
Abstract
Metabolic abnormalities such as obesity, insulin resistance, and type 2 diabetes mellitus are known to be associated with adipose tissue inflammation and impaired secretion of cytokines. Anti-inflammatory cytokine interleukin-4 (IL-4) was found to promote insulin sensitivity, glucose tolerance, and reduce lipid accumulation in vivo through multiple mechanisms, including direct regulation of lipolysis in adipocytes. However, little is known about its role in adipocyte glucose metabolism. This study reveals that IL-4 upregulates glucose uptake in adipocytes without additional activation of the insulin-dependent IRS1 (insulin receptor substrate 1)-Akt (protein kinase B) pathway. Moreover, the main transcription factor STAT6 (signal transducer and activator of transcription 6), regulated by IL-4, was not involved in adipocyte glucose uptake. The proteomic results showed that IL-4 upregulates expression of proteins involved in mitochondrial biogenesis, renewal, and glucose oxidation. Our study provides a new hypothesis, explaining protective effects of IL-4 against metabolic abnormalities through activation of adipocytes glucose utilization and maintenance of mitochondrial function under metabolic overload conditions.
Introduction
Adipose tissue is one of the central players in whole body metabolic homeostasis due to its lipid-buffering role, endocrine function, and the capability for thermogenesis. In the course of obesity and overfeeding, adipose tissue receives and stores excessive amounts of lipids, which leads to metabolic overload and hypertrophy of adipocytes. Adipocytes metabolic stress is inextricably linked to the development of latent chronic inflammation and systemic insulin resistance (Shoelson and others 2006; Lackey and Olefsky 2016; Orliaguet and others 2020). In light of this, there remains intense interest in the reduction of the proinflammatory phenotype of adipose tissue.
Type II inflammatory cytokine interleukin-4 (IL-4) is known to be involved in shifting the adipose tissue phenotype to anti-inflammatory state. IL-4 is predominantly secreted by type 2 helper T lymphocytes (Th2), M2 macrophages, eosinophils, and basophils (Zhang and others 2015; Junttila 2018; Liu and Nikolajczyk 2019). In vivo IL-4 administration increases insulin sensitivity and glucose tolerance, decreases weight gain and lipid accumulation in the adipose tissue, and normalizes free fatty acids, cholesterol, and triglycerides in mice serum (Ricardo-Gonzalez and others 2010; Chang and others 2012; Lee and others 2017; Youngblood and others 2020).
Multiple studies confirm that the described effects are mediated by immune cells (M2 macrophages), which affect insulin sensitivity of adipocytes through secreted factors (Nguyen and others 2011; Wu and others 2011; Qiu and others 2014; Shan and others 2017). At the same time, IL-4 can influence adipocytes directly (Lee and others 2017; Stafeev and others 2018; Shiau and others 2019). IL-4 can activate signal transducer and activator of transcription 6 (STAT6) factor in adipocytes as well as in immune cells (Lee and others 2017). Furthermore, in some cell types, it activates mitogen-activated protein kinases and phosphatidyl-inositol-3 kinase (PI3K)–protein kinase B (Akt) pathways (Junttila 2018; Keegan and others 2018; Li and others 2019).
IL-4 reduces the capability of preadipocytes for adipogenic differentiation by the STAT6-dependent mechanism and stimulates lipolysis by activating protein kinase A and hormone-sensitive lipase (Tsao and others 2014; Shiau and others 2019). In addition, Th2 cytokines reduce the expression of perilipin and peroxisome proliferator-activated receptor (PPAR) in adipocytes in visceral fat depot (Wu and others 2011). Thus, IL-4 can lead to weight loss by reducing the formation of new adipocytes and increasing the degradation of already existing fat stores due to lipolysis.
Free fatty acids, released in response to IL-4, can be degraded during increased thermogenesis or locomotor activity. Th2 cytokines play an important role in the induction of beige adipocyte differentiation (Ricardo-Gonzalez and others 2010; Nguyen and others 2011; Shan and others 2017). Depletion of eosinophils, as well as knockout of IL-4, disrupts the biogenesis of beige fat during cold induction in mice (Qiu and others 2014). Furthermore, IL-4 may regulate secretory phenotype of adipocytes, hepatocytes, and myocytes. It stimulates GDF15 (growth differentiation factor 15) production through JAK (Janus kinase)-STAT6 pathway and reduces systemic inflammation and insulin resistance (Lee and others 2017).
The mentioned findings delineate the important role of IL-4 in adipocytes lipid metabolism, differentiation, and communication with other cells and organs. However, the role of IL-4 in glucose metabolism of adipocytes is still not well characterized. In this study we investigate the influence of IL-4/STAT6 axis on glucose transport of adipocytes and analyze proteomic changes for further addressing IL-4 involvement in regulation of glucose tolerance.
Materials and Methods
The experiments were approved by the Institutional Ethics Board (National Medical Research Center for Cardiology, permit #385.06.2009).
