
Abstract
Interleukin 18 (IL-18) is a key cytokine involved in the activation of T and NK cells, which are major effector cells in tumor killing. However, recombinant IL-18 showed limited efficacy in clinical trials. A recent study showed the lack of efficacy was largely due to the existence of IL-18BP, a soluble decoy receptor for IL-18. It was shown that engineered IL-18 variants that maintained pathway activation, but avoided IL-18BP binding, could exert potent antitumor effects. In this study, we demonstrated an alternative strategy to activate IL-18 signaling through direct receptor dimerization. These results provide evidences that the IL-18 pathway can be activated by directly bridging the receptors and, therefore, bypassing the IL-18BP-mediated inhibition.
Introduction
Interleukin (IL)-18
Being such a critical cytokine, the activity of IL-18 is tightly regulated. IL-18 is produced as an inactive precursor and the maturation of IL-18 requires the cleavage by the enzyme CASP1 upon inflammasome activation (Chan and Schroder, 2019). In addition, the function of active IL-18 is balanced by a high-affinity inhibitory IL-18 binding protein, IL-18BP (Wang et al., 2023).
While wild type recombinant IL-18 failed to demonstrate efficacy in clinical studies (Tarhini et al., 2009), IL-18BP evasion mutants were able to exert antitumor effects. In a recent study, the authors demonstrated that IL-18BP binding deficient IL-18 variants were efficacious in multiple preclinical tumor models (Zhou et al., 2020). Although engineering the IL-18 protein is a promising approach to explore for anticancer immunity, it is hindered by the inherent instability of the cytokine as well as potential toxicities related to systemic administration of such a potent molecule.
In this study, we explored whether IL-18 signaling pathway can be directly activated through the dimerization of the IL-18R1 and IL-18RAP chains by taking advantage of the A/C Heterodimerizer system. We expressed chimeric IL-18 receptors fused to dimerization domains (DmrA and Dmrc binding domains). When stimulated with the A/C Heterodimerizer ligand, IL-18 signaling pathway was activated in cells through induced receptor dimerization. These results demonstrate that bringing the two receptor chains in proximity is sufficient for pathway activation and provide an alternative strategy in designing IL-18 therapeutics.
Materials and Methods
Materials
Plasmids containing DmrA/C sequences and the A/C Heterodimerizer ligand were obtained from Takara (Cat No. 635067). pBEF1A _IRES _PGK-Hygromycin plasmids were made in-house and used to make dual receptor expression cell lines. IL-18R1 and IL-18RAP expression fragments fused with DmrC or DmrA, respectively, were cloned into the dual expression pBEF1A backbone and were verified by sequencing. The plasmids used in this study are listed in Table 1.
Dimerization Domain Fused Interleukin-18 Receptor Expressing Plasmids
Cell culture and stable cell line generation
HEK-Blue™ Null-1 cells (Cat No. hkb-null1) and Jurkat-Dual™ cells (Cat No. Jktd-isnf) were obtained from InvivoGen. HEK and Jurkat cells were maintained in DMEM or RPMI medium with 10% fetal bovine serum, 2 mM
For generating stable cell lines expressing chimeric IL-18 receptors, 1.5 × 106 Null-1 Reporter HEK cells or 1 × 106 Jurkat/NK-92 cells were either transfected (JetPrime reagents from Polyplus) or electroporated (Neon transfection system) with a total of 2 μg of DNA consisting of each protein expressing plasmid (Table 1) plus piggyback transposase (pBO vector) in a ratio of 3:1 following manufacturer's protocol. After 72 h, the transfected cells were subcultured in fresh media containing 400 μg/mL hygromycin for selection and expansion.
Antibodies and flow cytometry
Monoclonal antibodies for human IL-18R1 (Clone H44), human IL-18RAP (Cat No. BAF118) and mouse IL-18R1 (Cat No. MAB1216) were obtained from Biolegend, Novus biologicals and R&D systems, respectively. Rabbit polyclonal antibodies were raised in-house against mouse IL-18RAP.
