
Abstract
Interleukin-7 (IL-7) is a cytokine belonging to the chemokine family. It plays a key role in the differentiation, development, and maturation of T lymphocytes and B lymphocytes, which is pivotal to adaptive immunity. In addition to its role in lymphocyte development, recent studies have indicated the antitumor functions of IL-7 in the tumor microenvironment. In this review, we discuss the role of IL-7 in tumors and summarize its antitumor potential and clinical application in lymphoma, leukemia, breast cancer, colon cancer, and so on. Furthermore, the combinational strategies of IL-7 and other antitumor drugs have been also discussed.
Introduction
The Interleukin 7 (IL-7) is a member of the IL-2 superfamily with a 4-alpha-helix bundle structure. It is mainly produced by bone-marrow and thymic stromal cells and functions as a hematopoietic growth factor (Fiorentino and others 1989). IL-7 was first discovered by Hunt and others in 1987 as a secreted factor that independently maintained the growth of B cell precursors from long-term bone marrow culture. In 1988, Namen and others purified a protein with this activity and cloned the gene for it. The protein was first named lymphopoietin-1, and then named IL-7 (Lee and others 1988).
IL-7 was determined to be a 25 kDa soluble secretory globular protein encoded by the IL-7 gene. The IL-7 receptor (IL-7R) is a heterodimeric complex composed of 2 chains, IL-7R subunit alpha chain (IL-7Rα, CD127) and common gamma chain (γC, CD133). The IL-7Rα is unique to the IL-7R, whereas γC is shared by several other cytokines, including IL-2, IL-4, IL-9, IL-15, and IL-21 (Greene and others 1986). The specific binding of IL-7 and IL-7Rα leads to dimerization of IL-7Rα and γC. The γC-associated Janus Kinase-3 (JAK3) and IL-7Rα-associated JAK1 are brought together, activated by transphosphorylation, and further phosphorylate the tyrosine residues in the cytoplasmic portions of IL-7Rα (Baird and others 1998).
The phosphorylated IL-7Rα acts as an anchored site for recruiting other signal molecules to form a complex in which they are phosphorylated and activated, including STAT5, PI3 kinase (PI3K), and Bcl2 protein (Benbernou and others 2000). As for the recruited transcription factor STAT, it is phosphorylated at the tyrosine residues by activated JAKs, and then it enters nucleus as a dimer and binds to the target gene STAT5a to regulate the transcription of downstream genes (Villarino and others 2015). Similarly, PI3K is brought close to the membrane as the p85 regulatory subunit of PI3K binds to the phosphorylated tyrosine residues on IL-7Rα.
This activates the p110 subunit of PI3K, which converts the substrate PIP2 into PIP3 (Yu and Cui 2016). Akt is recruited from the cytoplasm to the cell membrane through the binding of PIP3 to the N-terminal PH domain of Akt. And with the help of inositol 3-phosphate-dependent protein kinase 1 (PDK1) and 3-inositol 3-phosphate-dependent protein kinase 2 (PDK2), the threonine phosphorylation sites (Thr308) and the phosphorylation sites on Akt protein are phosphorylated to activate it (Asati and others 2016; Yu and Cui 2016). The signal pathway associated with IL-7 is shown in Fig. 1.