Cell culture
Mouse preadipocytes 3T3-L1 (American Type Culture Collection) were maintained in complete Dulbecco's modified Eagle medium (DMEM) with 4.5 g/L glucose (Gibco) supplemented with 10% FBS (HyClone) and 1% penicillin/streptomycin (Gibco). 3T3-L1 cells were differentiated into mature adipocytes according to Zebisch and others' (2012) protocol.
Preadipocytes were maintained in confluence for 2 days, followed by induction of adipogenesis with 100 nM insulin (Sigma-Aldrich), 0.5 mM 3-isobutyl-1-methylxanthine, 1 μM dexamethasone, and 2 μM rosiglitazone for 4 days and further subculturing in maintenance medium for another 4 days. Mature 3T3-L1 adipocytes were treated with 50 ng/mL mouse IL-4 (#I1020; Sigma-Aldrich) or transduced with lentiviral construct encoding mouse IL-4.
Plasmid construction and lentivirus production
Mouse IL-4 gene was synthesized in Evrogen (Russia) on the basis of mRNA sequence (GenBank, NM_021283). Mouse IL-4 sequence was cloned into lentiviral transfer vector LeGO-iG2 (#27341; Addgene) under spleen focus-forming virus promoter in Evrogen. Lentiviral transfer plasmid pGreenPuro (#SI505A-1; SBI) was used for shRNA expression.
Sequence of shSTAT6 top strand: GTTCAGATGCTTTCTGTTATTCAAGAGATAACAGAAAGCATCTGAACTTTTT; bottom strand: AAAAAGTTCAGATGCTTTCTGTTATCTCTTGAATAACAGAAAGCATCTGAAC; sh_scrambled top strand: ATCCGCGCGATAGTACGTATTTTCAAGAGAAATACGTACTATCGCGCGGATTTTT; bottom strand: AAAAATCCGCGCGATAGTACGTATTTCTCTTGAAAATACGTACTATCGCGCGGA. Oligonucleotides for top and bottom strands were synthesized in Evrogen, annealed and cloned in pGreenPuro according to SBI protocol with BamHI and EcoRI restriction enzymes.
Lentiviruses were produced in HEK293T (human embryonic kidney 293T line) cells transfected with transfer plasmid, envelope pMD2.G (#12249; Addgene), and packaging plasmids pMDLg/pRRE (#12251; Addgene), pRSV-Rev (#12253; Addgene), according to the protocol (Gándara and others 2018). After overnight calcium phosphate transfection, the medium was replaced with complete DMEM; virus was harvested in 24 h.
Lentiviral transduction of adipocytes
3T3-L1 adipocytes were transduced overnight with lentiviral stocks diluted 1:2 in complete DMEM, 8 μg/mL polybrene. The next day, medium was replaced with fresh culture medium and cells were incubated for 48 h before further experiments. Secretion of IL-4 in conditioned medium was detected using mouse IL-4 enzyme-linked immunosorbent assay (ELISA) kit (#M4000B; R&D Systems).
Western blot analysis
Adipocytes were lysed in radioimmunoprecipitation assay (RIPA) buffer (50 mM Tris-HCl, pH 8.0, 150 mM NaCl, 1% Triton X-100, 0.5% sodium deoxycholate, and 0.1% sodium lauryl sulfate). Samples were heated to 56°C in the Laemmli buffer, separated in SDS-PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis), and transferred to the polyvinylidene difluoride membrane (Laemmli 1970).
Membranes were blocked with 5% nonfat dried milk (AppliChem) and incubated with antibodies: pIRS1-Y612 (#44816; Thermo Scientific), pAkt-S473 (#4060; Cell Signaling), pAkt-T308 (#9275; Cell Signaling), insulin receptor substrate 1 (IRS1, #3407; Cell Signaling), Akt (#ab64148; Abcam, United Kingdom), pSTAT6-Y641 (#ab32520; Abcam), STAT6 (#ab44718; Abcam), GLUT1 (#ab652; Abcam), GLUT4 (#ab166704; Abcam), vinculin (#ab18058; Abcam), and β-actin (#ab6276; Abcam). Dilutions of all primary antibodies for Western blotting were 1:1,000. Appropriate horseradish peroxidase-conjugated secondary antibodies were purchased from Abcam (dilution 1:10,000). Proteins were visualized by enhanced chemiluminescence in Fusion FX gel-documentation system (Vilber-Lourmat, France) and quantified in GelAnalyzer2010a software.
[3H]-glucose radioassay
Mature adipocytes underwent deprivation in serum-free DMEM with 0.1% bovine serum albumin for 4 h and rinsed with DMEM with no glucose. The following operations were performed in DMEM with no glucose. Cells were stimulated with 100 nM insulin for 20 min and incubated in DMEM with 100 μM of 2-deoxyglucose (Sigma-Aldrich) and 0.1 μCi/mL [3H]-2-deoxyglucose (3H-2DG, #ART0103B; American Radiolabeled Chemicals) for 5 min. Cells were rinsed twice with cold phosphate-buffered saline and lysed in RIPA buffer. Samples were added into Beckman ReadySolv HP scintillation fluid (Beckman). The radioactivity was measured with a RackBeta counter (LKB Wallac, Sweden) and the results were protein normalized.
