
Abstract
Although the modulation of immunity by exercise has been a long-studied paradigm, the molecular pathways connecting the two are still not fully understood. Regular moderate aerobic exercise is associated with improved health and directly impacts the immune system by changing the proportion of cell subpopulations, their function, and interleukin production. The endocannabinoid system has gained importance as an immune modulator, affected by moderate aerobic promoting the production of endocannabinoids, which are ligands of the cannabinoid receptors (CBRs) expressed on the surface of all immune cells. Our group previously reported a reduction of lymphocytic populations in the spleen of chronically exercised rats, accompanied by an increase in CBR expression. Given the complex and compartmentalized nature of the immune system, we decided to study the effects of chronic exercise on the proportion of peripheral blood mononuclear cells, serum interleukins, and the expression of CBRs on these cells. Overall, our results indicate that chronic exercise decreases the proportion of T helper and Tγδ cells but increases the expression of cannabinoids (CBR1) on T helper and natural killer cells, and the production of interleukins, including IL-1β, interferon-gamma, tumor necrosis factor-alpha, IL-10, and IL-4, suggesting higher reactivity and efficiency from the immune system conferred by exercise.
Introduction
Since living a sedentary (SED
Nevertheless, the impact of exercise on the physiology of mammals is not restricted to the immune system; it can also alter several other aspects ranging from adipose tissue content to neurotransmitters (Fang et al., 2013; Gleeson et al., 2011; Hoffman-Goetz and Pedersen, 1994; Lancaster and Febbraio, 2014), and many of these physiological changes have a direct or indirect repercussion for the immune system elements. These pathways with immunoregulatory potential involve changes in energy substrates (Palmer et al., 2015), activation of signaling pathways directly relevant to immune regulation, such as the release of interleukin (IL)-6 by skeletal muscle (Lancaster et al., 2004), and the release of stress hormones, catecholamines (Padgett and Glaser, 2003), and neurotransmitters by the sympathetic and parasympathetic systems. The cannabinoid system, which remains understudied, is also implicated in these pathways.
The endocannabinoid system is composed of a variety of receptors (CBR), the most classically studied being CBR1 and CBR2, and its endogenous ligands (Abood, 2005; Howlett, 2005). As it has been demonstrated, anandamide and other endocannabinoids increase in circulation during and after an aerobic exercise bout (Heyman et al., 2012; Sparling et al., 2003). This increase in endocannabinoids could potentially generate an immune modulatory function through their interaction with the CBRs found on the immune cells (Galiègue et al., 1995). Furthermore, some studies have suggested an immune modulatory function of the endocannabinoid system, since it is capable of inhibiting proinflammatory responses, proliferation in cells from the immune system, and the amount of CBR's is highly increased in activated cells (Adhikary et al., 2012; Derocq et al., 1995; Klein et al., 2003).
A previous study from our group demonstrated that our chronic moderate aerobic exercise model reduced the proportion of CD3+CD4+ and CD45 RA+ cells in the spleen, enhanced the expression of CBR's in the same subpopulations, and enhanced the proliferation of natural killer (NK) cells (Valencia-Sanchez et al., 2019). Since the immune system is a complex and compartmentalized machinery, the objective of this study is to expand our knowledge on this popular and, in terms of public health, important exercise paradigm. We aim to test if these effects extend to peripheral elements of the immune system through the analysis of the subpopulation cells of peripheral blood mononuclear cells (PBMCs), the expression of CBR1 on their surface, and the assessment of the modulation in the production of several pro- and anti-inflammatory interleukins.
Our findings not only expand our knowledge of the physiological consequences of this particular type of exercise but also support the idea of an improved and more efficient immune system conferred by it.
Materials and Methods
Ethic statement
Animal experimental practices were conducted at the animal facilities of the Instituto de Investigaciones Biomédicas (IIB), Universidad Nacional Autónoma de México (UNAM). All experimental procedures described in this article were approved by the Institutional Care and Animal Use Committee (CICUAL), which strictly adheres to Mexican regulation (NOM-062-ZOO-1999), and the National Institute of Health (NIH) Guide for the Care and Use of Laboratory Animals. The euthanasia of experimental animals was performed humanely. The study was reported in accordance with ARRIVE guidelines.
Animals
Male Wistar rats weighting 250–300 g were obtained from our own breeding colony (IFC, UNAM). The animals were group-housed under control temperature (22°C ± 1°C) and 12 h light-dark cycle, and water and chow were provided ad libitum (Purina LabDiet 5015; Purina, St. Louis, MO). After being anesthetized with Pisabental® (PISA Farmacéutica, Guadalajara Jalisco, México), the animals were euthanized by cervical dislocation.
Exercise protocol
Three experimental groups were determined for this work: Exercised (EXE), Treadmill control (TC), and SED group. The animals in the EXE group ran on a treadmill 5 days a week for 10 weeks. After the habituation week to the treadmill, the animals were trained to run at 7.5 m/min for 10 min. The speed and exercise duration were progressively increased until the third week when they reached a speed of 15 m/min for 40 min, as previously reported (Valencia-Sanchez et al., 2019). This speed was maintained until the end of the 10-week period. It should be noted that this protocol corresponds to moderate and chronic training in rats (Carvalho et al., 2005) and that no stressful stimulus was used to encourage the animals to run on the treadmill.
Animals in TC were placed inside the treadmill and then turned on at minimum capacity (4 m/min) for 10 min, five times per week, during the 10-week period. Subsequently, the animals in this experimental group were exposed to the same conditions as those in the EXE group, including noise, vibrations, enclosure, and manipulation. Animals in SED group were kept under standard conditions for the same period as those in the other experimental groups. After the 10-week period, animals were allowed to rest for 24 h before being euthanized by an overdose of pentobarbital sodium (Pisabental, México), to eliminate any acute effects resulting from exercise or exposure to the treadmill.