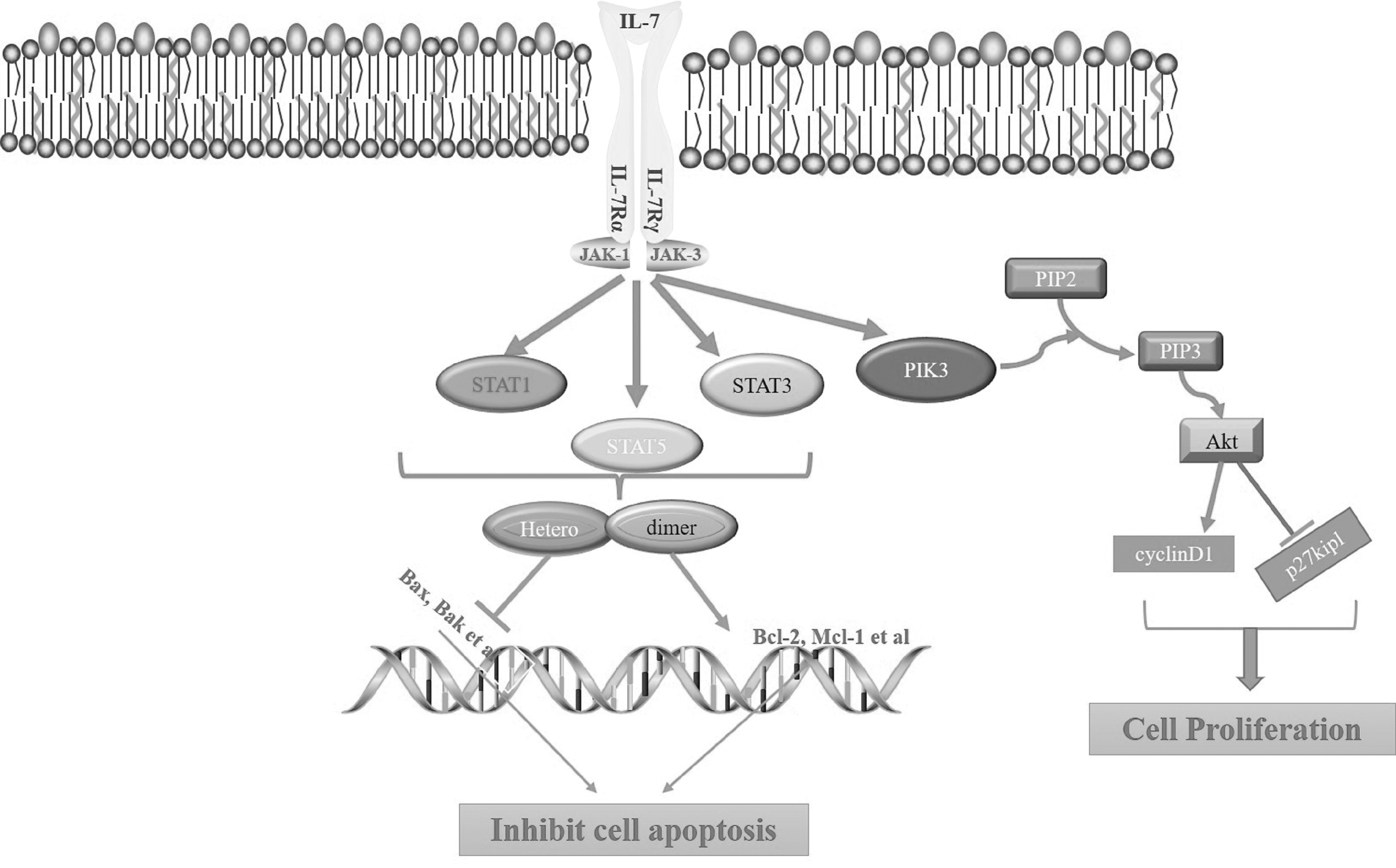
IL-7 signaling pathway. IL-7 specifically binds to the receptor to form a trimer composed of IL-7, IL-7Rα, and IL-7γc. The IL-7α in the activated trimer complex acts as an anchor site for recruiting other signal molecules to form a complex where they are phosphorylated and activated. SATA5 activated by phosphorylation regulates the transcription pathway to promote the expression of Bcl2 protein and inhibit the expression of BAX protein. Under these 2 actions, cell apoptosis is inhibited. PI3K activated by phosphorylation activates the Akt signaling pathway. Akt signal promotes cell proliferation by promoting cyclinD1 and inhibiting p27kip1. IL-7, interleukin-7; IL-7Rα, interleukin-7 receptor alpha; IL-7γc, interleukin-7 gamma chain.