GLUT4 traffic analysis
Membrane translocation of insulin-dependent glucose transporter GLUT4 was analyzed using expression of myc-GLUT4-mCherry fusion protein. Plasmid for lentivirus assembling pLenti-myc-GLUT4-mCherry was a gift from Weiping Han (Addgene plasmid #64049; RRID: Addgene_64049). Analysis was performed as previously described (Lim and others 2015). In brief, 3T3-L1 adipocytes were transduced with myc-GLUT4-mCherry as described in Plasmid Construction and Lentivirus Production and Lentiviral Transduction of Adipocytes sections. Adipocytes were deprived from FBS for 4 h and treated with 100 nM insulin (positive control) and 50 ng/mL IL-4.
After that cells were washed and fixed in 4% paraformaldehyde in nonpermeabilized condition. Samples were blocked with 10% goat serum for 1 h at room temperature, rinsed with phosphate-buffered saline, and incubated with anti-myc antibodies (dilution for immunocytochemistry 1:100, #ab9106; Abcam) for 18 h at +4°C.
Samples were rinsed and incubated with Alexa Fluor488 antirabbit IgG (#A11034; Molecular Probes) for 1 h at room temperature. Cells mounted in Vectashield Antifade Mounting Medium with 4,6-diamidino-2-phenylindole (DAPI; Vector Laboratories) were subjected to confocal microscopy on Leica Stellaris 5 (Leica, Germany). Fluorescence of mCherry (excitation: 587 nm, emission: 593–792 nm) demonstrated distribution of all GLUT4 molecules and Alexa488 fluorescence (excitation: 499 nm, emission: 504–587 nm) marked membrane-exposed GLUT4.
Immunocytochemistry and confocal microscopy
Adipocytes were differentiated on glass coverslips (Thermo Scientific), fixed with 4% paraformaldehyde, and treated with 0.2% Triton X100 and Antigen Unmasking Solution (#H-3301; Vector Laboratories). Samples were blocked with 10% donkey serum for 1 h at room temperature, rinsed with phosphate-buffered saline, and incubated with STAT6 antibodies (dilution for immunocytochemistry 1:100, #ab32520; Abcam) for 18 h at +4°C. Samples were rinsed and incubated with Alexa Fluor594 antimouse IgG (#A21203; Molecular Probes) for 1 h at room temperature. Cells were applied in Vectashield Antifade Mounting Medium with DAPI (Vector Laboratories). Confocal microscopy was performed on Zeiss LSM780 microscope (Zeiss, Germany).
Proteome analysis
Differentially expressed proteins (DAPs) in adipocytes treated with recombinant IL-4 for 1 or 24 h were analyzed by shotgun mass spectrometry. Cells were lysed in denaturation buffer (5 mM urea, 15% acetonitrile, 0.5% sodium deoxycholate, and 5 mM trichloroethyl phosphate based on 300 mM sodium phosphate buffer with pH 6.0) in an ultrasonic bath (45°C, 30 min), after alkylation in 0.2% 4-vinylpyridine at 22°C for 30 min. Samples were trypsinized at 1/50 trypsin/substrate mass ratio for 2 h at 38°C; the reaction was stopped with formic acid.
Peptides were enriched on column C-18 m-PrecolumnPepmap (Thermo Scientific) and separated on column C-18 AcclaimPepmap (Thermo Scientific) in acetonitrile gradient. Mass spectrometry analysis was performed on hybrid orbital high-resolution mass spectrometer with linear ion trap OrbitrapFusion coupled with “UltiMate 3000 Binare RSLCnano” HPLC system (Thermo Scientific). A 3-segment differentiating scanning method with variable use of activation, isolation, and detector types was used.
Proteins were identified using “Search Gui 3.3” software (Compomics) by the X!Tandem Vengeance 12.15.2. search algorithm. The quantitative analysis was based on the NSAF index (normalized spectral abundance factor) in a set of proteins common to a group of samples after alignment by the intensity of the reference spectra. Gene ontology analysis was performed in PANTHER version 11 database (Mi and others 2017).
Statistics
For all parameters, 3 independent experiments were performed with 3 replicates. Statistical analysis was performed using GraphPad Prism 8.0. The results are presented as mean ± standard deviation. Statistically significant differences were evaluated by 2-way ANOVA with post hoc Tukey's test; the P values <0.05 were considered statistically significant.