Flow cytometry
Blood was collected in EDTA-coated collection tubes (∼4 mL) and kept on ice until further processing. Lysis of erythrocytes was carried out using ACK buffer (150 mM NH4Cl, 10 mM KHCO3, 0.1 mM Na2EDTA, pH 7.3) for 10 min and washed three times with phosphate-buffered saline (PBS) afterward. After the last PBS wash, cells were resuspended in FACS buffer (PBS, Fetal Bovine Serum, 0.02% NaN3). Reagents were purchased from
Approximately 106 cells were incubated with the following antibodies to characterize the blood immune cell subpopulations. Antibodies used were Alexa Fluor® 488-conjugated anti-rat CD3 (clone IF4; BioLegend, San Diego, CA), PE Cy5-conjugated anti-rat CD4 (clone OX-35; BioLegend), PE-conjugated anti-rat CD8a (clone OX-8; BioLegend), PE-conjugated anti-rat CD45 RA (clone OX-33; BioLegend), Alexa Fluor® 647-conjugated anti-rat CD161 (BioLegend), biotin-conjugated anti-rat CD11b/c (OX-42; BioLegend), and PE-conjugated anti-rat TCRγδ (clone V65; BioLegend). PeCy5-Streptavidin was used for the identification of CD11b/c biotinylated antibody. CD11b/c was used to determine the percentage of monocyte/neutrophil subpopulations.
To determine the expression of CBR1 on immune cells, the polyclonal rabbit anti-cannabinoid receptor I (Abcam®), and the Secondary antibodies Alexa Fluor® 488-conjugated goat anti-rabbit immunoglobulin G (IgG) (Thermo Fisher Scientific, Rockford, IL) or DyLight® 649-conjugated anti-rabbit IgG (Vector Laboratories, Newark, CA) were included in the panel of staining for the immune cell markers.
Cytokine measurements
Eight different cytokines IL-1β, IL-4, IL-5, IL-6, IL-10, IL-13, interferon-gamma (IFN-γ), and tumor necrosis factor-alpha (TNF-α) were measured using the MILLIPLEX®MAP Rat Cytokine/Chemokine Magnetic Bead Panel (RECYTMAG-65K) (Merck KGaA, Darmstadt, Germany). TGF-β1 was determined using the MILLIPLEX®MAP TGF-β1 Single Plex Magnetic Bead Kit (TGFBMAG-64K-01) (Merck KGaA). Frozen serum samples were thawed and prepared according to manufacturer instructions. In brief, a mixture of magnetic beads specific from the eight cytokines described above was incubated with serum from the three experimental groups overnight at 4°C, then we used six serial dilutions to build the standards curve and used two quality controls provided by the manufacturer for each cytokine, first with a low range and second with a high range of concentration. After incubation, plates were washed and incubated with a mixture of correspondent biotinylated antibody, later, streptavidin-phycoerythrin was added.
Median fluorescence intensity was obtained from duplicates of serum samples from Wistar male rats that were assigned to one of three experimental groups: SED group, TC, or EXE. Samples from three independent experiments were carried out within each experimental group (n = 7) using a MAGPIX from LUMINEX by Millipore, which was facilitated at the Unidad de Investigación from Facultad de Veterinaria, UNAM.
The obtained data were further analyzed using MyAssays software package with the Four Parameter Logistic Curve fitting method to calculate the sample cytokine concentration. GraphPad PRISM software was used for graphic and statistical analysis. Results are expressed as mean ± standard error of the mean, n = 7. Significant changes were determined with one-way analysis of variance (ANOVA) (α = 0.05) followed by Tukey's post hoc test. Differences were considered statistically significant after P < 0.05.
Corticosterone analysis
Euthanasia of the experimental animals was carried out at the time set for the daily exercise period (14:00–16:00 h). Blood was obtained through cardiac puncture and immediately centrifuged at 4000 rpm to collect serum. The serum was then divided and stored at −70°C for later use in corticosterone and cytokine analysis. Corticosterone levels were assessed using a Corticosterone ELISA kit (Abcam® ab-108821) following the manufacturer's protocol. After the reaction was stopped, the plate was read using a Stat Fax 4200 microplate reader (Awarness Technology).
Statistical analysis
Immune cell subpopulation, the expression of CBR1, and the cytokine analysis were determined using one-way ANOVA, followed by a Tukey's post hoc test. A P-value of <0.5 was considered significant. Both the p-value and sample size (n) are reported in each section of the results. Normal distribution of the data was assessed using Shapiro-Wilk test.
Results
Chronic moderate exercise affects total weight but not water or food consumption in rats
To assess the impact of moderate exercise in animals, we made use of three experimental groups: EXE, TC, and SED group. First, we monitored the food and water consumption for each group on a weekly basis and recorded their body weight at the end of the 10-week period. Food and water consumption did not vary significantly across experimental groups (P = 0.3309, n = 6, and P = 0.3312, n = 6, respectively). However, a significant reduction in body weight was observed in the EXE group (452 g) compared to the two control groups, TC (490.8 g) and SED (495.8 g).
Immune cell proportion detection by flow cytometry
We analyzed the proportions of innate and adaptive immune cells in the blood of animals from the three experimental groups. We removed some samples due to staining variability to consider the reliability of our results. Therefore, sample sizes considered for this section are SED (n = 9), TC (n = 8), and EXE (n = 11). Figure 1 shows the staining and gating strategy used for the identification of these immune subpopulations.
