
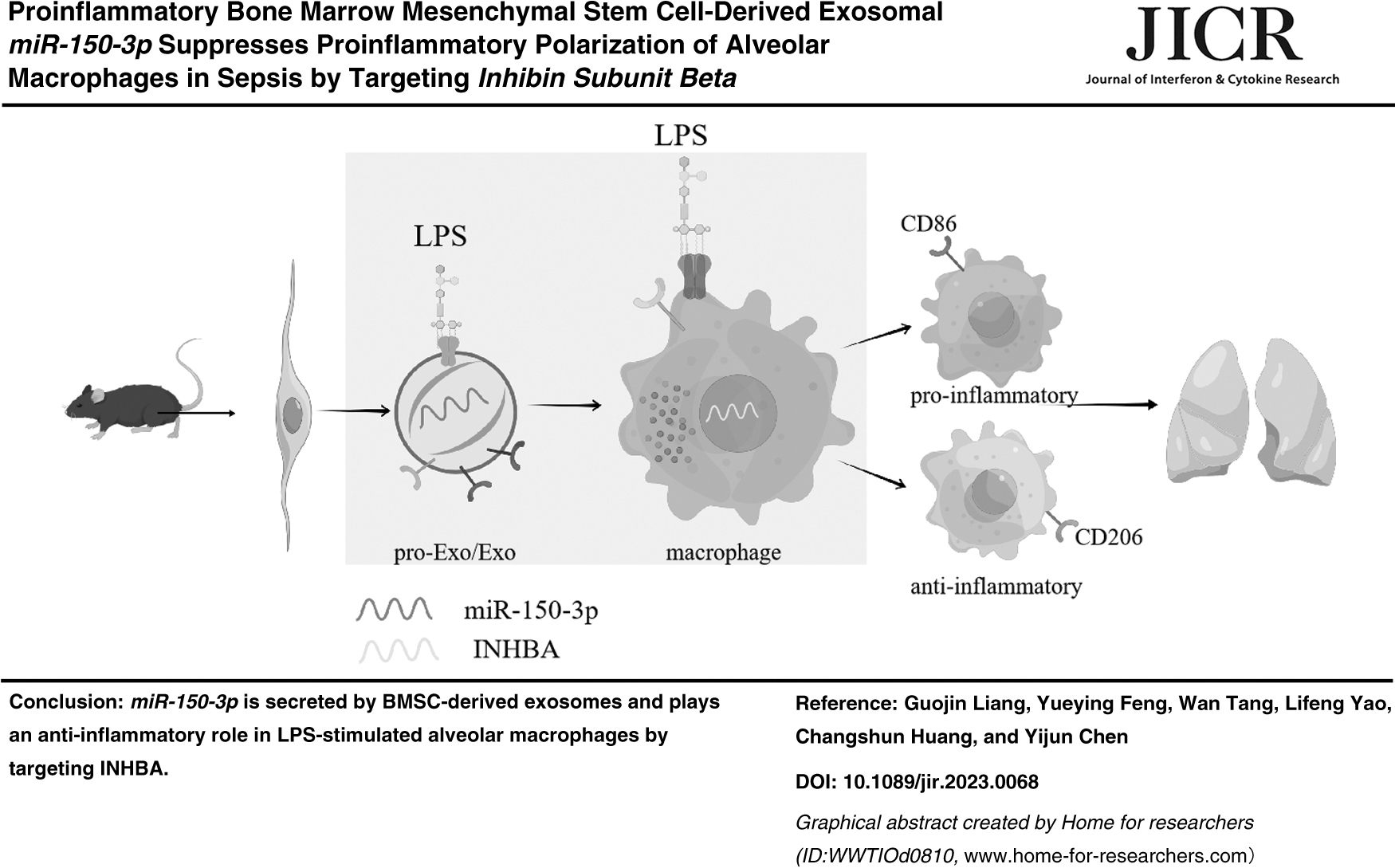
Abstract
Bone marrow mesenchymal stem cell (BMSC)-derived exosomes can protect lung tissues against sepsis, but its related mechanism remains elusive. BMSCs were primed with or without lipopolysaccharide (LPS) before extracting exosomes. The isolated exosomes were identified by transmission electron microscopy, nanoparticle tracking analysis, and western blot. LPS-stimulated macrophages were cocultured with exosomes for 24 h, followed by enzyme-linked immunosorbent assay, flow cytometry, and molecular experiments. Bioinformatics and luciferase assay were employed to investigate the interaction between miR-150-3p and inhibin subunit beta A (INHBA). MiR-150-3p expression was increased in exosomes in a proinflammatory environment. Exosomes suppressed proinflammatory polarization by downregulating IL-6, IL-1β, iNOS, and CD86, as well as promoted anti-inflammatory polarization by upregulating IL-10, ARG-1, and CD206 in LPS-stimulated macrophages. Such effects were more pronounced by LPS-primed exosomes, which was reversed in the absence of miR-150-3p. MiR-150-3p targeted INHBA. INHBA silencing decreased CD86 expression and increased CD206 expression in macrophages, but these effects were reversed by exosomal miR-150-3p inhibition. Proinflammatory BMSC-derived exosomal miR-150-3p suppressed proinflammatory polarization and promoted anti-inflammatory polarization of alveolar macrophages to attenuate LPS-induced sepsis by targeting INHBA.
Introduction
Sepsis is a serious life-threatening systemic inflammatory response syndrome caused by the dysregulation of infection in the host (Singer et al., 2016). Acute lung injury (ALI) and acute respiratory distress syndrome (ARDS) resulted from sepsis are the main causes of high mortality of patients (Kumar, 2020). As the first line of defense of the lung, alveolar macrophages exhibit overactivation when the body is invaded by lipopolysaccharide (LPS) and lead to a cascading release of various cytokines and inflammatory mediators, which is considered to be the root cause of ALI (Arora et al., 2018). Accumulating evidence has shown that microenvironmental stimuli-depended transformation of classically activated (M1 phenotype) and alternatively activated (M2 phenotype) macrophages plays a crucial role in sepsis progression by harnessing inflammatory response (Liu et al., 2018; Wang et al., 2019). Hence, there is growing interest in understanding the regulatory mechanisms of macrophage polarization with the aim of developing novel therapeutic strategies to improve outcomes of patients with sepsis.