Results
IL-4 activates glucose uptake in 3T3-L1 adipocytes
In this study, we aimed to more closely examine the role of IL-4 in adipocytes glucose metabolism. We produced 3T3-L1 adipocytes (Supplementary Fig. S1) expressing either mouse IL-4 or green fluorescent protein, as a control (mock group). Expression of IL-4 in transgenic adipocytes was verified by ELISA. IL-4 concentration in conditioned medium increased up to 70 ng/mL.
We investigated the regulation of glucose transport by IL-4 using measurement of basal and insulin-stimulated 3H-2DG uptake in adipocytes. Basal glucose uptake in IL-4 expressing adipocytes was higher than in control and mock groups (Fig. 1A). Insulin-stimulated glucose uptake was also increased by IL-4, but to the same extent as basal without potentiation of insulin effect. Furthermore, we sought to test whether IL-4 influences insulin signaling, which is the crucial regulator of glucose metabolism in adipocytes.
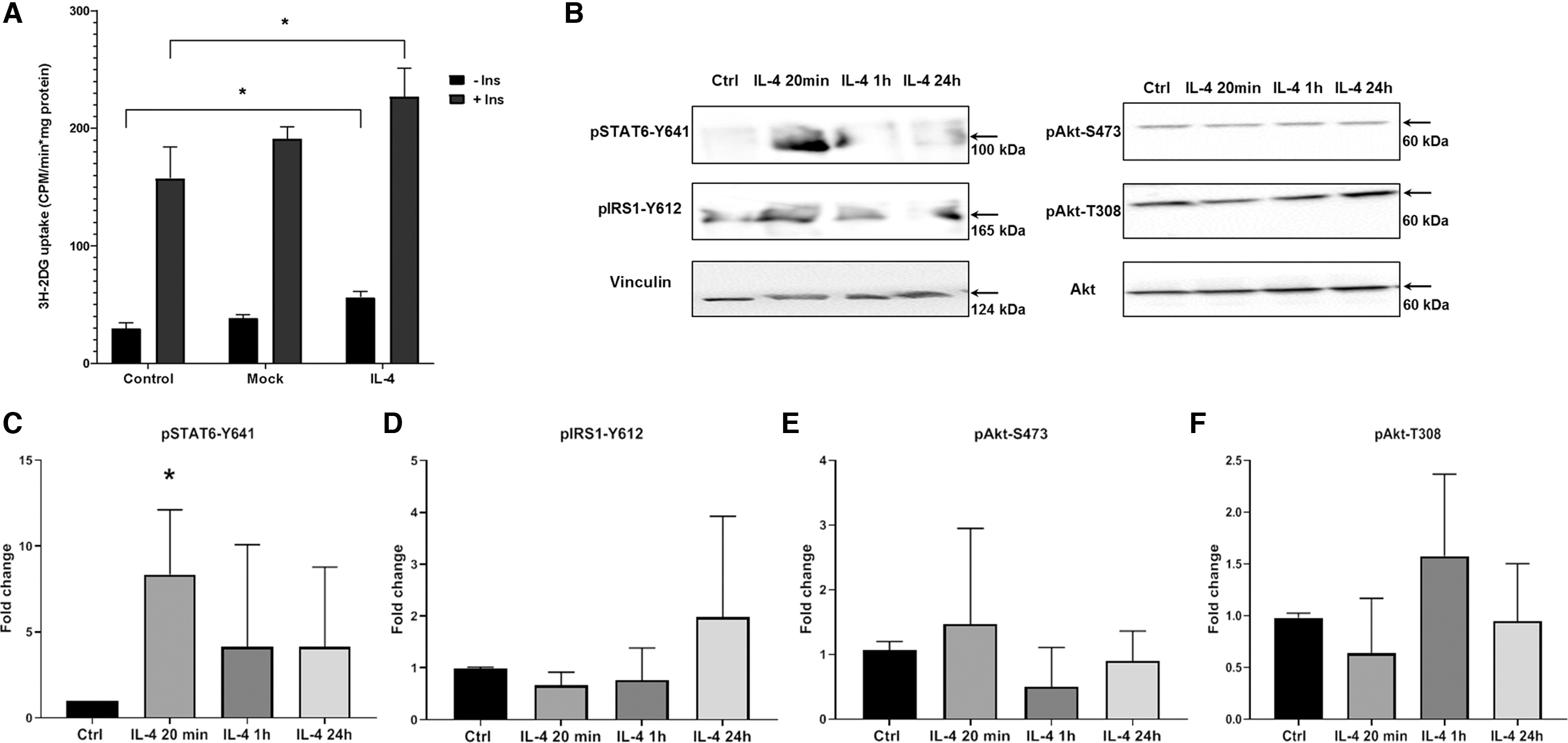
IL-4 activates glucose uptake with no effect on insulin signaling cascade in mature 3T3-L1 adipocytes.
To test time-dependent response of insulin signaling on IL-4, we treated adipocytes with recombinant IL-4 in concentration close to conditioned medium of transduced cells (Fig. 1B–F). Phosphorylation of IRS1-Y612, Akt-T308, and Akt-S473 was not affected at all time points examined, showing absence of IL-4 effect on insulin signaling. However, we observed activation of STAT6 phosphorylation of Y641, necessary for activation of transcription factor.