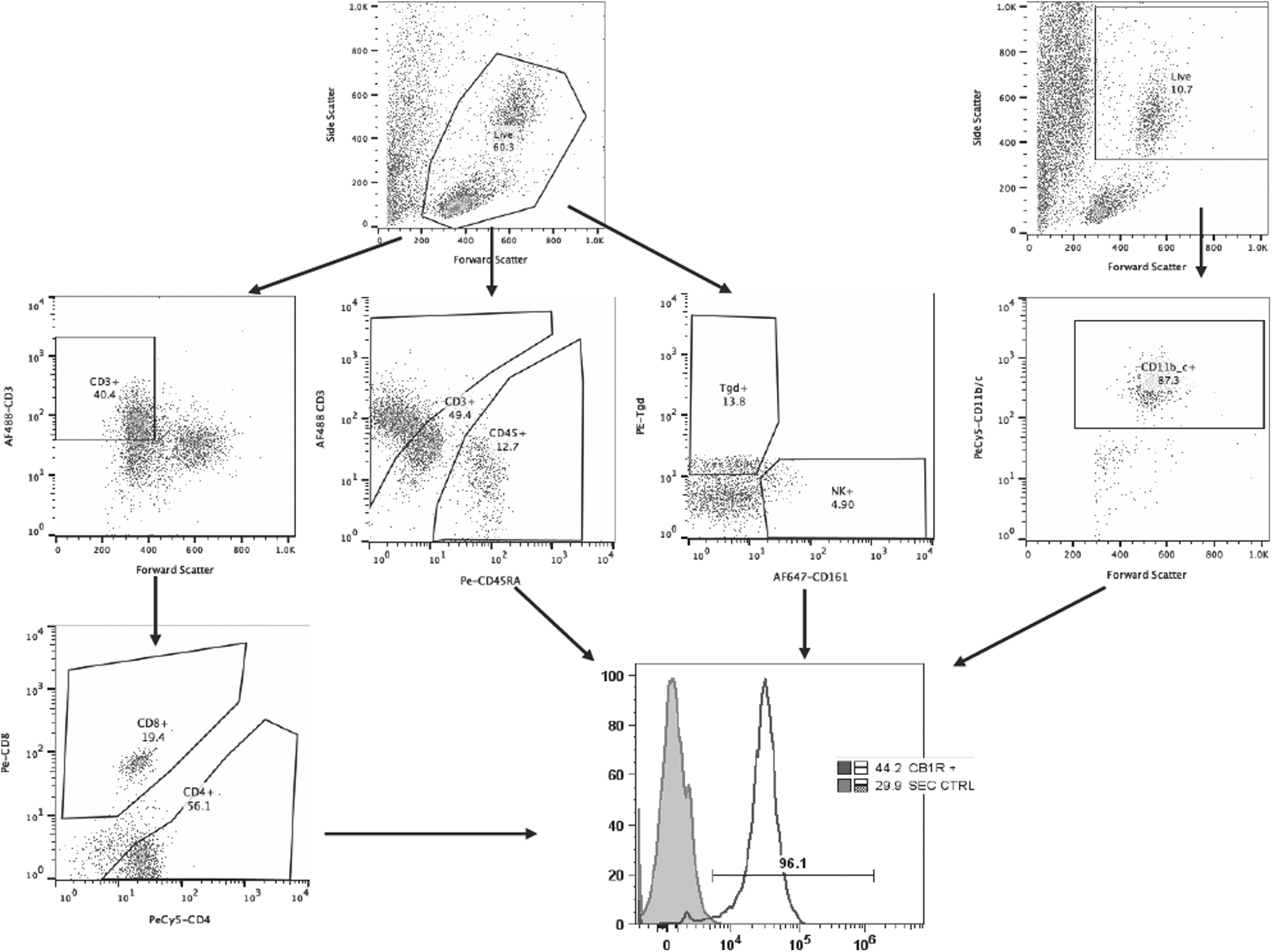
Gating strategy for the selection of immune subpopulations. Viable cells were selected by size and complexity in a FSC versus SSC dot plot. T Lymphocytes were selected, first, by the expression of CD3 complex and then by the presence of CD4 and CD8 coreceptors. B cells were gated according to the expression of CD45RA, and CD3 was included in this mix to get the T cells/B cells ratio, Tγδ- and NK cells were stained in a third mix, and NK46 and Tγδ antibodies were used, respectively. Finally CD11b/c was used to identify the monocyte/granulocyte subpopulations. CB1 staining was included in each mix pf antibodies and analyzed for each immune cell subpopulation. NK, natural killer.
Innate immune cell detection by flow cytometry
From the innate immunity, NK cells (CD161+, P = 0.1542) and monocyte/neutrophil (CD11b/c+, P = 0.8514) did not present significant differences across experimental groups (Fig. 2).

Alteration in the proportion of immune cells from the innate response by chronic exercise in blood.
Adaptive immune cell populations detected by flow cytometry
From the adaptive immunity, CD3+ (P = 0.8227) and CD3+ CD8+ (P = 0.0029) presented as a consequence of the chronic exercise. Nevertheless, T helpers (CD3+ CD4+, P = 0.0113) presented a significant reduction in those animals from EXE compared to TC and SED (Figs. 2 and 3). It is noteworthy that CD3+ CD8+ cells were reduced in EXE and TC compared to SED, suggesting a probable effect from the exercise treadmill and not from the exercise per se (Fig. 3).