Exosomes are defined as biological vesicles (30–150 nm in diameter) produced by cellular exocytosis and carry substances such as proteins, lipids, DNA, and microRNAs (miRNAs) (Doyle and Wang, 2019). As a mediator of cell–cell communication, exosomes are selectively taken up by target cells and influence the biological functions of recipient cells by delivering their packaged cargoes, thus playing an important role in immune response, signaling, and antigen presentation (Kalluri and LeBleu, 2020). Bone marrow mesenchymal stem cells (BMSCs), with their low immunogenicity and immunomodulatory property, become a new research hotspot for effectively regulating inflammatory responses of the body in recent years (Barrachina et al., 2016; Zhang et al., 2020). Although many studies have documented the protective effect of BMSCs against lung injury in sepsis, the underlying molecular mechanisms have not been fully elucidated (Deng et al., 2020; Deng et al., 2022; Lv et al., 2022).
MicroRNAs (miRNAs) are a class of endogenous noncoding RNAs consisting of 15–25 nucleotides and their seed regions are able to bind to the 3′-untranslated region (UTR) of mRNAs, playing a regulatory role in various biological functions of cells (Zhang et al., 2022). According to previous studies, it has been demonstrated that dysregulation of miRNAs is strongly associated with the occurrence and development of multiple inflammatory diseases, including sepsis (Liu et al., 2021a; Renaudineau et al., 2021; Zhang et al., 2021). In recent years, increasing miRNAs have been found to be involved in the progression of sepsis by regulating macrophage polarization, cell apoptosis, or autophagy (Fu et al., 2022; Jiao et al., 2021; Liu et al., 2021b; Zheng et al., 2021). Previously, it has been revealed that miR-150-3p presents a significantly low expression in exosomes of patients with heat stroke (Li et al., 2021). In the study of osteoporosis, animal BMSCs have been found to be enriched with miR-150-3p and their exosomes are able to promote the proliferation and differentiation of osteoblasts by delivering miR-150-3p and thereby alleviate the disease progression (Qiu et al., 2021).
However, the effect of miR-150-3p on regulating macrophage polarization remains unclear.
Therefore, this study was attempted to investigate whether BMSC-derived exosomal miR-150-3p influences the polarization of macrophages in sepsis and further revealed its related molecular mechanism.
Methods
Animal and ethical statement
C57BL/6 mice (male, 10–12 weeks old, 22–25 g) were provided by Beijing HFK Bio-Technology Co., Ltd (Beijing, China) and were reared in a standard animal care room (50% humidity, day–night cycles) with available food and drinking water. All experiments involving animals in this study had been granted by the Committee of Laboratory Animals of Zhejiang Baiyue Biotechnology Co., Ltd (ZJBYLA-IACUC-20220325), and all procedures abided by the guidelines of the China Council on Animal Care and Use.
Isolation of alveolar macrophages and BMSCs
For isolating alveolar macrophages, mice (n = 10) were deeply anesthetized by 2% isoflurane (R510-22-10; Reward Life Science, Shenzhen, China) and sacrificed in the occurrence of cervical dislocation to obtain lungs. Then, lungs from mice were drawn and washed with phosphate-buffered solution (PBS; P1020; Solarbio). After a 10 min centrifugation (1,000 rpm), alveolar macrophages were isolated and then, were cultured in Roswell Park Memorial Institute 1640 medium (RPMI, 11875119; Thermo Fisher, Waltham, MA) at 37°C with 5% CO2 for 4 h (Xiaoyu et al., 2017). The pure alveolar macrophages were obtained after removing the unattached cells by PBS.
The isolation of BMSCs from mice (n = 10) was carried out as previously described (Islam et al., 2012). First, mice were deeply anesthetized by 2% isoflurane (R510-22-10; Reward Life Science, Shenzhen, China) and sacrificed in the occurrence of cervical dislocation to obtain tibias and femurs. Next, the bone marrow cavity was exposed, and DMEM/F12 (11320033; Thermo Fisher) containing 10% fetal bovine serum (FBS; 12103C; Sigma-Aldrich, St. Louis, MO) and penicillin–streptomycin liquid (P1400; Solarbio) was used to flush bone marrow out through a syringe equipped with a 22-gauge needle. After that, the collected bone marrow was centrifuged at 1,200 rpm for 8 min to remove the supernatant, and BMSCs were maintained in a culture flask at 37°C with 5% CO2 for 72 h before nonadherent cell removal and fresh medium replacement. When BMSCs were at passage 3–5 with a passaging rate of 1:3, they were harvested for in vitro experiments.
Cell transfection
To forcedly downregulate miR-150-3p in macrophages or INHBA in BMSCs, miR-150-3p inhibitor (miR20004535-1-5) and inhibitor negative control (miR2N0000002-1-5) were provided by RIBOBIO (Guangzhou, China), as well as short hairpin RNA (shRNA) targeting INHBA (shINHBA; sense: 5′-GCTAGAGTTTACATATTTA-3′, antisense: 5′-TAAATATGTAAACTCTAGC-3′) and shRNA negative control were designed by GenePharma (Shanghai, China). Transfection was performed when the indicated cells were at 90%–95% confluence in a 6-well plate using Lipofectamine 2000 (11668030; Thermo Fisher) or Lipofectamine RNAiMAX (13778075; Thermo Fisher) according to the manufacturer's protocol. After a 48-h incubation at 37°C, the cells were assigned to quantitative real-time reverse transcription–polymerase chain reaction (qRT-PCR) for verification.
LPS treatment
The isolated macrophages or BMSCs were maintained in the culture medium to reach over 80% confluence and washed with PBS twice. Afterward, fresh culture medium containing LPS (100 ng/mL, L861706; Macklin, Shanghai, China) was used to incubate the cells for 24 h as per guidelines (Xu et al., 2019).
Extraction and identification of exosome
Exosomes in conditioned culture medium of BMSCs pretreated with or without LPS were extracted as previously described (Deng et al, 2020). In brief, the culture medium of BMSCs was changed by DMEM/F12 containing 10% exosome-free FBS when the cells grew to around 80% confluence. After a 48-h incubation, the culture supernatant was subjected to gradient centrifugations at 4°C, followed by filtration with the application of a 0.22-μm filter (MPGL04001, Billerica, MA). Then, the obtained medium was given an ultracentrifugation at 100,000 rpm for 70 min to settle down exosomes. Lastly, exosomes were collected and purified after PBS washing and the second ultracentrifugation. As for identification, a transmission electron microscope (EM208S; Olympus, Tokyo, Japan) was utilized to observe the morphology of exosomes; nanoparticle tracking analysis was used to detect the size distribution of exosomes; western blot was employed to assay exosome-related proteins, including tumor susceptibility 101 (TSG101), calnexin, and CD63.