The Role of IL-7 in Development of Lymphocytes
The first biological function of IL-7 is discovered in the development of T lymphocytes and B lymphocytes (Bazdar and others 2015). B cells are derived from the differentiation and development of hematopoietic stem cells from bone marrow. Previous studies indicated that the role of IL-7 is not directly affected by IL-7 itself in the development of B cells, but more likely there is a certain connection between IL-7 and pre-B cell receptors during the survival, proliferation, and differentiation of B cells (Fisher and others 1995; Valenzona and others 1996; Mertsching and others 1998).
This connection is that IL-7-induced proliferation of B cell progenitor cells depends on IL-7Rα chain tyrosine residues. The role of the α chain on the differentiation of B lymphocytes cannot be replaced by the IL-2Rβ chain (Corcoran and others 1996). However, current research results show that IL-7 is necessary for the development of B cells in mouse models but not necessary for humans (Puel and others 1998; Kohn and others 2012). Through administration of IL-7 to normal mice and lymphocyte-deficient mice, the result showed an increase of mature B cells in the mice (Damia and others 1992; Kohn and others 2012).
T cells are derived from lymphoid stem cells in the bone marrow and undergo differentiation and maturation in the thymus. After lymphoid progenitor cells migrate from the bone marrow to the thymus, many lymphocyte lineages are produced in the thymus, where the development of T lymphocyte lineages depends on or is affected by IL-7 (Vosshenrich and others 2006; Kim and others 2021). The expression of IL-7R regulates the development and proliferation of T cells in the thymus. When IL-7R signaling was blocked, T cells differentiated into CD4+ T cells. On the contrary, IL-7R signaling can differentiate into CD8+T cells (Issuree and others 2017). When IL-7 is used to stimulate fresh T cells, T cells will proliferate in a dose-dependent manner; knocking out IL-7R will stop the proliferation of T cells.
CD4+ T cells and CD8+ T cells have different response effects to IL-7, and IL-7 has a stronger effect on CD8+ T cell subsets than on CD4+ T cells (Welch and others 1989; Geiselhart and others 2001).
The Tumor Inhibitory Role of IL-7
Recent researches have shown that IL-7 and IL-7R have a certain inhibitory effect on chronic inflammation, autoimmune diseases, and blood diseases (Conlon and others 2019). Especially, IL-7 has significant antitumor effects in various tumors, including glioma, melanoma, lymphoma, leukemia, prostate cancer, and glioblastoma (Ye and others 2001).
In the mouse model, the xenografted tumor growth was significantly inhibited after administration of IL-7 in vivo, and even some individual xenografted tumor experienced complete tumor regression (Ye and others 2001). The administration of IL-7 to the rat brain glioma model can effectively inhibit tumor growth and enhance the antitumor effect of INF-γ (Fritzell and others 2013). In another study, after the administration of IL-7 in fibrosarcoma tumor-bearing mice, the tumor growth was significantly inhibited and the survival of the mice was prolonged. After IL-7 treatment, the proportion of CD4+ and CD8+ T cells in the tumor site, lymph nodes, and spleen increased significantly, and lymphocyte infiltration in the tumor microenvironment increased (Tang and others 2014).
Besides, IL-7 can enhance the survival and homeostatic proliferation of CD8+ T cells (Hashimoto and others 2019). In 1 study, by enhancing IL-7 signal, CD8 effector T cells increased, and tumor antigen-specific CD8+ T cell response was enhanced.(Sun and others 2006; Colombetti and others 2009). However, the number of memory CD8+ T cells did not increase, indicating that IL-7 is not necessary for the formation of memory CD8+ T cells, but after the formation of memory CD8+ T cells, IL-7 can maintain cell homeostasis to achieve tumor-specific cells answer (Klonowski and others 2006; Hand and others 2007). The aforementioned results have shown that IL-7 has good antitumor effects in a variety of tumor models, and these effects are closely related to the antitumor immunity activated by IL-7.