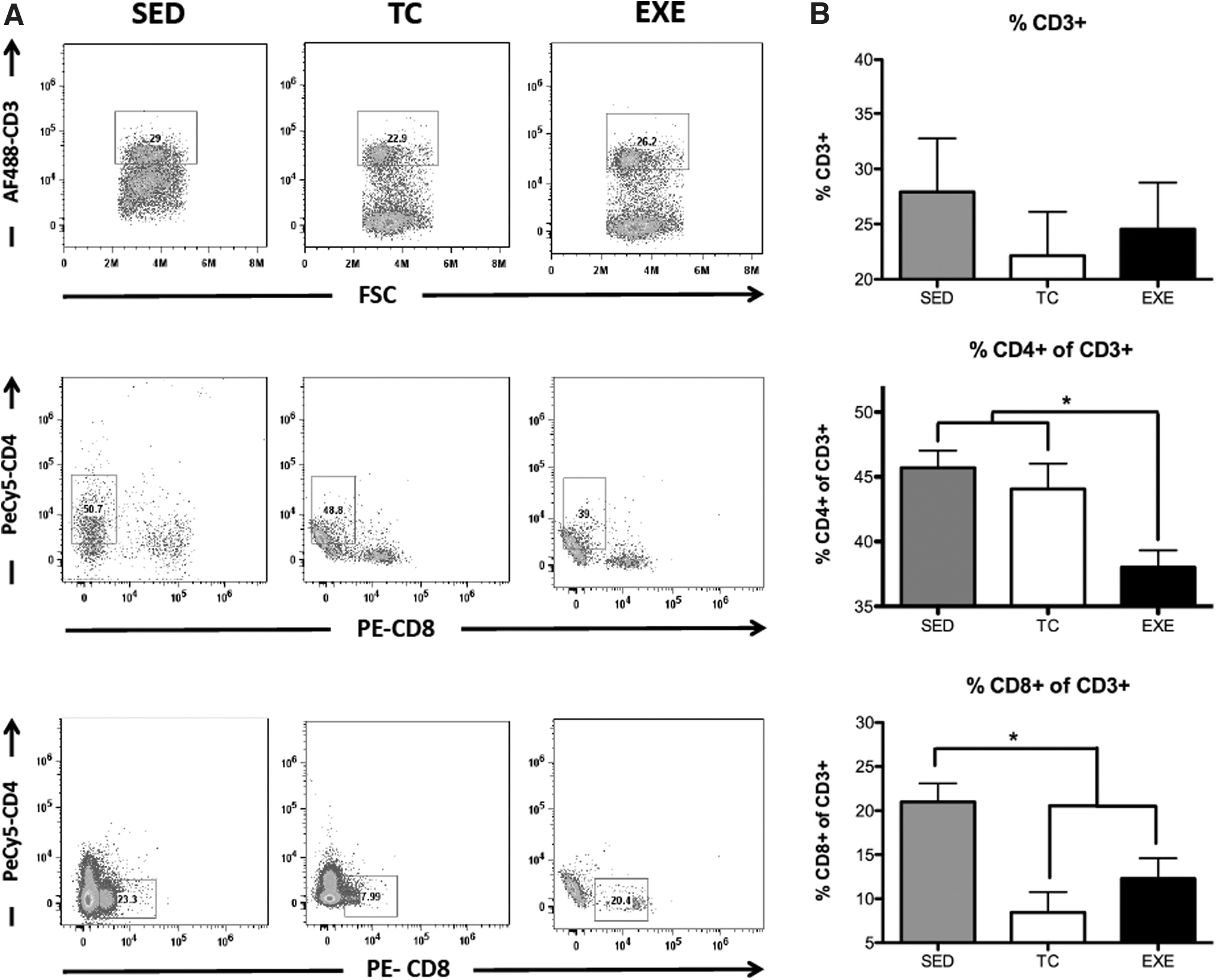
Changes in the proportion of circulating T lymphocytes by chronic exercise.
Tγδ and CD45 RA+ lymphocytes detected in circulation by flow cytometry
It is noteworthy, that Tγδ lymphocytes (P = 0.0020) presented a significant reduction in those animals from EXE compared to TC and SED (Fig. 4). In the case of CD45 RA+ (P = 0.8733) lymphocyte populations, those did not show significant changes across experimental groups (Fig. 4).
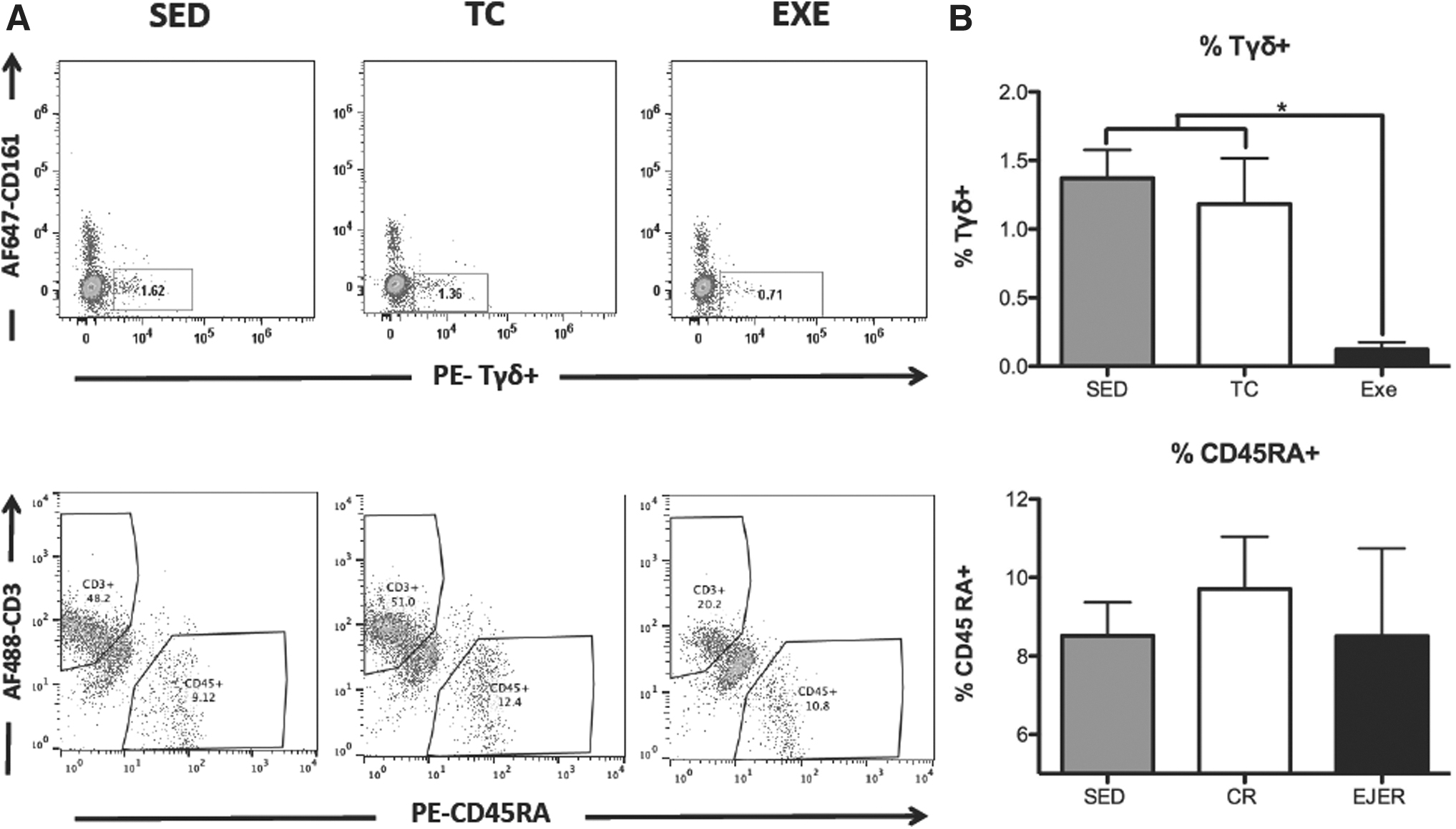
Changes in the proportion of circulating Tγδ and B cells by chronic exercise.
Modulation of CBR1 in PBMC after chronic-moderate exercise