Coculture
Exosomes (10 μg/mL) derived from BMSCs, which had been prestimulated with or without LPS was added into the culture medium of macrophages, followed by a 24-h incubation (Xu et al, 2019). Thereafter, the cells were exposed to 100 ng/mL LPS for another 24 h. As for negative control, macrophages were merely incubated with 100 ng/mL LPS for 24 h.
Enzyme-linked immunosorbent assay
After coculture of macrophages and exosomes, cell culture supernatant was harvested by a 20-min centrifugation at 4°C. Subsequently, enzyme-linked immunosorbent assay (ELISA) was carried out to examine the content of interleukin 6 (IL-6), IL-1β, and IL-10 using commercial mouse ELISA Kits from R&D Systems (SM6000B, SMLB00C; Minneapolis, MN) and FineTest (EM0100-CM1; Wuhan, China) in accordance with corresponding manual instructions.
Western blot
Total protein was extracted by RIPA buffer (R0010; Solarbio), and quantitatively determined by the BCA Protein Assay Kit (23227; Thermo Fisher). Equal protein sample was electrophoresed on 6% or 8% SDS-PAGE, then blotted onto polyvinylidene fluoride membranes (IPVH00010; Solarbio), followed by blocking incubation. Next, the separated proteins were probed with antibodies (Abcam) against TSG101 (ab133586, 1:1,000, 44 kDa), calnexin (ab22595, 1:1,000, 75 kDa), CD63 (ab217345, 1:1,000, 26 kDa), inducible nitric oxide synthase (iNOS; ab178945, 1:1,000, 131 kDa), arginase-1 (ARG-1; ab233548, 1:5,000, 35 kDa), INHBA (PA5-109858, 1:2,000), or GAPDH (ab8245, 1:10,000, 36 kDa) at 4°C overnight, and then, HRP-conjugated Goat Anti-Rabbit IgG (FNSA-0004; FineTest) or Goat Anti-Mouse IgG (FNSA-0003; FineTest) was diluted in TBST solution (T917679; Macklin) and incubated the membranes at room temperature (RT) for 1 h. Eventually, all protein bands were analyzed in an imaging system (5200 Multi; Tanon, Shanghai, China) after being treated with Chemiluminescence Substrate (E917968; Macklin).
Bioinformatics analysis and luciferase assay
Putative mRNAs targeted by miR-150-3p were predicted with the application of TargetScan (
HEK-293 cells (CL-0001; Procell, Wuhan, China) cultured in Minimum Essential Medium (PM150467; Procell) containing 10% FBS was applied to perform luciferase assay. Before the experiment, the 3′-UTR of INHBA, including or excluding binding sites (5′-UGUACCAC-3′) for miR-150-3p was inserted into psiCHECK-2 (C8021; Promega, Madison, WI) to construct INHBA-WT or INHBA-MUT reporter plasmids, which were cotransfected with miR-150-3p mimic (miR10004535-1-5; RIBOBIO) or mimic control (miR1N0000002-1-5; RIBOBIO) into HEK-293 cells using Lipofectamine 2000 for 48 h. The following luciferase assay was carried out with the application of Dual Luciferase Reporter Gene Assay Kit (RG027; Beyotime, Shanghai, China) according to the manufacturer's protocol. The result of luciferase activity in the cells was detected using a luminometer (GloMax-96; Promega).
RNA isolation and qRT-PCR
Total RNA or miRNA from macrophages or BMSCs was isolated by TRIzol (15596026; Thermo Fisher) and then, was reversely transcribed into complementary DNA (cDNA) using miRNA First-Strand cDNA Synthesis (B532453; Sangon Biotech, Shanghai, China) or the PrimeScript RT Kit (RR037B; TaKaRa, Tokyo, Japan). Afterward, the amplification of cDNA was developed in a thermal cycler (ABI7500; Applied Biosystems, Carlsbad, CA) using TB Green Mixture (RR430A; TaKaRa).
All primer sequences were as follows (5′-3′): miR-150-3p (CGCTGGTACAGGCCTGGG, AGTGCAGGGTCCGAGGTATT), U6 (CAGCGCAGAATCACCCCAT, CGCACCATGTCAATGATCTCA), IL-6 (TAGTCCTTCCTACCCCAATTTCC, TTGGTCCTTAGCCACTCCTTC), IL-1β (GCAACTGTTCCTGAACTCAACT, ATCTTTTGG GGTCCGTCAACT), iNOS (TGCCAGGGTCACAACTTTACA, CTCTCCACTGCCCCAGTTTT), IL-10 (GCTCCAAGACCAAGGTGTCT, AGGACACCATAGCAAAGGGC), ARG-1 (CTCCAAGCCAAAGTCCTTAGAG, GGAGCTGTCATTAGGGACATCA), INHBA (GTGGAGTTGGAGCTTTGTGAAG, TGTTTTGTCAGCCGGCTCTT), and GAPDH (AGGTCGGTGTGAACGGATTTG, TGTAGACCATGTAGTTGAGGTCA). The fold changes representing relative miRNA or mRNA expression was calculated by the 2−ΔΔCt method (Livak and Schmittgen, 2001), and U6 or GAPDH was functioned as the normalizer.
Flow cytometry
After coculture, the percentage of macrophages polarized toward proinflammatory phenotype or anti-inflammatory phenotype was assayed using flow cytometry. Briefly, the cocultured macrophages were harvested in centrifuge tube and incubated with FITC-conjugate CD86 antibody (11-0862-82; Thermo Fisher) or CD206 antibody (MA5-16870; Thermo Fisher) at RT in the dark. After 20 min, the cells were rinsed with PBS and analyzed in a flow cytometer (FACS Caliber; BD Pharmingen, San Diego, CA).