IL-7 and Antitumor Immunity
In addition to promoting the development and maturation of lymphocytes, IL-7 exhibits potent antitumor properties. When T lymphocytes recognize tumor-specific antigens and become activated in the tumor microenvironment, they play a significant role in the antitumor immune response. Thus, researchers are trying to upregulate the expression of IL-7 in the tumor microenvironment, because IL-7 can promote CD8+ T cell maturation and recruit CD8+ T cells to tumor tissues to act on tumor cells. To achieve the satisfactory antitumor immunity, many methods have been adopted (Fig. 2).
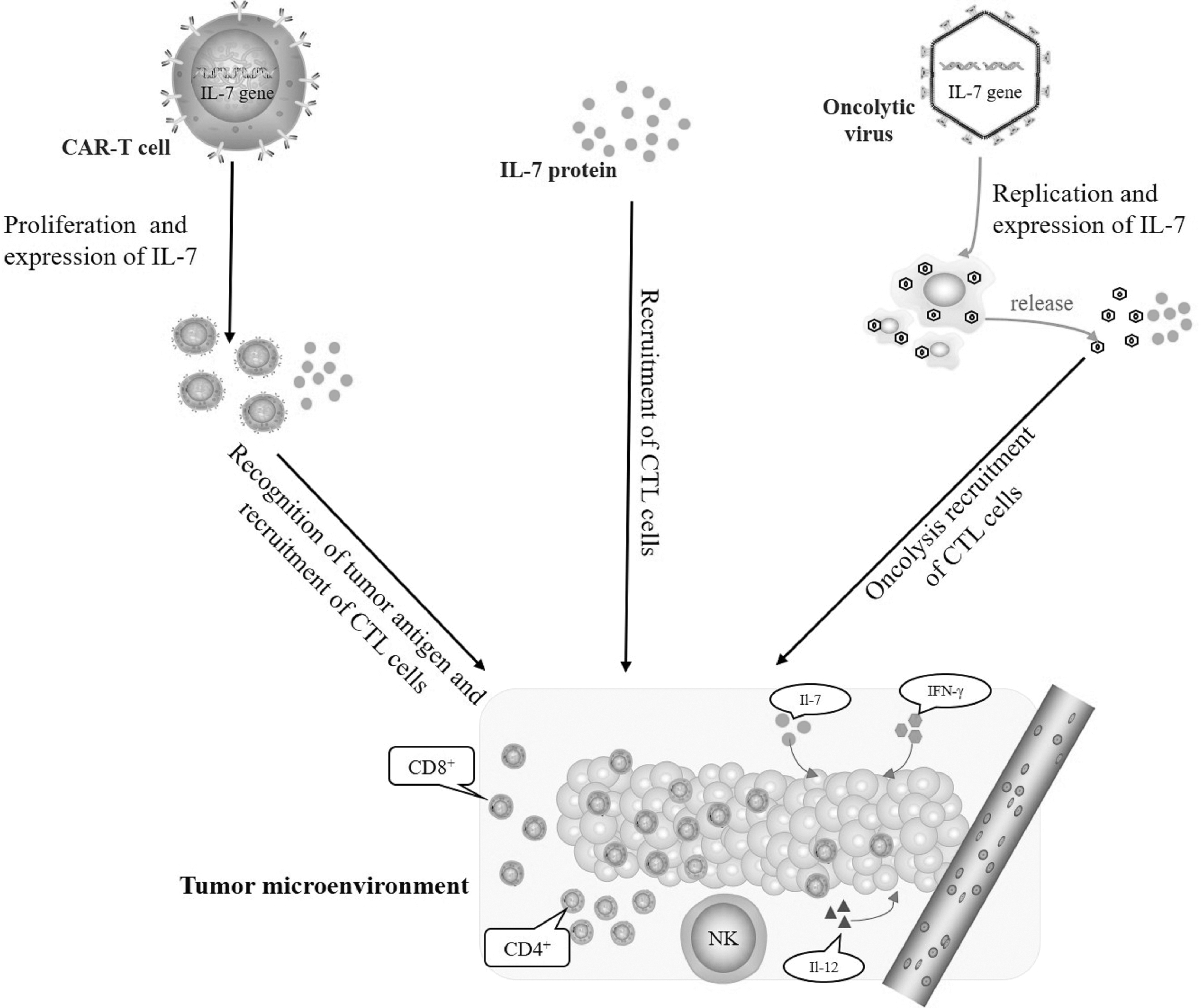
Strategies of antitumor immunity mediated by IL-7. After targeting tumor, the virus and CAR-T cells play a role in killing tumor cells, and promote the tumor microenvironment to produce IFN-γ, IL-12, and other cytokines. The expression upregulation of IL-7 promotes the survival and proliferation of CD8+ T cells, and enhances the antitumor immune effect.