For flow cytometry analyses, 1 × 106 individual cells were stained with anti-human or anti-mouse primary antibodies against IL-18 receptors IL18R1 and IL-18RAP for 1 h followed by 30 min of staining with phycoerythrin-secondary antibody at 4°C. Afterward, the cells were washed, resuspended in FACS buffer, and analyzed using a FACS Attune Nxt 14 Color. The expression analyses were carried out using FlowJo_v10.8.1 software.
Reporter assay
5 × 104 IL-18 chimeric receptor expressing HEK reporter cells or 1 × 105 Jurkat-Dual NF-κB/interferon regulatory factor (IRF) cells were seeded into 96-well plates. The cells were stimulated with increasing concentrations of the A/C Heterodimerizer ligand (Cat No. 635055). IFNa obtained from Peprotech was used to treat the Jurkat Hu-N cell line at a concentration of 5 × 104 U/mL. After overnight incubation, the reporter activities were detected through Quanti-Blue assay for HEK-blue cells and Quanti-Luc/Blue assay for Jurkat cells following manufacturer's protocol (InvivoGen).
Homogeneous time-resolved fluorescence assay
IL-18 pathway activation in NK-92 cells were measured by detecting the release of IFNγ in the supernatant. Before measuring IFNγ, detection antibodies IFNγ Eu cyprate antibody and IFNγ XL antibody from the Human IFNγ homogeneous time-resolved fluorescence (HTRF) Kit (PerkinElmer) were diluted and mixed together in detection buffer following the manufacturer's instruction (Cat No. 62HIFNGPEG). Four microliters of the antibody mixture was combined with 16 μL of NK-92 cell culture supernatant in a white 384-well low volume plate (Cat No. 784075; Greiner Bio-One).
The plate was sealed and incubated overnight at room temperature. HTRF signals were recorded using a PHERAstar microplate reader. The ratio of the acceptor and donor emission signals was calculated for each well using the following equation: (
Results and Discussion
Construction of stable cell lines expressing the chimeric IL-18 receptors
There has been significant interest in exploring cytokine-independent receptor dimerization for pathway activation. Different approaches were explored, including the use of diabodies (Moraga et al., 2015), leucine zipper domains (Nakase et al., 2012) and fluorescent proteins (Engelowski et al., 2018). In this study, artificial IL-18 receptor dimerization was induced by fusing the dimerization domains (DmrA and DmrC) to the receptors and supplementing the dimerization inducer, the A/C Heterodimerizer ligand.
The plasmids expressing both the IL-18R1/DmrC and IL-18RAP/DmrA chimeras (N- or C-terminus dimerization domain fusion) were transfected into HEK Null-1 NF-κB reporter cells to make stable cell lines (Fig. 1a). The activation of the IL-18 pathway can be detected using the Quanti-Blue assay (InvivoGen), which measures the activity of the secreted embryonic alkaline phosphatase (SEAP) in an absorbance assay.