Statistical analyses
Measurement data were shown as mean ± standard deviation. Statistical comparison was analyzed either by independent-samples t-test, or by one-way analysis of variance. All statistical analyses were carried out using GraphPad Prism 8.0 (GraphPad Software, Inc., San Diego, CA). Differences were taken to be statistically significant if P-values were <0.05.
Results
Exosomes were successfully isolated from BMSCs and LPS stimulation increased the expression of miR-150-3p in BMSC-derived exosomes
As captured in Fig. 1A, BMSC-derived exosomes were structurally manifested as cup-shaped vesicles and double lipid membranes, which is consistent with the classical morphology of exosomes (Toh et al., 2018). The result of nanoparticle tracking analysis demonstrated that the diameter of exosomes was mainly in the range of 100–200 nm (Fig. 1B). Additionally, the results of western blot confirmed that BMSC-derived exosomes showed positive for exosomal markers (TSG101 and CD63) and negative for endoplasmic reticulum protein, calnexin (Fig. 1C). Previous evidence shows that BMSC-derived exosomes under a proinflammatory environment can attenuate LPS-triggered ARDS (Deng et al, 2020) To mimic the proinflammatory environment in the body, BMSCs were maintained in LPS for 24 h. Compared with normal BMSC-derived exosomes, it was observed that miR-150-3p expression was increased in BMSC-derived exosomes after LPS treatment, as assayed by qRT-PCR (Fig. 1D, P < 0.001).

The identification of BMSC-derived exosomes and miR-150-3p expression.
BMSC-derived exosomes suppressed inflammation in LPS-stimulated macrophages
Next, we stimulated macrophages with LPS for 24 h, followed by coculture with BMSC-derived exosomes. Compared with normally cultured macrophages, qRT-PCR determined that LPS increased IL-6, IL-1β, iNOS, IL-10, and ARG-1 expressions in macrophages (Fig. 2A, P < 0.001). After a 24-h coculture, BMSC-derived exosomes reduced IL-6, IL-1β, and iNOS expressions and enhanced IL-10 and ARG-1 expressions in LPS-stimulated macrophages (Fig. 2A, P < 0.001), and these tendencies were more pronounced when the exosomes were primed with LPS (Fig. 2A, P < 0.01). As revealed in Fig. 2B–D, LPS enhanced the release of IL-6, IL-1β, and IL-10 from macrophages (P < 0.01). Compared with macrophages undergone LPS stimulation alone, the presence of BMSC-derived exosomes suppressed IL-6 and IL-1β releases and promoted IL-10 release (Fig. 2B–D, P < 0.001), and these tendencies were more pronounced in the presence of proinflammatory BMSC-derived exosomes (Fig. 2B–D, P < 0.001).
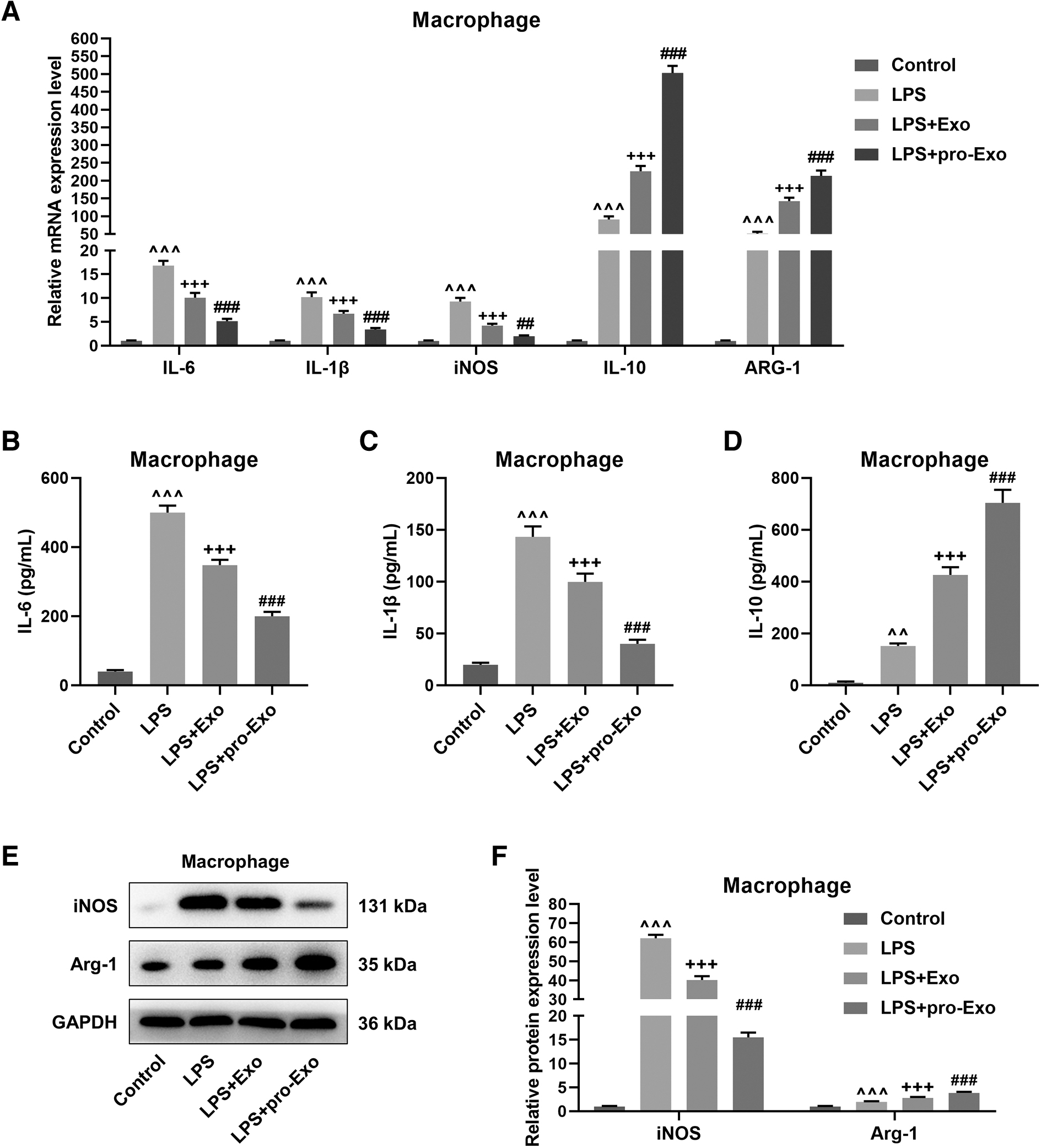
The effect of proinflammatory BMSC-derived exosomes on the polarization of LPS-treated macrophages.