We analyzed changes in CBR1 expression in PBMCs of the innate and adaptive immunity. CD11b+ cells from the innate immunity did not exhibit significant changes in CBR1 expression between the experimental groups. However, CD161+ cells showed an increase in CBR1 expression in EXE compared to TC and SED. Only CD3+ CD4+ cells of the adaptive immunity showed a significant increase in CBR1 expression compared to both control groups. CD3+CD8+, Tγδ+, and CD45RA+ cells did not show significant variation in CBR1 expression between experimental groups due to exercise. Notably, CD3+ cells showed a significant reduction in CBR1 expression in EXE and TC compared to SED, suggesting an effect unrelated to the exercise itself (Fig. 5).
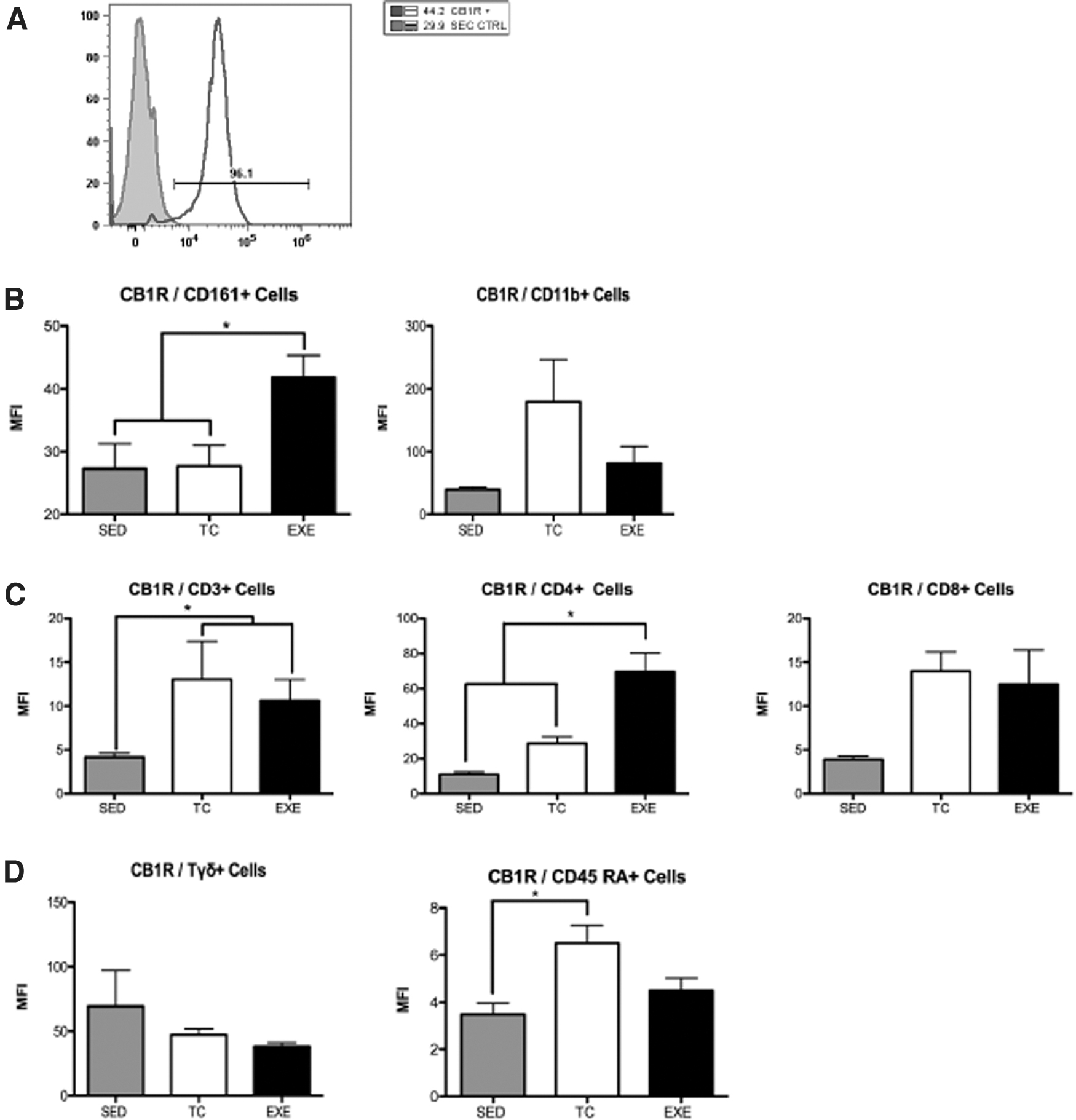
Expression of CBR1 in PBMC's of chronically EXE rats.