Recombinant IL-7 treatment
Owing to its role in recruiting T cells and promoting antitumor immunity, direct injection of recombinant IL-7 proteins is used to treat malignant tumors. In mice in vivo experiments, the researchers injected hIL-7 recombinant protein into the tumors of malignant melanoma and colon cancer mouse models. hIL-7 has a significant inhibitory effect on tumor growth and metastasis. Antitumor activity is related to a significant increase in the number of tumor-infiltrating CD4+ CD8+ T cells and activated dendritic cells (DCs) (Lai and others 2011; Han and others 2017). The combination of IL-7 and granulocyte-macrophage colony stimulating factor (GM-CSF) enhances the antitumor response of a GM-CSF-secreting tumor cell immunotherapy, which is mainly achieved by the increase of activated DC and T cells in lymphoid tissues (Li and others 2007).
In a mouse lymphoma model, supplementation with exogenous recombinant IL-7 can significantly expand and maintain CD4+ effector cells and improve the therapeutic effect of lymphoma (Ding and others 2017). The treatment of IL-7 and INF-γ combined with α-CTLA-4/α-PD-1 therapy also showed a better tumor suppression effect in mouse bladder cancer models than that of single treatment (Shi and others 2016), this effect is mainly achieved by reversing immunosuppression by IL-7/IL-7R. The aforementioned studies suggest that in addition to its role in increasing lymphocyte proliferation, IL-7 can also be used as an adjuvant co-administered with GM-CSF, IFN-γ, or immune checkpoint inhibitors α-CTLA-4 and α-PD-1 to enhance the antitumor effect of these drugs.
Combinational therapy of IL-7 and chimeric antigen receptor-T
Chimeric antigen receptor (CAR)-T cells are artificially added (chimeric) receptors for specific antigens to normal T cells, so that T cells can perform effective killing functions in tumor cells expressing specific antigens (June and Sadelain 2018). Currently, CAR-T cells are mainly used to treat hematological malignancies, including acute lymphocytic leukemia, chronic lymphocytic leukemia, lymphoma, and multiple myeloma (Zhao and others 2018). Two CD 19-specific CAR-T cell therapies have shown significant efficacy in patients with relapsed/refractory B cell carcinoma, and were approved by the U.S. Food and Drug Administration (FDA) in 2017 (Lee and Kim 2019).
In June 2021, Fosun Kate's CD19-specific CAR-T cell therapy product Akirensai injection (FKC876, Yescarta) for the treatment of adult relapsed or refractory large B cell lymphoma was approved for marketing in China (National Medicine Standard S20210019). In recent years, CAR-T therapies are broadly used in various solid tumors, and have made great progress in clinical trials. Inspired innovative CAR design coupled with improvements in gene transfer technology, combined with advances in gene editing, may improve their effectiveness in solid tumors.
Zhou and others (2019) found the number of CAR-T cells are significantly increased, and its antitumor activity is also significantly improved when IL-7/IL-15 and CAR-T are used in combination. It suggests that IL-7 may play an important role in the proliferation of CAR-T cells. Recently, Adachi and others designed a CAR-T cell that can express IL-7 and CCL19 (7 × 19 CAR-T cells). In vivo, the 7 × 19 CAR-T cells induce the infiltration of DCs and immune cells such as CD4+ and CD8+ T cells into tumor microenvironment. And it achieved complete regression of pre-established solid tumors and prolonged mouse survival.