BMSC-derived exosomes promoted anti-inflammatory–phenotype polarization of LPS-stimulated macrophages
As revealed in Fig. 2E, F, LPS stimulation upregulated iNOS and Arg-1 proteins in macrophages (P < 0.001). Coculture with normal BMSC-derived exosomes reduced the protein expression of iNOS but elevated the protein expression of Arg-1 in LPS-stimulated macrophages (P < 0.001), which were reinforced when the exosomes were pretreated with LPS (P < 0.001). In addition, the results of flow cytometry demonstrated that LPS significantly increased the percentage of CD86 and slightly increased the percentage of CD206 in macrophages (Fig. 3, P < 0.001). In contrast with macrophages undergone LPS stimulation alone, it was discerned that the percentage of CD86 was decreased and the percentage of CD206 was increased in LPS-stimulated macrophages after coculture with normal exosomes (Fig. 3, P < 0.001), and these situations more noticeable when the exosomes were pretreated with LPS (Fig. 3, P < 0.001).

The effect of proinflammatory BMSC-derived exosomes on CD86 and CD206 expressions in LPS-treated macrophages. BMSCs were pretreated with or without LPS for 24 h, followed by exosome extraction. Macrophages were cocultured with or without exosomes for 24 h, and stimulated with LPS for 24 h. Flow cytometry was performed to analyze the level of CD86 and CD206 in macrophages. ^^^ P < 0.001, versus Control; + P < 0.05, +++ P < 0.001, versus LPS; ### P < 0.001, versus LPS + Exo.
MiR-150-3p depletion reversed the effect of proinflammatory BMSC-derived exosomes on LPS-stimulated macrophages
After cell transfection, miR-150-3p inhibitor considerably downregulated the expression of miR-150-3p in BMSCs (Fig. 4A, P < 0.001) and yielded from LPS-primed BMSCs (Fig. 4B, P < 0.001). To confirm the potential effect of miR-150-3p on the polarization of macrophages in sepsis, we conducted rescue experiments the depletion of miR-150-3p upregulated IL-6 and IL-1β as well as downregulated IL-10 in both macrophages and cell culture supernatant under the LPS-stimulated coculture environment, which reversed the regulatory effect of proinflammatory BMSC-derived exosomes (Fig. 4C–F, P < 0.05). Meanwhile, the decreased iNOS protein level and the increased ARG-1 protein level induced by proinflammatory BMSC-derived exosomes in LPS-stimulated macrophages were evidently restored after the loss of miR-150-3p (Fig. 4G, H, P < 0.001). Furthermore, the inhibiting effect of proinflammatory BMSC-derived exosomes on CD86 expression and its promoting effect on CD206 expression in LPS-stimulated macrophages were also negated by miR-150-3p depletion (Fig. 5, P < 0.001).
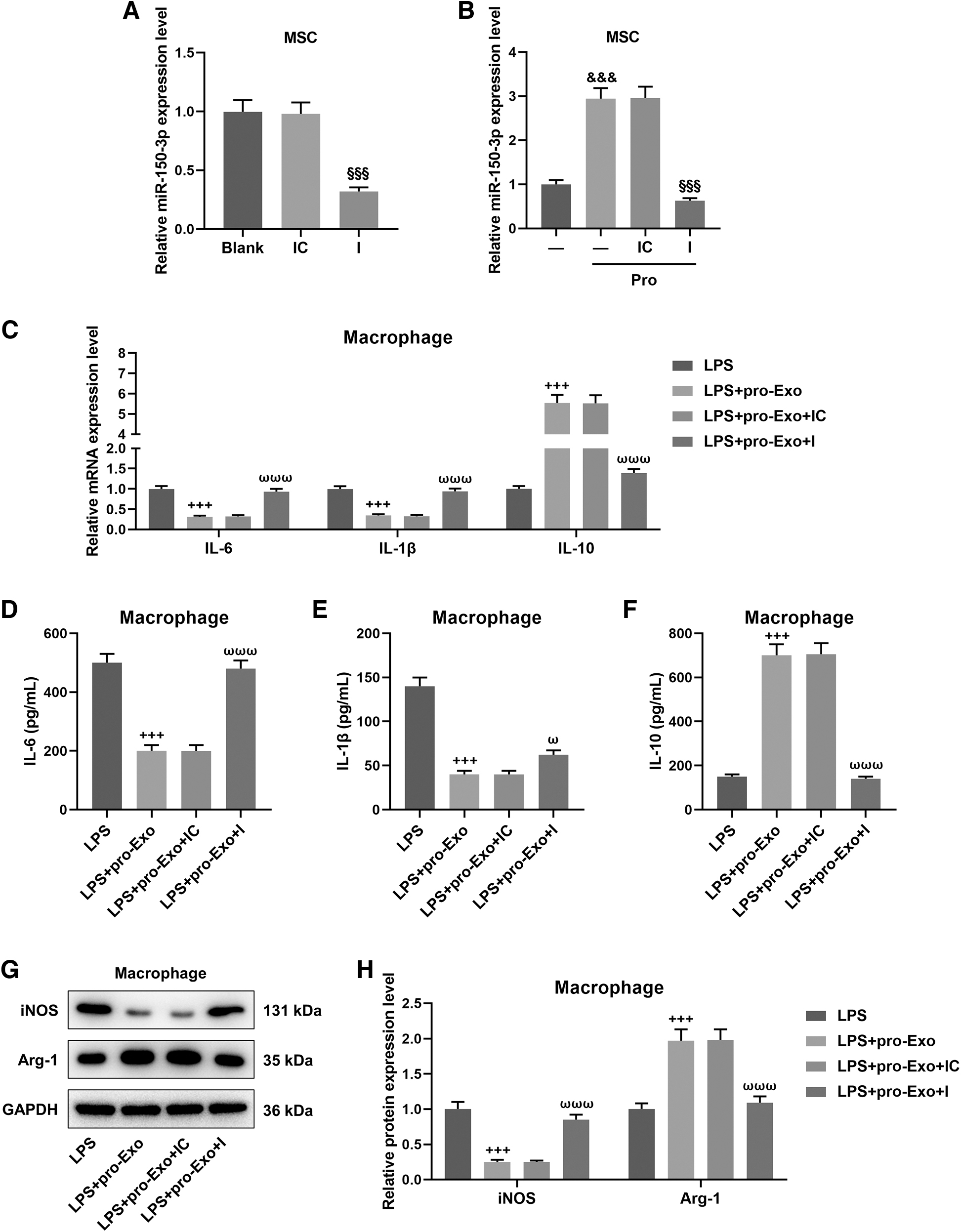
The involvement of proinflammatory BMSC-derived exosomal miR-150-3p in the polarization of LPS-treated macrophages.