Glucose uptake activation can be the result of regulation of glucose transporters expression or translocation to the plasma membrane (Lim and others 2015). We examined whether IL-4 potentiates expression of main glucose transporters GLUT1 and GLUT4 (Fig. 2B–D) and exposure of GLUT4 on plasma membrane (Fig. 2A) in adipocytes. We found that IL-4 has no effect on GLUT1 and GLUT4 endogenous expression and does not increase appearance of GLUT4 in plasma membrane, analyzed by localization of myc-GLUT4-mCherry fusion protein. These results reveal a new role of IL-4 in adipocyte glucose uptake. However, this effect is not mediated through insulin signaling and GLUT1 and GLUT4 activation.

IL-4 has no effects on inducible glucose transporter GLUT4 traffic and on endogenous expression of GLUT1 and GLUT4.
IL-4 induces STAT6 phosphorylation and nuclear translocation in adipocytes
The STAT6 pathway is known to transmit signals from IL-4 receptor into the nucleus in myeloid cells. We, therefore, investigated whether STAT6 is activated by IL-4 in adipocytes. We first confirmed that STAT6 is phosphorylated at Y641 in 3T3-L1 adipocytes in response to IL-4 stimulation for 20 min and 1 h (Fig. 1B, C). Then, localization of STAT6 was analyzed by confocal microscopy (Fig. 3A). The accumulation of STAT6 in the nucleus was observed after 20 min incubation with IL-4 and decreased at 1 and 24 h. These data indicate that the effects of IL-4 on adipocyte metabolism may involve a mechanism dependent on STAT6.
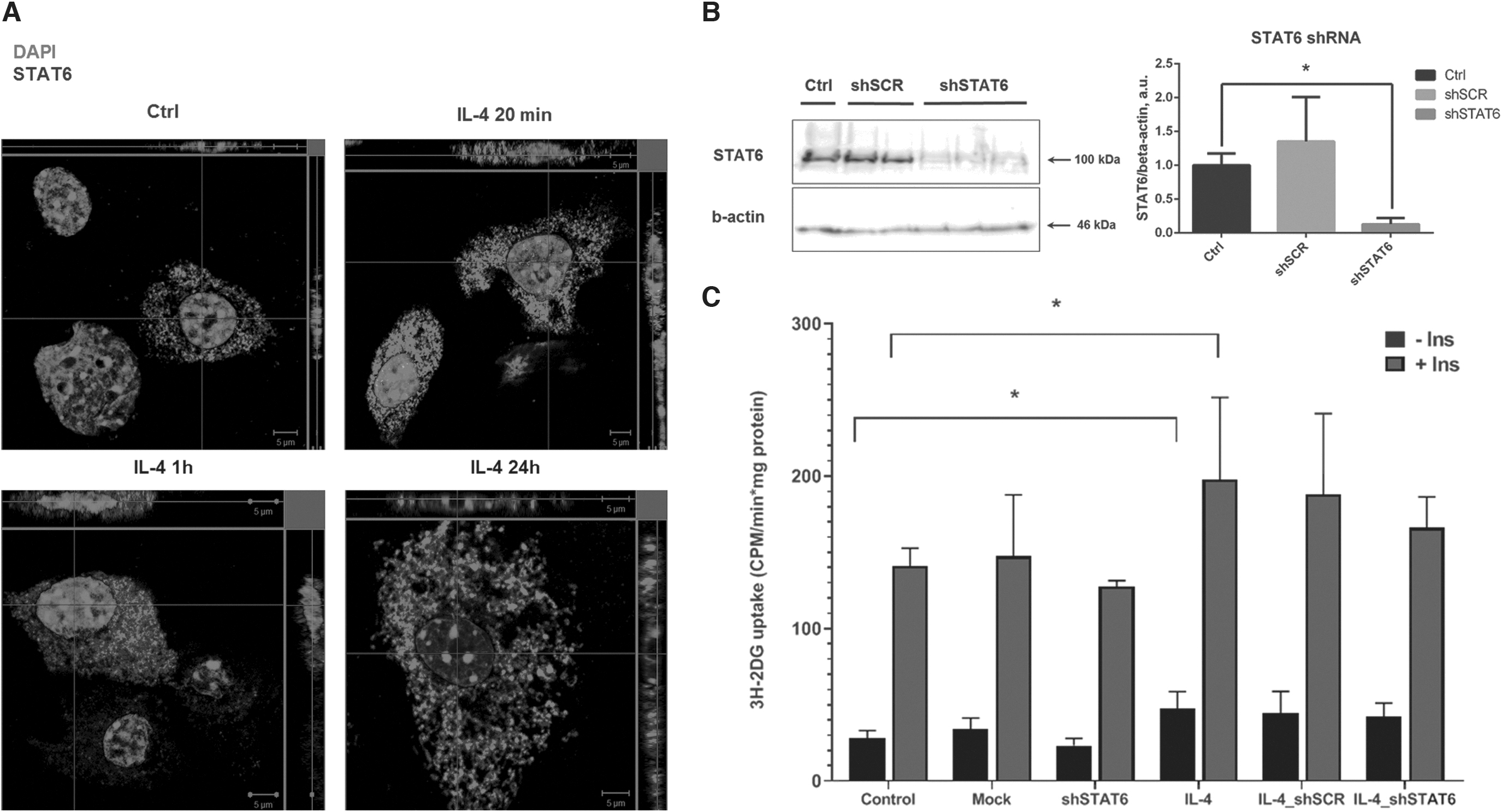
STAT6 translocates into nucleus under IL-4 but does not mediate IL-4 effect on glucose uptake.
