyde-3-phosphate dehydrogenase (1:1,000, #8884; CST) at 4°C overnight.
Next, the membranes were rinsed with Tris-buffered saline-tween buffer. Subsequently, membranes were cultured with horseradish peroxidase-labeled secondary antibody at room temperature for 2 h. In addition, protein band system OD was observed using the electrochemiluminescence kits (Bio-Rad Laboratories, Hercules, CA) and gel imaging, analyzed, and quantified by an image tool.
Statistical analysis
The results are shown in mean ± standard deviation. One-way analysis of variance (ANOVA) was used for comparing different groups and Tukey's multiple comparisons test for pairwise comparisons after ANOVA. GraphPad Prime v8.0 (GraphPad Software, San Diego, CA) was employed to get data analysis. P was attained using 2-tailed test and P < 0.05 indicated significant difference.
Results
VD reduces levels of IgE, IL-4, and IL-5 in the serum of asthmatic guinea pigs
At the beginning of this study, levels of IgE (Fig. 2A), IL-4 (Fig. 2B), and IL-5 (Fig. 2C) in the serum of asthmatic guinea pigs from each group were analyzed through ELISA. Levels of IgE, IL-4, and IL-5 in the OVA group were higher than those in the Con group (all P < 0.05). Meanwhile, the VD group showed quenched levels of IgE, IL-4, and IL-5 compared with the OVA group (all P < 0.05), illustrating that VD could adjust levels of IgE, IL-4, and IL-5 to normal performance; thus, it may be a possible target in asthma treatment.
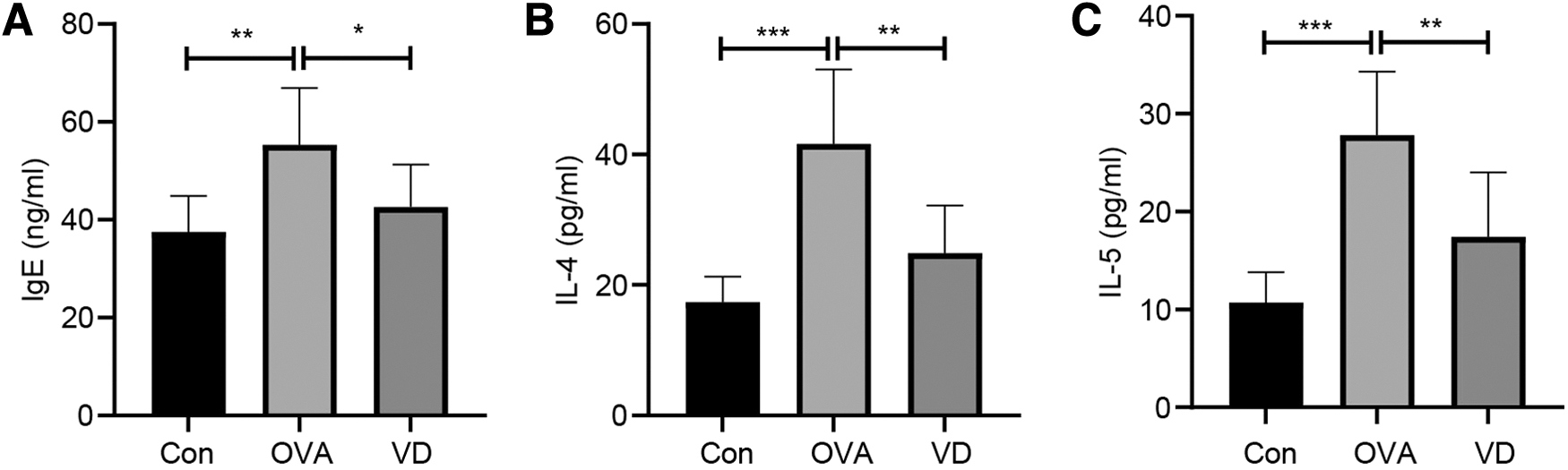
VD reduces levels of IgE, IL-4 and IL-5 in the serum of asthmatic guinea pigs.