Modulation of circulating cytokines in EXE rats
Modulation of cytokine levels has been observed after acute bouts of exercise. We therefore determined the concentration of key cytokines in serum samples from chronically EXE rats and control experimental animals (Fig. 6). Animals from EXE exhibited significantly increased levels of proinflammatory cytokines, such as IL-1β (EXE 216 ± 69, TC 55 ± 6, SED 77 ± 11, P < 0.0001), IFN-γ (EXE 322 ± 35, TC 228 ± 25, SED 226 ± 14, P = 0.0098), or TNF (EXE 9 ± 2, TC 4.8 ± 0.44, SED 4.8 ± 0.6, P < 0.0001). Anti-inflammatory or regulatory cytokines such as IL-4 (EXE 83 ± 11, TC 46 ± 5, SED 55 ± 4, P = 0.0011), IL-5 (EXE 183 ± 29, TC 142 ± 13, SED 117 ± 2, P < 0.05), and IL-10 (EXE 303 ± 70, TC 131 ± 35, SED 125 ± 18, P < 0.0001) were also found increased in group EXE. While IL-13 and TGF-β1 displayed a slight decrease in EXE subjects, the change was not statistically significant. IL-6 was undetectable in all samples examined (Fig. 6).
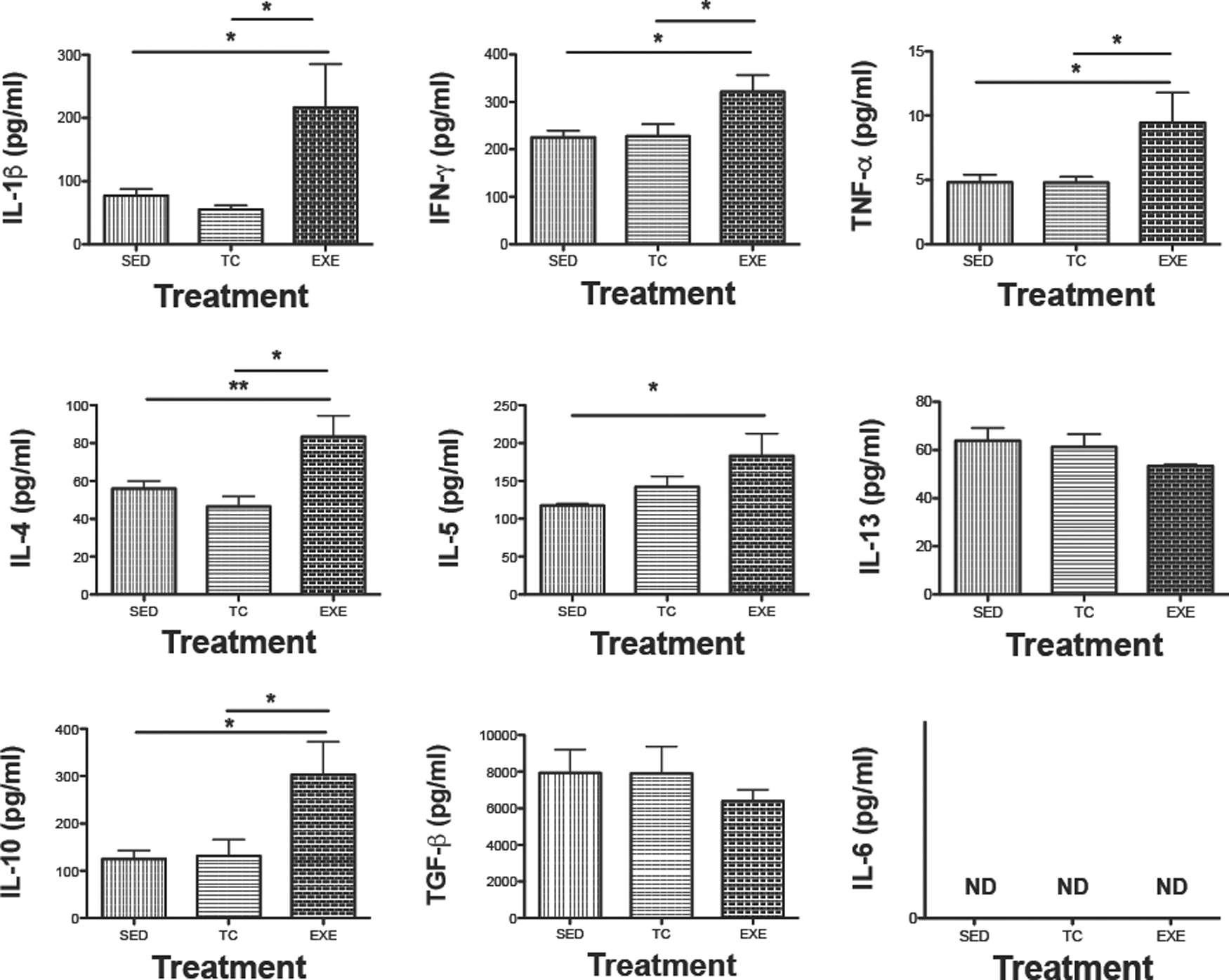
Modulation of levels of different cytokines in serum due to chronic exercising. Determination of indicated cytokines with Milliplex kit in serum samples from different experimental groups: SED, TC, or EXE are expressed as mean ± standard error, n = 7. ND, nondetected levels of IL-6 cytokine. *P < 0.05, **P < 0.001. IL-6, interleukin-6.