To test this, we generated 3T3-L1 adipocytes with STAT6 knockdown using lentiviral constructs encoding shSTAT6. shRNA decreased STAT6 expression by 90% (Fig. 3B). Despite activation of STAT6 in adipocytes, we found that IL-4 is able to stimulate glucose uptake in absence of STAT6. Overall, our data show that STAT6 is dispensable for IL-4 effect on glucose uptake and suggest existence of other mechanisms.
IL-4 activates mitochondrial gene expression and antioxidant transcription factors in 3T3-L1 adipocytes
To provide insight into possible mechanisms of short- and long-term IL-4 action in adipocytes, we performed proteomic analysis of 3T3-L1 adipocytes treated with IL-4 for 1 h and for 24 h in 3 repeats. In each analysis on average, 610 DAPs were detected. We normalized results of each experiment to internal control and identified proteins up- or downregulated in all experiments (Fig. 4A). In total, IL-4 stimulation for 1 h induced differential expression of 83 proteins (50 upregulated, 33 downregulated); IL-4 for 24 h influenced 112 proteins (47 upregulated, 65 downregulated) (Supplementary Table S1).
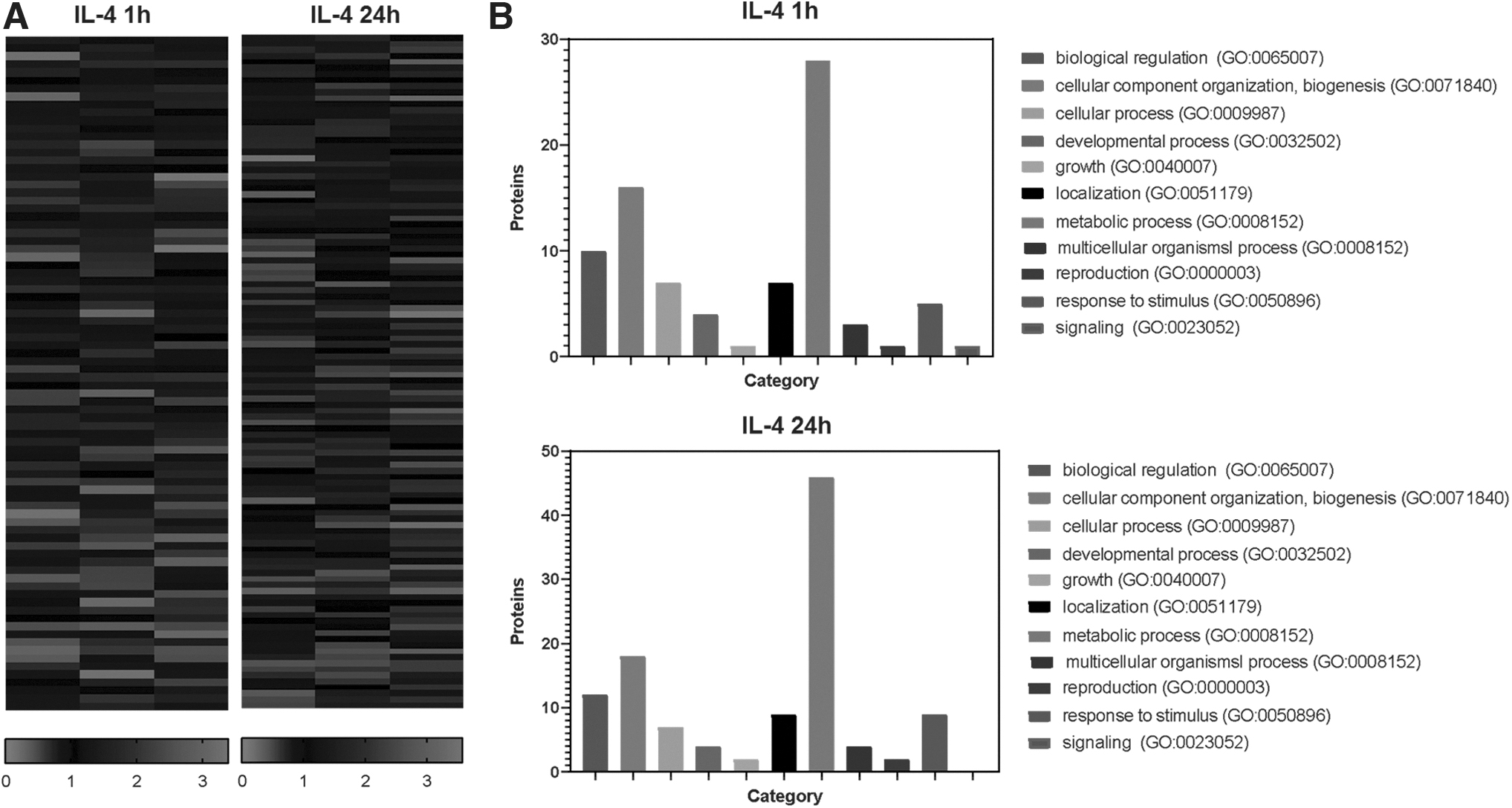
IL-4 regulates metabolic, mitochondrial, and oxidative response proteins expression in adipocytes.