VD regulates cell counts in the BALF from asthmatic guinea pigs
To investigate the effect of VD on airway inflammation of asthmatic guinea pigs, BALF was extracted from euthanatized guinea pigs and then analyzed (Fig. 3A–E), from which it was shown that compared with the Con group, the OVA group exhibited the elevated contents of total cells in BALF, eosinophils, neutrophils, and lymphocytes, but limited number of monocytes (all P < 0.05). However, the VD group had the opposite consequences (all P < 0.05). That is to say, VD restricted OVA-induced airway inflammation.
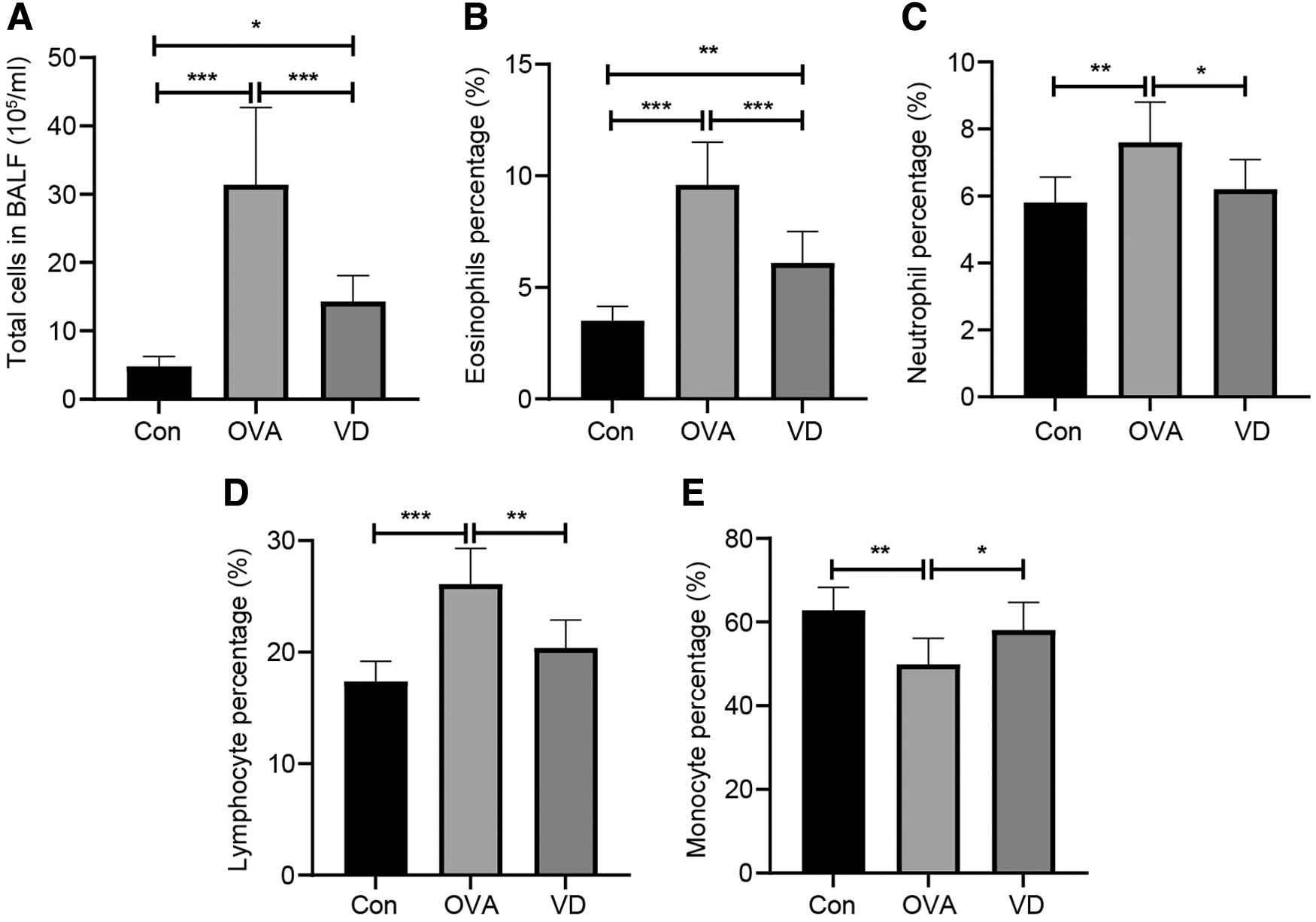
VD regulates cell counts in the BALF from asthmatic guinea pigs,
VD reverses inflammatory cell infiltration in lung tissues from asthmatic guinea pigs
Our experiment further analyzed inflammation in pathological lung tissues from each group. According to the results of hematoxylin and eosin (H&E) staining (Fig. 4A), lung tissue sections from the Con group showed no abnormally pathological change in tissue. The OVA group expressed thickened bronchial smooth muscle, narrowed alveolar space, and conspicuous inflammatory cell infiltration in bronchus, perivascular region, and alveolar space, which were reversed by VD administration. Compared with the Con group, both the OVA group and the VD group had increased staining scores, and the VD group exhibited lower staining score than the OVA group (Fig. 4B). Although the VD group showed less pathologically abnormal performance than the OVA group, it remained a small amount of inflammatory cell infiltration. The above results supported that VD reduced OVA-induced inflammatory cell infiltration.