Corticosterone levels do not differ across experimental groups
Corticosterone levels from blood serum of six rats from each experimental group showed a significant variance (ANOVA, n = 6, P = 0.0473). Nevertheless, Tukey's post hoc analysis failed to reveal a significant difference in the interaction across experimental groups, with mean levels 46.8 ng/mL for SED, 102.6 ng/mL for TC, and 96.5 ng/mL for EXE (not shown).
Discussion
Chronic moderate aerobic exercise has been widely recognized as a crucial component for maintaining good health and fitness (Fleg, 2012; Ruegsegger and Booth, 2018). As SED lifestyles become increasingly prevalent (Gleeson, 2007; Gleeson et al., 2011), exercise has gained popularity and has become a major focus of research in both normal physiological conditions and pathological contexts (GROUP, 2002; Sugiura et al., 2001). In a previous study, we investigated the long-term effects of chronic moderate aerobic exercise on splenocytes in rats, revealing changes in the proportions of immune cell subpopulations from the adaptive immunity, as well as the expression of CBRs in these cells, and an enhancement in NK cell proliferation (Valencia-Sanchez et al., 2019). Building upon these findings, our current study aimed to expand our understanding of the effects of the same exercise paradigm on PBMCs.
Consistent with previous research from our group, we observed a reduction in the proportion of T helper (CD4 + ), and Tγδ cells were affected by our exercise paradigm, presenting a reduction in EXE compared to TC and SED. Similarly, to what has been previously reported by our group, these two immune cell subpopulations are also affected in the spleen of EXE rats. These changes, although counterintuitive, further support previous reports about the diminution of cellular components in athletes undergoing intense training programs (Moro-García et al., 2014), opening the question of what the relationship between a health enhancement and a diminution of cellular components of the immune system might be.
Studies utilizing similar exercise paradigms have demonstrated that exercise-induced effects on the immune system extend beyond changes in cellular composition (Gustafson et al., 2017; Saito et al., 2003; Shimizu et al., 2008) and encompass enhanced interleukin production in vitro (Lancaster and Febbraio, 2014), cellular proliferation, cytotoxic activity, and phagocytosis (Murphy et al., 2004; Radom-Aizik et al., 2013; Syu et al., 2012). In our study, we quantified pro- and anti-inflammatory interleukins and found increased levels of IL-1β, IFN-γ, and TNF in EXE animals, similarly to IL-10 and IL-4. Notably, IL-6 was not detected in any of the analyzed samples, and unfortunately the amount of blood serum was not enough to repeat the experiment, but it might indicate that this particular cytokine is not circulating if there is not an insult and might prevent chronic inflammatory conditions.
The increase in interleukins in animals from EXE cannot be interpreted as an acute response to the exercise, since the animals were allowed to rest for 1 day after the last bout of exercise, and as previously described, the transient increase of IL-6 and the so-called myokines return to normal levels after 1 h (Gleeson et al., 2011).
These data suggest an increased release and production of interleukins systemically, regardless of their specific response profiles (Th1 or Th2). Elevated amount of IFN-γ and IL-4 is associated with detrimental effects of excessive physical activity and are considered a major risk factor for allergic disorders. Conversely, an increase of IL-1ß and IL-8 might contribute to an overactive immune elements, by increasing mobility of immune cells and activating Th1 monocytes/macrophages. These findings are in line with previous studies that have demonstrated the complex interplay between exercise and immune regulation. While regular moderate aerobic exercise is generally associated with improved health and immune function, excessive or intense exercise can lead to immune dysregulation and increased susceptibility to certain conditions. The precise balance between immune activation and regulation is crucial for maintaining optimal immune responses and preventing immune-related disorders.
As reviewed by Docherty et al., determining a true “resting” cytokine profile creates a logistic issue. The subject's cytokines profile changes with regular exercise, nonetheless to what extent remains debated (Docherty et al., 2022). The benefits provided by chronic moderate exercise are reflected in the immune response, the immune cells involved are mostly monocytes and lymphocytes subpopulations, where the Th1/Th17 ratio and Th2 balance not only in proportion but also in effector molecules released are important to control inflammation, which ultimately could lead to chronic diseases. In homeostasis, proinflammatory cytokines such as TNF-α, IL-1β, and IFN-γ produced by macrophages and lymphocytes are required to perform physiological functions from response to foreign invaders, bone remodeling, and so on. On the contrary, moderate exercise stimulates Th2 response where lymphocytes produce IL-4 and IL-10 cytokines necessary to control and modulate these inflammatory cytokines to reach homeostasis.
In our study, we were not capable of determining the levels of IL-6, an important myokine released by skeletal muscles immediately after acute exercise. This result concurs with previous reports of its transient increase after exercise, followed by a drastic decrease. These low values probably reached levels below our kit's detection threshold. However, other cytokines follow different temporal dynamics, and an increase of pro- and anti-inflammatory cytokines has been extensively reported after an intense quote of exercise (7).
In this protocol, animals were terminally bled 24 h after the last exercise session, and a combination of pro- and anti-inflammatory cytokines was found to increase only in the EXE group, suggesting that this change can persist for longer periods (IL-1β, TNF-α, IFN-γ, IL-4, IL-5, and IL-10). It is a limitation of our protocol to not have included different and longer terminal points, which could have allowed an accurate assessment of the rectification of these IL levels.