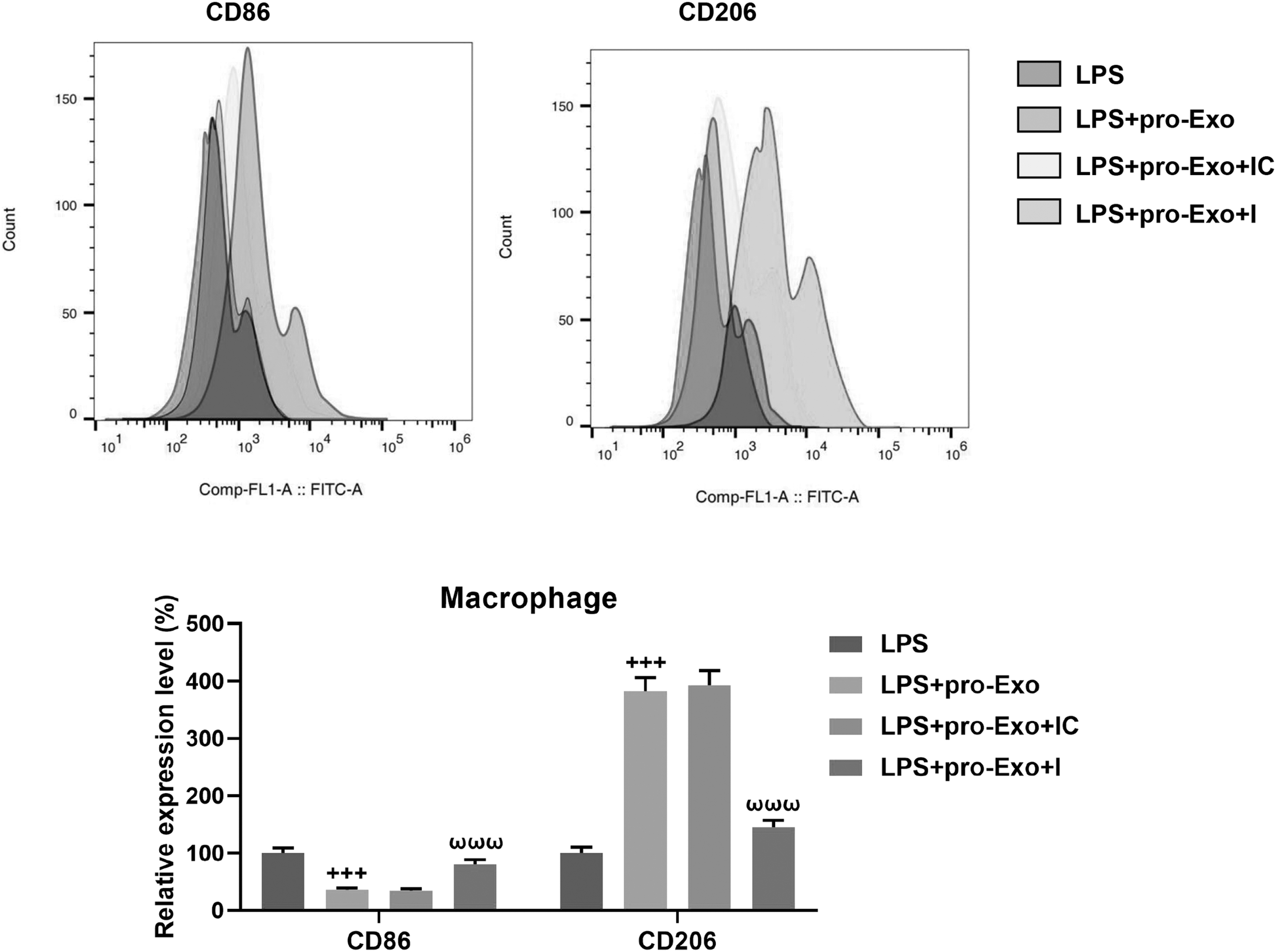
The involvement of proinflammatory BMSC-derived exosomal miR-150-3p in CD86 and CD206 expressions in LPS-treated macrophages. BMSCs were subjected to miR-150-3p inhibition through cell transfection and then, were pretreated with LPS for 24 h before exosome extraction. Macrophages were cocultured with or without exosomes for 24 h, and stimulated with LPS for 24 h. Flow cytometry was performed to analyze the level of CD86 and CD206 in macrophages. +++ P < 0.001, versus LPS; ωωω P < 0.001, versus LPS + Exo + IC.
Exosomal miR-150-3p regulated the polarization of macrophages by targeting INHBA
On the basis of bioinformatics analysis, 3 mRNAs (AK3, CTSC, INHBA) were obtained as downstream targets of miR-150-3p during the polarization of macrophages (Fig. 6A). Since a previous finding that INHBA was highly expressed in proinflammatory macrophages (Su et al., 2016), we selected INHBA as a possible target gene in the following investigations. As shown in Fig. 6B, TargetScan analyzed putative binding sites of INHBA 3′-UTR for miR-150-3p. According to the results of luciferase assay, it was determined that miR-150-3p mimic evidently suppressed the luciferase activity of INHBA-WT in HEK-293 cells (Fig. 6C, P < 0.01), confirming that INHBA is a bona fide target of miR-150-3p. By cell transfection, the expression of INHBA in macrophages was markedly downregulated by shINHBA (Fig. 6D, P < 0.001).