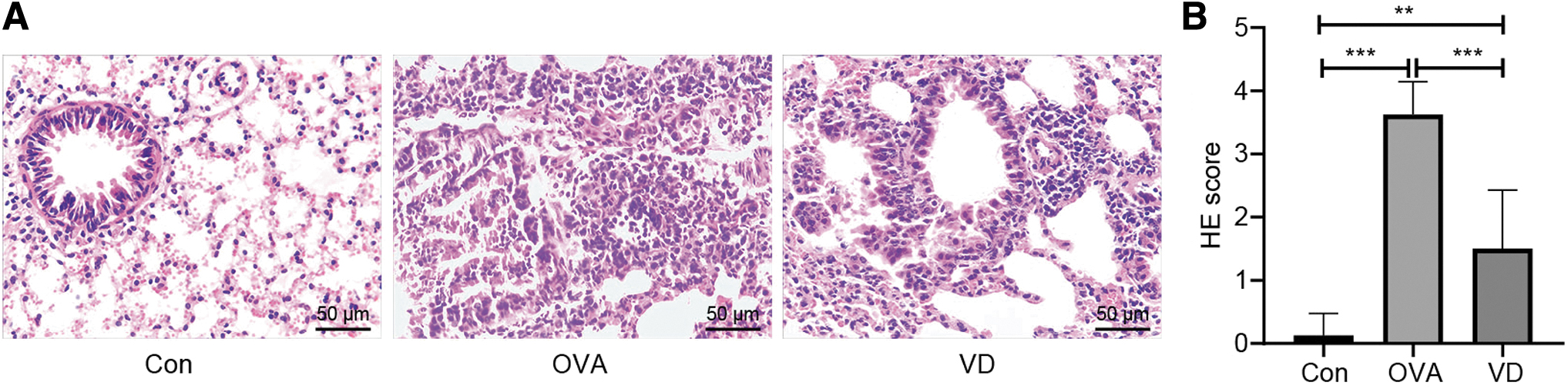
VD reverses inflammatory cell infiltration in lung tissues from asthmatic guinea pigs.
VD enhances mTOR expression and inhibits autophagy in lung tissues from asthmatic guinea pigs
mTOR-mediated autophagy plays a dominant role in allergic airway inflammation (Zou and others 2019; Li and others 2020). However, the mechanism of VD in regulating mTOR-mediated autophagy in asthma is to be investigated. Therefore, mTOR level (Fig. 5A, B) and autophagy cytokines (LC3B, Beclin1, Atg5, and p62) in lung tissues of guinea pigs in each group were determined by Western blot analysis (Fig. 5C, D), and the results showed that lung tissues of guinea pigs in the OVA group possessed lower levels of p-mTOR/mTOR and p62, but higher levels of autophagy cytokines LC3II/LC3I, Beclin1, and Atg5 than those in the Con group. Compared with the OVA group, the VD group showed promoted levels of p-mTOR/mTOR, degraded levels of autophagy cytokines LC3II/LC3I, Beclin1, and Atg5, and increased p62 level (all P < 0.01). It was indicated that VD could activate mTOR and inhibit autophagy level in OVA-treated guinea pigs.