Data from this study showed an increase in the expression of CBR1 in T helper (CD4 + ) and Tγδ cells in EXE animals. This result is consistent with previous data obtained from the same cell subpopulations in the spleen of EXE rats (Valencia-Sanchez et al., 2019). Staining for CBR2 was also performed, but unfortunately, poor quality was observed, which prevented reliable results in blood samples, contrary to what was reported previously in the spleen. To our knowledge, this work is one of the first to assess the expression of CBR's through flow cytometry. Improvement in the staining technique will be necessary when performing this analysis in blood samples due to the optical interference of erythrocytes debris.
Above all, the increase of CBR1 observed in T helper (CD4 + ) and Tγδ cells from EXE animals suggests an enhancement in the modulatory requirement of those cells, as it has been demonstrated that activated cells increase their expression of CBR's (Klein et al., 2003; Tanasescu and Constantinescu, 2010), and even higher expression is found in proinflammatory cells (Bouaboula et al., 1993; Chiurchiu et al., 2015).
Finally, we would like to address those changes that are not attributable to exercise, meaning the differences observed in EXE and TC compared to SED. For instance, the population of cytotoxic t cells (CD8 + ) is reduced in both experimental groups exposed to the treadmill in operation, suggesting a probable effect of stress. Although our analysis of corticosterone levels did not show significant differences across the experimental groups, it is known that different immune cell subpopulations present different sensitivity to stress molecules, including other glucocorticoid hormones (Hoffman-Goetz and Pedersen, 1994; Shaw et al., 2017).
The expression of CBR1 in CD3 + cells was also increased on EXE and TC compared to SED, once again suggesting an effect related to manipulation and the exposure to the treadmill. It has been demonstrated that T cells can be activated by glucocorticoids, usually driving those cells toward Th2, which in turn increases the expression of CBR's in their surface (Strehl et al., 2019).
Even though our team is concerned about providing an understanding as broad as possible of the effects that chronic exercise has on the immune system. In this work, we decided to focus exclusively on male rats for methodological reasons. In this work, we assessed the effects of chronic exercise, which demanded training the animals for long periods, by far exceeding the length of the estrous cycle of female rats, affecting behavior, hormone levels, neurotransmitters, and possibly endocannabinoid levels of male rats (Moser et al., 2023; Rodríguez-Manzo and Canseco-Alba, 2023). To diminish possible confounds, we determined that the study of both sexes in this particular matter would be best studied separately, since space and equipment would have been shared.
The results obtained from this research point to the cannabinergic system as a mediator of the immune response and composition during exercise training. Notwithstanding, whether the changes observed in the expression of the CBRs is a primary or secondary response after physical activity, is still a subject to be studied. Our findings point to the relationship between the cannabinoid system and the immune system, as a possible substrate for therapeutic intervention in the future. Taking into consideration that some portion of the population needs the beneficial effects of exercising but finds themselves physically limited, this field of research is a promising area in public health.
The findings of this study have important implications for our understanding of the physiological consequences of chronic moderate aerobic exercise. By expanding our investigation beyond the spleen to peripheral immune system elements, we provide insights into the systemic effects of exercise on immune function. In addition, the involvement of the cannabinoid system as a potential mediator of exercise-induced immune modulation adds a novel aspect to the existing knowledge in this field.
However, it is important to acknowledge the limitations of our study. The specific mechanisms through which the cannabinoid system influences immune function and the underlying signaling pathways require further investigation. In addition, our study focused on a specific exercise paradigm, chronic moderate aerobic exercise, and may not fully capture the effects of other exercise modalities. Future research should explore these aspects to gain a comprehensive understanding of exercise-immune system interactions.
Conclusions
In conclusion, our findings demonstrate that chronic moderate aerobic exercise leads to alterations in peripheral immune cell populations, CBR expression, and cytokine production. These results provide valuable insights into the immunomodulatory effects of exercise and highlight the potential involvement of the cannabinergic system. Further research in this area will deepen our understanding of exercise-induced immune remodeling and contribute to the development of exercise interventions for optimizing immune health.
Footnotes
Acknowledgments
We thank Unidad de Investigación from Facultad de Veterinaria, UNAM for facilitating the use of MAGPIX equipment from LUMINEX by Millipore.
Authors' Contributions
Conceptualization, J.M-.M. and K.E.N-.C.; Methodology, S.V-.S., K.E.N-.C., C.A.G-.C., and V.H.D.R-.A.; Validation, K.E.N-.C. and C.A.G-.C.; Formal analysis, J.M-.M; Resources, J.M-.M.; Writing—original draft preparation, S.V-.S., C.A.G-.C., and J.M-.M.; Writing—review and editing, J.M-.M., S.V-.S., and C.A.G-.C.; Supervision, J.M-.M.; Funding acquisition, J.M-.M. and K.E.N-.C. All authors have read and agreed to the published version of the article.
Data Availability Statement
The datasets generated and analyzed during the current study are included in the present article. Also, they are openly available from the corresponding author on request.
Institutional Review Board Statement
All experimental procedures in the animals were approved by the Institutional Care and Animal Use Committee (Full name: Consejo para el Cuidado y Uso de Animales de Laboratorio), adhering to Mexican regulation (NOM-062-ZOO-1999), and in accordance with the recommendations from the NIH of the United States of America (Guide for the Care and Use of Laboratory Animals).
Animal Use Statement
The present study is reported in accordance with ARRIVE guidelines.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was funded by Grants IN-218922 (KENC) and IN-202723 (JMM) from Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT), Dirección General de Asuntos del Personal Académico (DGAPA), UNAM.