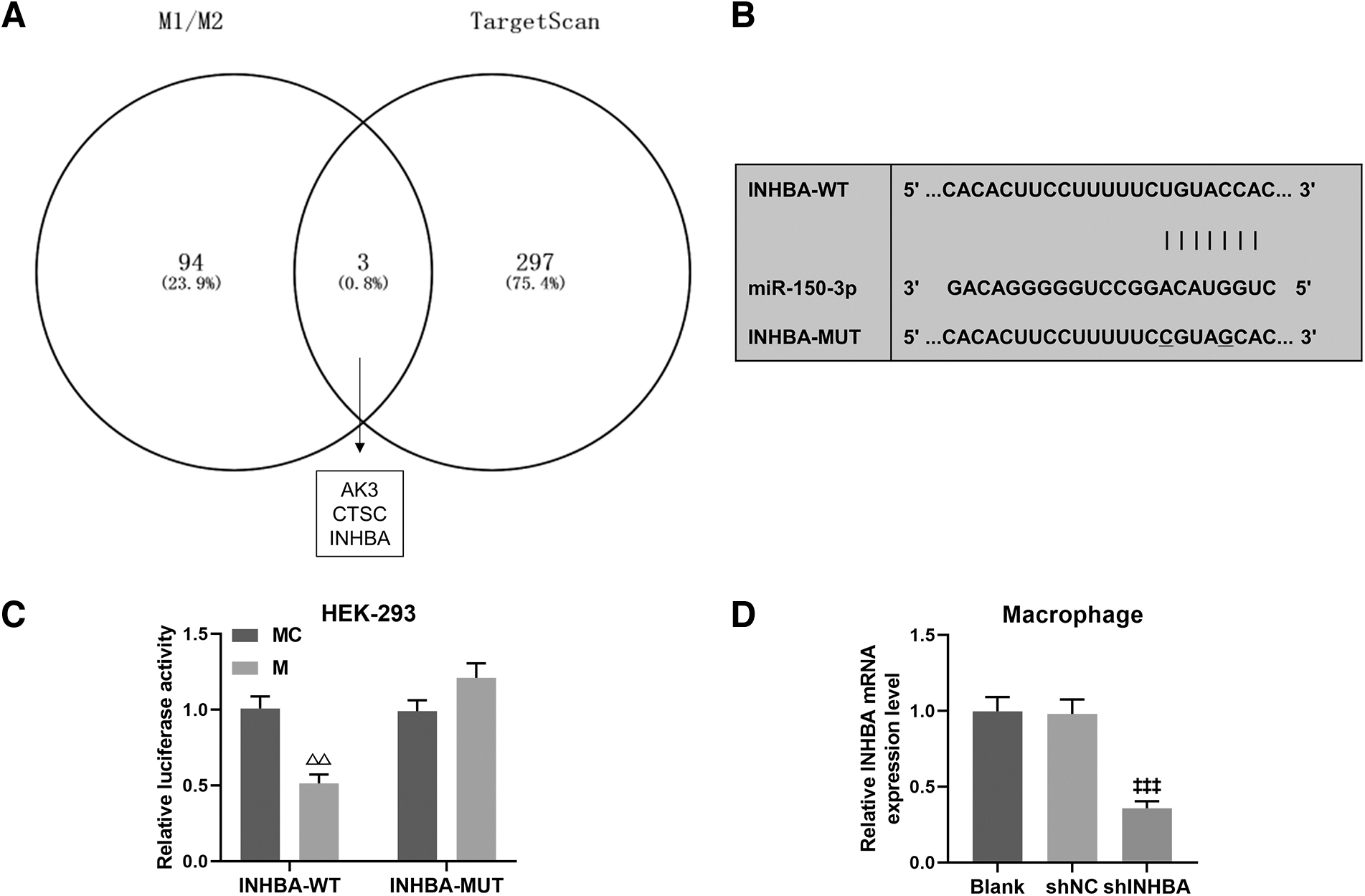
The prediction and verification of miR-150-3p targeting INHBA.
After coculture of macrophages and exosomes, INHBA silencing inhibited anti-inflammatory–phenotype polarization of macrophages by downregulating CD86 and facilitated anti-inflammatory–phenotype polarization of macrophages by upregulating CD206 (Fig. 7, P < 0.05), and these effects were reversed when miR-150-3p was exhausted in exosomes (Fig. 7, P < 0.05). On the contrary, miR-150-3p depletion exosomes increased the percentage of proinflammatory macrophages by upregulating CD86 and reduced the percentage of anti-inflammatory macrophages by downregulating CD206 (Fig. 7, P < 0.001), which were counteracted by INHBA silencing (Fig. 7, P < 0.001).

The regulatory effect of miR-150-3p/INHBA axis on macrophage polarization. BMSCs were transfected with miR-150-3p inhibitor before exosome extraction, and macrophages was transfected with shINHBA. After a 24-h coculture of macrophages and exosomes, flow cytometry was performed to analyze the level of CD86 and CD206 in macrophages. † P < 0.05, ††† P < 0.001, versus IC + shNC; Ω p < 0.05, ΩΩΩ p < 0.001, versus IC + shINHBA; @@@ p < 0.001, versus I + shNC.
Discussion
At present, the main treatments for ALI in sepsis include anti-inflammatory therapy, hormonal therapy, and ventilator support. Although these treatments exert a positive therapeutic effect on patients, most antimicrobial drugs are not effective in improving patient survival with the increasing number of multidrug-resistant bacterial strains (Chang and Holcomb, 2016). M1 macrophages and M2 macrophages are 2 classical subtypes of macrophages, and owing to the plasticity of macrophages, these 2 phenotypes can be transformed into each other depending on microenvironmental stimuli. It is wildly accepted that the disruption of dynamic balance of M1 and M2 macrophages is responsible for the development of many inflammatory diseases, including sepsis (Funes et al., 2018). M1 macrophages account for the dominant proportion in the early stages of sepsis and play a promoting role in cellular immunity by continuously releasing proinflammatory cytokines, such as IL-6, TNF-α, and IL-1β, to kill pathogenic microorganisms, resulting in an uncontrolled inflammation to exacerbate tissue damage (Jiao et al, 2021).
Available studies have documented that restoring the homeostasis of M1 and M2 macrophages by inhibiting M1 polarization and promoting anti-inflammatory M2 polarization can mitigate disease progression in sepsis mouse models (Dang and Leelahavanichkul, 2020; Feng et al., 2019).