VD enhances mTOR expression and inhibits autophagy in lung tissues from asthmatic guinea pigs.
VD palliates airway inflammation in asthmatic guinea pigs by activating mTOR and suppressing autophagy
To further explore the effect of mTOR activation on the process of VD affecting airway inflammation in asthmatic guinea pigs, the rapamycin was intraperitoneally injected 30 min before the administration of VD. Levels of IgE, IL-4, and IL-5 (Fig. 6A–C) in the serum of guinea pigs of each group were analyzed by ELISA. Afterward, we found that compared with the VD + DMSO group, the VD + rapamycin group had elevated levels of IgE, IL-4, and IL-5 in the serum of guinea pigs (all P < 0.05). The BALF cell counting analysis (Fig. 6D–H) found that compared with the VD + DMSO group, the VD + rapamycin group exhibited upregulated contents of total cells in BALF, eosinophils, neutrophils, and lymphocytes, and downregulated content of monocytes (all P < 0.05).

VD palliates airway inflammation in asthmatic guinea pigs by activating mTOR and suppressing autophagy.
In addition, results of H&E staining (Fig. 6I, J) unveiled that the VD + rapamycin group showed thickened bronchial smooth muscle, narrowed alveolar space, and increased inflammatory cell infiltration in bronchus, perivascular region, and alveolar space in guinea pigs compared with the VD + DMSO group, and the staining score in the VD + rapamycin group was higher than that in the VD + DMSO group (P < 0.01).
Eventually, levels of mTOR and autophagy cytokines (LC3B, Beclin1, Atg5, and p62) in lung tissues of guinea pigs in each group were examined by Western blot analysis (Fig. 6K, L), which showed that compared with those in the VD + DMSO group, levels of p-mTOR/mTOR and p62 were declined, and levels of autophagy cytokines LC3II/LC3I, Beclin1, and Atg5 were increased in lung tissues of the VD + rapamycin group. It was suggested that rapamycin reversed the improvement of airway inflammation in OVA-treated guinea pigs by VD, which in turn indicated that VD might reduce airway inflammation in asthmatic guinea pigs by activating mTOR to inhibit autophagy.
Discussion
Asthma referred to a refractory and heterogeneous disease, whose prediction and prognosis brought about unbearable burden to public medical system and resulted in surprisingly high death rate (Jones and others 2018). Oxidative stress generated from the process during inflammatory response in asthma was reversed by vitamin family through diet or supplement (Barrera-Mendoza and others 2018). VD was exhausted in serum of subjects with asthma, and it was responsible for frustrating prognosis, augmented asthma, encouraged airway inflammatory reactions, and damaged lung functions (Hall and Agrawal 2017). Therefore, we were triggered to explore the possible function of VD in the biological processes of asthma. Eventually, we discovered that VD could dampen airway inflammation in asthma by regulating autophagy.
The first critical finding from this experiment was that VD reduced levels of IgE, IL-4, and IL-5 in the serum of asthmatic guinea pigs. Highly expressed IgE in young asthmatic patients indicated impaired lung functions (Rajendra and others 2012). IgE was also associated with airway inflammation and poor pulmonary functions (Fairs and others 2010). Furthermore, when insufficient VD vitality occurred in asthmatic individuals, IgE level was upregulated and asthma was exacerbated (Manousaki and others 2017). On the other hand, release of IL-4 and IL-5 from adaptive and innate immune system was a remarkable sign of serious asthma (Robinson and others 2017). Liu and others (2017) unveiled that in a chronic allergic airway disease, decrease of IL-5 level was coupled with the reduction of inflammatory response in airway of lung, suggesting that IL-5 was detrimental for inflammation amelioration.