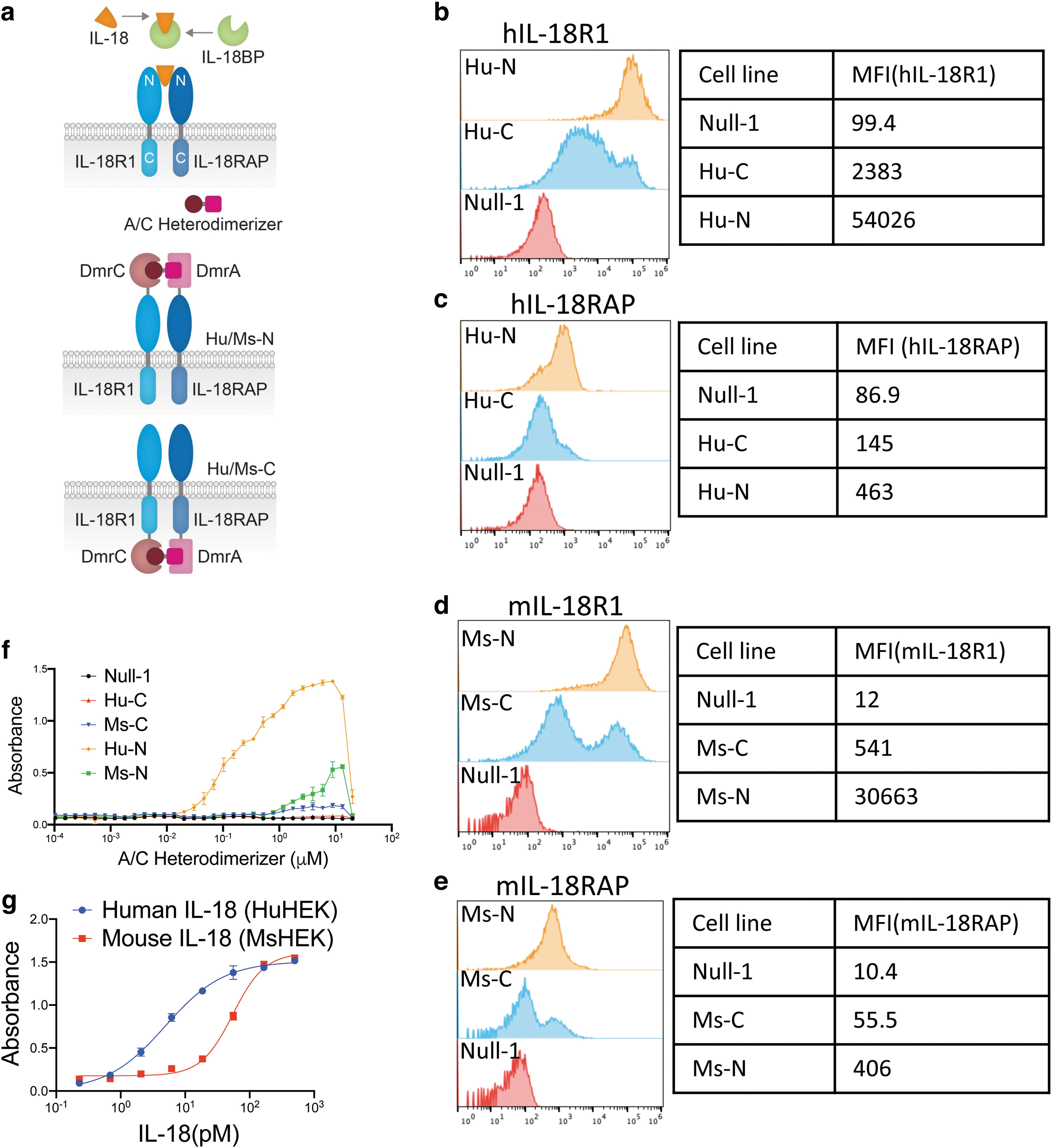
Construction of reporter cell lines expressing IL-18 chimeric receptors and functional assessment of IL-18 pathway activation in HEK cells.
Cell lines with either human or murine IL-18 receptor chimeras were made and the N or C terminal fusion were designated as Hu/Ms-N or Hu/Ms-C, respectively. The expression of these receptors was verified using flow cytometry (Fig. 1b–e). Dimerization domains fused at the N-terminus showed better protein expression in both species (Fig. 1b–e). For human IL-18RAP, we can hardly detect its expression when its C-terminus was fused to DmrA (Fig. 1c). These data demonstrated that the receptor proteins did not tolerate C-terminal fusions very well.
Functional assays to test the IL-18 signaling activation
IL-18 receptors are commonly activated upon binding of the IL-18 cytokine. This process starts by the binding with the low-affinity receptor IL-18R1 and is followed by the recruitment of IL-18RAP (Yasuda et al., 2019). This resulting ternary complex then activates downstream signaling, including NF-κB (Yasuda et al., 2019). To test whether we could activate the IL-18 pathway by bridging the IL-18 receptors through chemically induced domain heterodimerization, the culture medium of the stable cell lines was supplemented with an increasing dose of the A/C Heterodimerizer.