Previously, BMSCs have been demonstrated to play a protective role in attenuating lung injury in response to sepsis through regenerative repair, immunomodulation, antioxidation, and antiapoptosis (Gill and Gorlick, 2021; Guo et al., 2021; Swaminathan et al., 2018). Additionally, evidence has shown that tail vein injection of human umbilical cord-derived MSCs could induce polarization of macrophages toward M2 phenotype, which is attributable to exosome-mediated cell–cell communication (Song et al., 2017). It is increasingly believed that BMSC-derived exosomes share similar anti-inflammatory and immune regulation functions as BMSCs in that they can deliver key information (proteins, mRNA, miRNA, etc.) from BMSCs to targeted cells during the intercellular communication, and thereby participate in many cellular physiopathological processes (Li et al., 2015). Currently, it has been confirmed that BMSC-derived exosomes can reduce the inflammatory response in the lung and promote the recovery of damaged lung tissues, indicating that they have great promise in the treatment of ARDS (Deng et al, 2020).
However, how BMSC-derived exosomes affect alveolar macrophage polarization in a septic environment needs to be elucidated. In this study, we observed that the increased level of proinflammatory cytokines (IL-6, IL-1β) was inhibited, and the increased level of IL-10 was further promoted in LPS-treated macrophages after the coculture of BMSC-derived exosomes. As a famous anti-inflammatory cytokine, IL-10 is mainly secreted by macrophages, T cells, or NK cells (Martinez-Espinosa et al., 2021). The overexpression of IL-10 in macrophages has been previously reported to attenuate LPS-driven inflammation in endothelial cells (Yi et al., 2022). Collectively, it is indicated that BMSC-derived exosomes could reduce inflammatory transformation of macrophages upon LPS stimulation. According to qRT-PCR and western blot, we also discerned that BMSC-derived exosomes downregulated iNOS and upregulated ARG-1 in LPS-treated macrophages. Moreover, a reduction of CD86 level and an elevation of CD206 were induced by BMSC-derived exosomes in LPS-treated macrophages.
It is well known that the increase of CD86 and iNOS expressions marks the polarization of macrophages toward proinflammatory M1 phenotype, and when macrophages exhibit an anti-inflammatory property, they specifically express CD206 and ARG-1 (Liu et al., 2019). Based on abovementioned findings, it is revealed that the anti-inflammatory effect of BMSC-derived exosomes on LPS-driven lung injury is accomplished by inducing macrophage polarization from M1 phenotype to M2 phenotype. Intriguingly, proinflammatory stimuli like LPS or TNF-α act as a warning to MSCs, thereby causing them to exhibit a greater effect on combating inflammation (Liu et al., 2016; Selvasandran et al., 2018; Wang et al., 2009). We subsequently pretreated BMSCs with LPS to mimic a proinflammatory environment in vitro, revealing that LPS-primed exosomes exerted a stronger effect on promoting M2 polarization and suppressing M1 polarization of LPS-stimulated macrophages, which are in accordance with the findings of Xu et al. (2019). However, the molecular mechanism functioning in this process is lack of evidence.
As a carrier shuttling between cells, exosomes have been found to harbor a variety of miRNAs to play different biological functions during the development of inflammation-related diseases (Jiao et al, 2021; Park et al., 2022). Previously, abundant miR-150-3p as cargoes in BMSC-derived exosomes were identified by Qiu et al. (2021), and this miRNA is considered as a promising target for treating osteoporosis. In patients with heat stroke, the deficiency of plasma-derived exosomal miR-150-3p was reported to participate in intercellular communications in the occurrence of cascade inflammation (Li et al, 2021). Although miR-150 has been reported to be one of abnormal miRNA expressions with serum obtained from patients with sepsis and its low expression is associated with disease severity (Zhou et al., 2015), there are few reports regarding the regulatory effect of miR-150-3p on sepsis progression. In this study, we determined that the expression of miR-150-3p in BMSC-derived exosomes was considerably upregulated in a culture environment containing LPS.
After forcedly downregulating miR-150-3p in BMSCs, the effect of exosomes on regulating M1 and M2 polarization of LPS-stimulated macrophages was impaired, suggesting that BMSC-derived exosomal miR-150-3p is implicated in macrophage polarization through intercellular crosstalk. Mechanistically, we further screened out downstream mRNAs (AK3, CTSC, INHBA) of miR-150-3p through bioinformatics analysis. Among them, both CTSC and INHBA have been reported to be positively involved in the polarization of M1 macrophage (Dai et al., 2021; Liang et al., 2022), but the upregulation of INHBA was confirmed to contribute to the polarization of M2 macrophage in systemic juvenile idiopathic arthritis (Li et al., 2016). In this study, miR-150-3p was validated to target INHBA, as shown by luciferase assay. The aberrant expression of INHBA have been broadly found to be associated with metastasis and infiltration of various tumors, but its biological function in inflammatory diseases remains unclear (Yu et al., 2021; Zhao et al., 2021).
In this study, we found that INHBA silencing in macrophages promoted M2 macrophages by downregulating CD86 and upregulating CD206, which was reversed by miR-150-3p depletion in BMSC-derived exosomes. On the other hand, the regulatory effect of miR-150-3p depletion on macrophage polarization was negated by INHBA silencing.
Conclusion
In conclusion, this study demonstrates that BMSC-derived exosomes can ameliorate sepsis-caused inflammation in ALI by suppressing proinflammatory macrophages and promoting anti-inflammatory macrophages, which is mediated by intercellular communication. Furthermore, our present findings substantiate, for the first time that miR-150-3p is secreted by BMSC-derived exosomes and plays an anti-inflammatory role in LPS-stimulated alveolar macrophages by targeting INHBA. Therefore, this study suggested that BMSC-derived exosomal miR-150-3p as a potential therapeutic strategy provide new insights for future treatment of septic ALI.
Footnotes
Authors' Contributions
G.L.: Substantial contributions to conception and design; Y.F., W.T., L.Y., C.H., and Y.C.: Data acquisition, data analysis, and interpretation; G.L. and Y.F.: Drafting the article or critically revising it for important intellectual content; All authors: Final approval of the version to be published; and G.L., Y.F., W.T., L.Y., C.H., and Y.C.: Agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of the work are appropriately investigated and resolved.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by The Ningbo Science and Technology Program Project [202003N4272].