It also enhances the survival of CAR-T cells in tumor microenvironment. CAR-T cells and recipient immune cells collaborated to exert antitumor activity. Thus, compared with conventional CAR-T cells, 7 × 19 CAR-T cells have superior antitumor activity (Adachi and others 2018). Zhao and others used constitutively activated IL-7 receptor (C7R) to enhance the antitumor function of the constructed CAR-T cells. In vitro co-incubation experiments with target cells found that co-expressed C7R can significantly improve the activation, proliferation, and killing effects of CAR-T cells. In vivo experiments show that enhanced CAR-T cells have obvious antitumor activity in the TNBC (triple-negative breast cancer) subcutaneous xenograft model, and can prolong the survival time of CAR-T cells in vivo (Zhao and others 2020).
He and others constructed a novel CAR-T cells expressing IL-7 based on NKG2D to treat prostate cancer. By co-expression with IL-7, NKG2D-CAR T cells have a better antitumor effect and can more effectively inhibit the growth of xenograft tumors (He and others 2020; Zhang and others 2021; Borst and others 2022). In the system of CAR-T cells-mediated co-expression of IL-7, IL-7 regulates the increase of Bcl2 expression and inhibits cell apoptosis. IL-7 can also enhance the proliferation of CD8+ T cells, leading to the continued survival of CAR-T cells and the maintenance of antitumor vitality (Vella and others 1998; Fluxá and others 2018). The aforementioned research results demonstrate that CAR-T cell therapy combined with IL-7 expression in the treatment of solid tumors has a good antitumor immune effect and a good clinical application prospect.
IL-7 arming oncolytic virotherapy
Oncolytic viruses (OVs) are viruses that selectively infect and lyse tumor cells. The mechanism of action is to directly lyse cells and induce tumor immunity (Jhawar and others 2017). In the process of virus replication and cell lysis, the virus will also reinfect neighboring cells to amplify the response (Mullen and Tanabe 2002). Lysed tumor cells can also enhance antigen presentation and immune activation by releasing tumor-associated antigens, cytokines, and damage associated molecular patterns (Kaufman and others 2015). At present, >10 viruses have been used to develop OVs.
The 4 most widely used viruses are adenovirus (31%), herpes virus (24%), reovirus (20%), and vaccinia virus (12.5%). Other researches include Newcastle disease virus, Coxsackie, measles virus, and so on (Goradel and others 2021). A total of 3 types of OVs have been approved for marketing in the world, namely Rigvir (2004/Latvia) for the treatment of melanoma (Doniņa and others 2015; Alberts and others 2016), Ocorine (H101) for the treatment of head and neck squamous cell carcinoma (2005/China), and T-VEC for the treatment of melanoma (2015/FDA) (Aurelian 2016).
Increasing studies have attempted to use OVs carrying cytokine genes to enhance the oncolytic effect and antitumor immune effect. In the research of treating glioblastoma, Huang and others (2021) constructed an oncolytic adenovirus loaded with IL-7. The combination of oAD-IL-7 and CAR-T can promote T cell proliferation and reduce T cell apoptosis. In in vivo experiments with tumor-bearing xenogeneic mice as experimental subjects, both oAD-IL-7 and CAR-T can prolong the survival time of mice and reduce the burden of tumors. oAD-IL-7 can promote the proliferation and persistence of tumor-infiltrating CAR-T.
Nakao and others constructed an oncolytic vaccinia virus expressing IL-7 and IL-12 (hIL-7-VV and mIL-12-VV). Intratumoral injection of the virus into melanoma and colon cancer-bearing immunocompetent mice can activate the immune state, leading to complete tumor regression. In tumor models that do not respond to checkpoint inhibitor monotherapy, the combination of virotherapy and antiprogrammed cell death-1 (PD-1) or anticytotoxic T lymphocyte antigen 4 (CTLA 4) antibodies further improves antitumor activity. Administration with vaccinia virus carrying IL-7 and IL-12 genes can make tumors sensitive to immune checkpoint blockade through changes in immune status (Nakao and others 2020).