To determine the potential functions of DAPs, they were annotated in the PANTHER database. The annotation indicated that DAPs could be classified into 11 classes (Fig. 4B). Gene ontology classes covered many important cellular processes, including metabolism, biological regulation, organelle organization, and biogenesis. For the DAPs detected after both 1 and 24 h of IL-4 treatment, the most enriched biological process was related to metabolism.
Furthermore, we highlighted proteins upregulated >50% under IL-4 (Supplementary Table S2) and demonstrated increased expression of mitochondrial proteins, such as succinate-CoA ligase (Q9Z2I9), ADP/ATP translocase 2 (P51881), citrate synthase (Q9CZU6), F-ATP-synthase subunit gamma (Q91VR2), protein NipSnap homolog (O55126), prohibitin (P67778), and FAM162A (family with sequence similarity 162 member A; Q9D6U8).
To suggest possible transcription factors involved in IL-4–mediated proteome changes, we analyzed DAPs in the TRANSFAC database. The results showed that transcription factors Nrf2 (nuclear factor erythroid 2-like 2), HIF1a (hypoxia inducible factor 2 alpha), GABPA (GA binding protein transcription factor subunit alpha), PPARa, and c-Jun regulate 3 or more DAPs in IL-4 for 24 h.
Discussion
Obesity and type 2 diabetes are associated with latent chronic inflammation in adipose tissue. Although the positive role of anti-inflammatory cytokine IL-4 in the regulation of systemic insulin sensitivity and glucose metabolism has been brought into sharp focus, relatively few are known about mechanisms of its action in adipocytes. In this study, we demonstrate the role of IL-4 in the regulation of adipocytes glucose uptake and analyze molecular mechanisms of the process.
According to previous studies, IL-4 is able to improve insulin sensitivity and glucose tolerance through activation of M2 macrophages and thermogenic adipocytes in adipose tissue, secretion of insulin-sensitizing cytokines, and downregulation of lipid storage in adipocytes (Qiu and others 2014; Lee and others 2017; Shiau and others 2019). Our results reveal that IL-4–expressing mature adipocytes take up more glucose in basal and insulin-stimulated states. This effect aligns with previously reported effects of IL-4 on adipocyte metabolism, because glucose uptake can precede thermogenesis or lipid turnover.
Activation of glucose transport is often associated with increased expression of dominant glucose transporters (GLUT1 and GLUT4) or their plasma membrane translocation (especially, insulin-responsive GLUT4). However, in our study, IL-4 has no effect on GLUT1 and GLUT4 content and localization. We suggest that glucose transport under IL-4 stimulation can be realized through other members of GLUT family, expressed in adipocytes, such as GLUT3, GLUT8, GLUT10, GLUT11, and GLUT12 (Chadt and Al-Hasani 2020).
First, we hypothesized that IL-4 stimulates glucose uptake through the IRS1 phosphorylation and PI3K-Akt signaling that is the main insulin-dependent pathway for stimulation of glucose transport in adipocytes. Moreover, the IRS1-PI3K pathway was found to be activated by IL-4 in immune cells (Keegan and others 2018). We found that insulin signaling (phosphorylation of IRS1-Y612 and Akt-T308, S473) is not induced by IL-4 in mature adipocytes. It indicates that increased glucose uptake is not the result of IL-4 capacity to promote IRS1 and PI3K signaling.
This observation is consistent with the findings of previous studies on hepatocytes and myocytes showing that IL-4 activates basal glucose uptake without Akt stimulation (Yang and others 2018; Chang and others 2019). However, some studies report contradictious effects (Chang and others 2012). In addition to IRS1, IL-4 is able to activate IRS2 and following PI3K-Akt pathway in immune cells. However, whether IRS2 is mostly a positive or a negative regulator of PI3K remains debatable (Keegan and others 2018). A potential contribution of IRS2 in adipocyte metabolism should be investigated in further studies.