As measured by the NF-κB reporter activation assay, a robust dose–response was detected in stimulated Hu-N cell line (Fig. 1f). Similar but weaker activation was detected in cell lines expressing the murine chimera. For cell lines expressing Hu-C, we barely detected any activity, which might be due to the failure of the expression of the chimeric IL-18RAP (Fig. 1f). To explore why significantly higher activity was observed for the cell line expressing Hu-N compared with the one expressing Ms-N, we compared the response of cell lines expressing the wild-type IL-18 receptor.
We found that the cell line expressing the human IL-18 receptors was 10-fold more sensitive to syngeneic IL-18 stimulation than the one expressing the murine receptors (EC50 5 pM versus EC50 55 pM, for the human and mouse system respectively), suggesting such difference might be an intrinsic variance between the two species or due to the compromised activity of murine receptors when expressed in a human cell line (Fig. 1g).
At concentrations above 8 μM, a hook effect was detected. This was expected as oversaturating the chimeric receptors with the A/C Heterodimerizer ligand would result in the compound binding to the receptor chains individually instead of serving as a domain dimerization bridge.
To test whether we could detect IL-18 independent pathway activation in physiologically relevant cells, Jurkat and NK-92, human T and NK cell lines, respectively, were selected as parent cell lines to make similar chimeric receptor expressing stable cell lines. Based on the results obtained in HEK cells (Fig. 1f), we only constructed the cell lines expressing Hu-N. The expression of the IL-18 chimeric receptors was validated by flow cytometry (Fig. 2a, b).
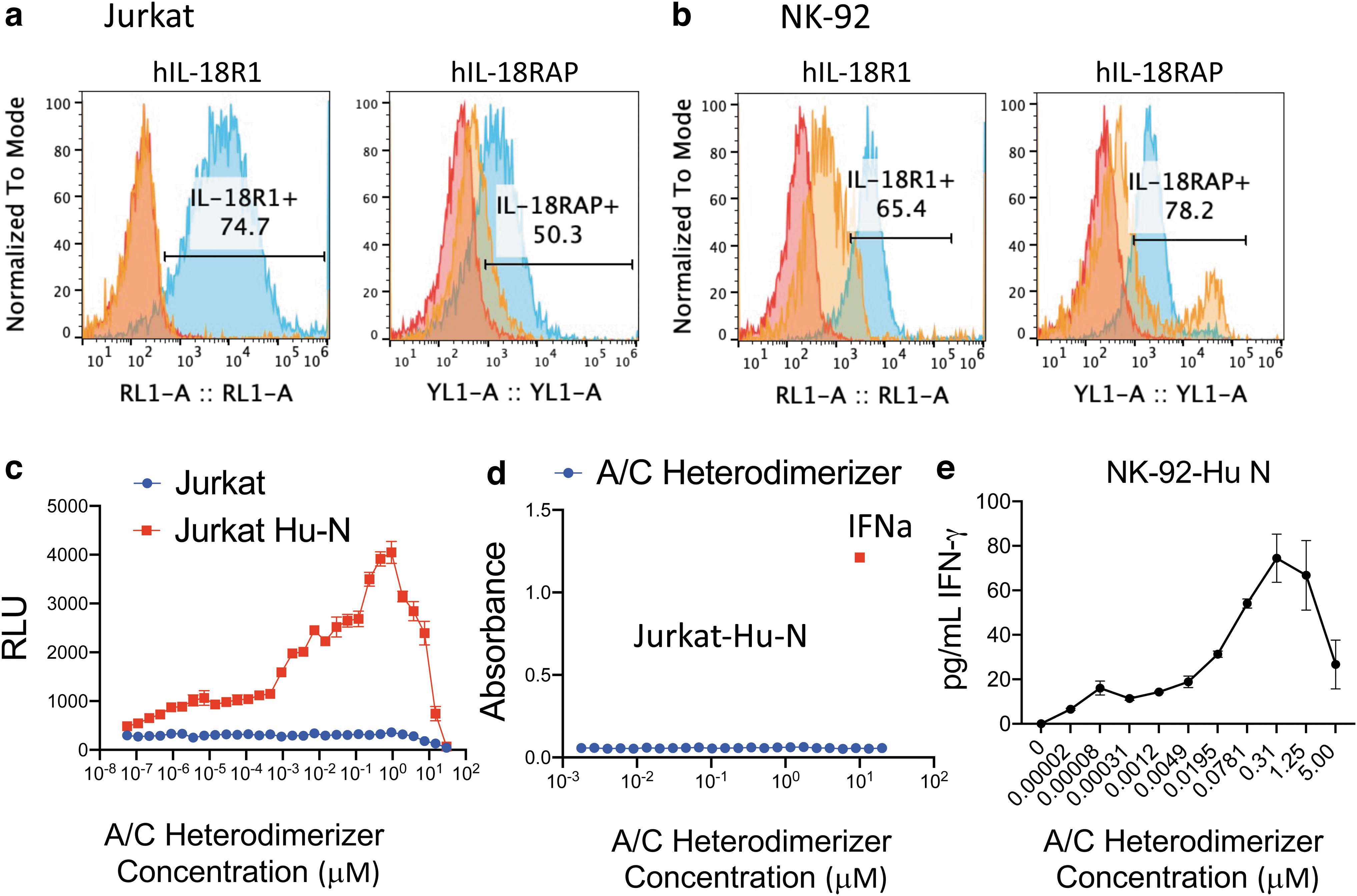
Construction of IL-18 chimeric receptor cell lines and functional assessment of IL-18 pathway activation in Jurkat and NK-92 cells.
Both the IL-18R1 and the IL-18RAP receptors were expressed in these stable cell lines and their expression levels were significantly higher than the endogenous IL-18 receptors in the parent cell lines, shown as the blue and orange histograms, respectively (Fig. 2a, b). The Jurkat cell line contains dual reporters, which enable us to detect the activation of the NF-κB pathway by the inducible NF-κB responsive Lucia luciferase in the Quanti-Luc assay and the interferon pathway by the IRF-inducible SEAP in the Quanti-Blue assays (InvivoGen).