CD8+ T cells are one of the important cell types, which directly or indirectly exert antitumor effects by releasing cytolytic molecules and inflammatory cytokines in the tumor microenvironment. IL-7 alone can increase the diversity of CD8+ T cells in tumors, and this effect is significantly enhanced in the presence of IL-12 (Tasaki and others 2021). By studying the phenotypic changes of CD8+ T cells in the tumor, it was found that the expression of inducible costimulatory factor (ICOS) increased in PD-1-CD8+ T cells. Unlike previously reported ICOS+ CD8+ T cells, a subset of ICOS+ PD-1-CD8+ T cells exhibited effector functions characterized by granzyme B expression. This discovery reveals a potential direction of the oncolytic vaccinia virus treatment mechanism (Yamashita and others 2021).
Recently we constructed an oncolytic vaccinia virus capable of expressing IL-7. The results of in vitro studies show that the high expression of IL-7 has no effect on virus replication. The expression of IL-7 improves the killing effect of the virus on tumor cells and induces high expression of cytokines such as TNF-α and IFN-γ (unpublished data). Further in vivo animal studies are ongoing. The aforementioned results show that the oncolytic virus combined with IL-7 expression in the treatment of malignant tumors has a good tumor killing and antitumor immune effect, indicating a promising clinical application.
Clinical application of IL-7-mediated tumor therapy
IL-7 has exerted great antitumor potential both in in vitro and animal experiments. Therefore, it suggests that IL-7 has good application prospects in clinical trials (Table 1). Xu and others (2014) found that the combined effect of IL-7 and IL-15 is crucial in increasing the number of CD8+ CD45RA+ CCR7+ in CAR-T cell products and enhancing its persistence and antitumor activity in preclinical models. This combination can improve the therapeutic benefit of CAR-T therapy in the clinical trials (NCT00586391 and NCT00709033). O Trédan and others (2015) conducted a study on the optimal treatment time of a recombinant IL-7 (CYT107) for the expansion of lymphocyte pools in patients with metastatic breast cancer. The results demonstrate that CYT107 can expand the CD4+ T cell pool, but the optimal treatment time needs further research to explore (NCT01362107). The detailed clinical trials for tumor therapy related to IL-7 are listed in Table 1.
Clinical Trials of Interleukin-7 Mediated Tumor Therapy
IL-7, interleukin-7.
Conclusion and Perspectives
IL-7 plays an active role in activating antitumor immunity, which is mainly achieved by enhancing the proliferation and survival of T lymphocytes. IL-7 improves the antitumor efficacy of CAR-T cell therapy through enhancing the activation of T lymphocytes. Thus, chemokine-based immunotherapy plays a major role in the treatment of advanced malignant tumors such as melanoma, colon cancer, and triple-negative breast cancer. It is necessary for us to expand our understanding of the role of IL-7 in antitumor mechanisms. It has been suggested the signaling pathways downstream of IL-7R, JAK\STAT pathway and PI3K\AKT\mTOR pathway, might play an important role in the antitumor effect of IL-7.
Further research is needed to determine the mechanism of action. On the contrary, some studies have pointed out that IL-7 has a pro-tumor role in the development and progression of tumors, but this view is very controversial. Regarding the contradictory evidence presented in the literature, we speculate that the distribution and expression of IL-7 in physiological locations may be different in tumor progression. Besides, further research will undoubtedly focus on exploring the molecular mechanism of IL-7 in antitumor immune response, and developing promising and effective antitumor immunotherapy based on IL-7.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Public Welfare Technology Project of Zhejiang Province (LGF21H160033) and Zhejiang Medical Technology Plan Project (2021KY047) and the Grant for 521 talent project of ZSTU.