In patients with allergic bronchopulmonary aspergillosis, when VD level was promoted, IL-4 expression was reduced, indicating the reversed relationship between VD and IL-4 (Dodamani and others 2019). In that sense, VD could render conduve effect on asthmatic patients. Next, it was discovered that VD regulated cell counts in the BALF from asthmatic guinea pigs with the involvement of decreased numbers of eosinophils, neutrophils, and lymphocytes, and growing number of monocytes. As pivotal features in asthma, neutrophils discouraged therapy corticosteroid efficiency to asthma, while eosinophils and lymphocytes predicted complicated syndromes, airway remodeling, augmented inflammatory reactions, and airflow blockage (Amin 2016; Perlikos and others 2016; Bhalla and others 2018). Besides, supplement of monocytes tended to be a favored choice when it comes to the extensive recovery of pulmonary immunity functions after infection (Machiels and others 2017).
Previous advances suggested that VD quenched the expression of eosinophils, neutrophils, and lymphocytes in asthma and other immunity disorders (Vasiliou and others 2014; Cantorna and others 2015; Vanherwegen and others 2017). According to a recent assay developed by Carlberg, VD could mediate the performance of monocytes when it was challenged by antigens and immune cells, and thus to enforce the innate immune system (Carlberg 2019). From these evidences collected above, VD could ameliorate asthma by the regulation of wicked or benign cytokines in lung tissues and airway.
Interestingly, our data corroborated that VD reversed inflammatory cell infiltration in lung tissue from asthmatic guinea pigs. Inflammation impairs pulmonary peripheral and parenchyma airways, which can impose irreversible damage on airways (Barnes 2016). Suppression of the active inflammatory reaction in BALF and pulmonary tissues was a signature for an effective drug targeting asthma (Liang and others 2017). Since VD modulated the inflammatory response in asthma, it was related to limited inflammation, relieved symptoms, and benign prognosis (Hall and Agrawal 2017). In addition, VD enhanced mTOR expression and inhibited autophagy in lung tissues from asthmatic guinea pigs as evinced by declined levels of LC3B, Beclin1, and Atg5 and upregulated level of p62.
Multiple literatures indicated the principal role of autophagy in asthma as autophagy exacerbated immune systems, inflammatory response, mitochondrial damage, eosinophil secretion, and oxidative stress (Lv and others 2020; Silveira and others 2020). VD treatment prevented lung injury infected by Aspergillus fumigatus through the inactivation of autophagy (Hu and others 2020). Autophagy restricted by mTOR activation is an important part in reduced airway inflammation in allergic asthmatic mice (Wu and others 2020). When mTOR was exhausted in mice with airway inflammation, asthma was exacerbated, while mTOR activation was correlated with alleviated asthma (Zou and others 2018). Although the research about the relation between VD and mTOR in asthma is seldom found, VD was deemed as a promising treatment in diabetic cardiomyopathy as it ameliorated autophagy by regulating mTOR (Wei and others 2017). LC3B downregulation contributed to pulmonary damage and lung function in bronchopulmonary dysplasia (Wu and others 2019a).
Strong expression of Beclin1 and Atg5 was remarkable in individuals with asthma (McAlinden and others 2019). VD could decrease autophagy by downregulating the levels of LC3B and Beclin 1 in lupus nephritis (Yu and others 2019). p62 level was discouraged in individuals suffering from asthma, which coupled with severe inflammatory symptoms and autophagy (McAlinden and others 2019). As a prominent indicator of anti-autophagy, p62 content was downregulated in cells with enhanced autophagy and this downregulation was reversed by VD administration (Yu and others 2019). In other word, VD alleviated asthma by restricting inflammation and autophagy through mTOR activation.
In summary, our study supported that VD could dampen airway inflammation in asthma by inducing mTOR to restrict autophagy. These results discovered a novel approach for asthma treatment. Still, this is just a preclinical research; although our findings provide therapeutic implication in asthma treatment, the experiment results and effective application into clinical practice need further validation. However, there are still some limitations in our experiment, for example, we failed to figure out the specific mechanism of VD inhibiting autophagy in asthmatic guinea pigs by inducing mTOR. In the future, if possible, we will further explore other targets of VD. More attention will be paid on seeking reliable therapeutic targets of asthma.
Authors' Contribution
All authors contributed to data analysis, drafting, and revising the article, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Footnotes
Author Disclosure Statement
The researcher claims no conflict of interests.
Funding Information
The study was funded by Joint Fund project of Hubei Provincial Health Commission (Item No. WJ2019H218).