For NK-92 cells, IFNγ was detected from the supernatant of the treated cells in HTRF assays. As shown in Fig. 2c and e, we detected dose-dependent activation by adding the A/C Heterodimerizer in these engineered cell lines by measuring NF-κB reporter activity or IFNγ secretion directly. To further confirm the DmrA/C system does not activate other receptors nonspecifically, we tested whether the A/C Heterodimerizer could activate the interferon pathway in the Jurkat Hu-N cell line. In Fig. 2d, we could detect robust interferon pathway activation upon IFNa treatment but not by supplementing A/C Heterodimerizer ligand, demonstrating the DmrA/C system has no effect on the interferon receptors.
Compared with the HEK cells, we noticed that the Jurkat and NK cells were much more sensitive as we started to detect activity at low picomolar concentrations of the compound. Similarly, we detected the hook effect at higher compound concentrations. Overall, these results demonstrated that the IL-18 signaling pathway can be directly activated by receptor dimerization, even in the absence of IL-18.
Conclusion
In this study, we demonstrated an alternative approach to activate the IL-18 signaling pathway by artificial receptor dimerization, which did not require any IL-18 supplementation and was not subjected to IL-18BP inhibition. A recent study showed the activation of the IL-18 pathway by using bispecific antibodies against IL-18R1 and IL-18RAP (Lipinski et al., 2023). Such IL-18 independent pathway activation paves the way for the development of molecules artificially dimerizing the two IL-18 receptor chains, which can be used in IL-18 immunotherapy.
Footnotes
Acknowledgments
We thank Tao Sun for introducing us to the A/C Heterodimerizer system.
Authors' Contributions
Y.M. and R.H performed and analyzed the experiments. T.M. and I.L. designed and provided all the constructs used in this study. Conception by M.M., J.S., N.W., B.B., and Y.Z. Y.Z. supervised the study, data analysis and interpretation, and wrote the original draft; all authors participated in the revision of the article.
Author Disclosure Statement
All authors are or were employees of Genentech, a member of the Roche group, and may hold Roche stock or stock options.
Funding Information
This work was supported by Genentech, Inc.