The role of STAT6 transcription factor in IL-4 effects in nonimmune cells was described in the context of adipocyte differentiation, secretion of cytokines, and liver oxidative metabolism (Ricardo-Gonzalez and others 2010; Chang and others 2012; Lee and others 2017). Regarding adipogenic differentiation, controversial results were reported. Tsao and others (2014) declare STAT6-dependent inhibition of adipogenic differentiation. In contrast, Deng and others (2000) demonstrate STAT6-independent effects. Our results suggest that IL-4 can transiently activate STAT6 in mature adipocytes. However, it is not essential for IL-4–dependent upregulation of glucose uptake.
To get on the trail of possible mechanisms of glucose uptake upregulation by IL-4, we analyzed transient and prolonged changes in proteome under IL-4. Both transient and prolonged IL-4 action results in regulation of proteins involved in metabolic processes. Transient IL-4 treatment resulted in upregulation of proteins involved in glycolysis (phosphoglycerate kinase 1), tricarboxylic acid (TCA) cycle (succinate dehydrogenase and isocitrate dehydrogenase), and pentose phosphate pathway (6-phosphogluconate dehydrogenase and transaldolase). Prolonged IL-4 action also influenced catabolic enzymes (glyceraldehyde-3-phosphate dehydrogenase, acyl-CoA dehydrogenase, citrate synthase, and others) and ATP-synthase subunits.
However, under prolonged IL-4, some glycolytic and TCA cycle enzymes decreased, possibly reflecting desensitization or compensatory mechanisms. This study is supported by previous findings of omics studies reporting regulation of catabolic processes under IL-4 in adipocytes and mitochondrial proteins in lymphocytes (Shiau and others 2015; Ruiz-Lafuente and others 2018). Among the most strongly regulated proteins, we also found proteins involved in mitochondrial quality control, renewal, and apoptosis (FAM162A, NipSnap, prohibitin) (Wei and others 2017; Dao Trong and others 2018; Abudu and others 2019).
Furthermore, TRANSFAC analysis revealed that detected proteins could be regulated by transcription factors involved in autophagy, antioxidant defense, mitochondrial oxidation (Nrf2, PPARa), and in cytokine expression and uncoupling (c-Jun) (Yang and others 2014; Kovac and others 2015; Brocker and others 2018; Xiao and others 2019). Collectively, IL-4 is a potential activator of glucose oxidation in glycolysis and TCA, resulting in enhanced glucose uptake by adipocytes. Furthermore, IL-4 can possibly support high rates of oxidation keeping mitochondria healthy.
Finally, together with previous studies, our study provides insights into the regulation of metabolism in insulin-dependent tissues by immune mediators. During chronic nutritional overload and obesity, activation of low-grade inflammation in adipose tissue impairs metabolism, mitochondrial function of metabolically active cells, and downregulates insulin sensitivity. Secretion of anti-inflammatory cytokine IL-4 is able to improve systemic metabolism due to activation of adipocytes glucose transport, mitochondrial oxidation, and prevention of mitochondrial damage during metabolic stress.
Limitations
This study was performed only on 1 mouse cell line 3T3-L1, differentiated in adipocytes. IL-4 role in adipocytes glucose metabolism should be further evaluated in primary mouse and human cell cultures and in vivo. The effects of IL-4 on glucose uptake were analyzed in cells, expressing IL-4, but effects on cell signaling were analyzed under recombinant IL-4 stimulation for detection of time-dependent changes of phosphorylation that cannot be accurately measured in IL-4–expressing cells.
Conclusion
Our study made a contribution to understanding the crosstalk between immunity and metabolism by showing that IL-4 regulates adipocytes glucose uptake. Moreover, the action of IL-4 in adipocytes is not mediated through classical mechanisms of STAT6 or IRS1 activation reported for immune cells. Activation of glucose uptake likely results from IL-4 action on mitochondrial metabolism, biogenesis, and renewal. Finally, our study elaborates previous studies of IL-4 metabolic action and provides background for further investigation of IL-4 mechanisms in nonimmune cells.
Footnotes
Authors' Contributions
S.M. and I.S. designed the study, collected, analyzed, and interpreted the data, and wrote and reviewed the article. I.B. and E.Z. contributed to confocal microscopy and lentivirus construction techniques. E.M. collected and analyzed the data. A.K. performed mass spectrometry analysis. E.S. contributed to lentiviral construction design. M.M. reviewed the article. Y.P supervised the study and reviewed the article. All authors have read and approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by Russian Foundation of Basic Research Grant No. 20-015-00100. Mass spectrometry analysis was performed on the equipment of the “Human Proteome” Common Use Centre supported by the Ministry of Education and Science of the Russian Federation (Agreement No. 14.621.21.0017; ID RFMEFI62117X0017).
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2