
Abstract
Psoriasis is a chronic, immune-mediated inflammatory skin disease characterized by epidermal thickening and inflammatory cell infiltration. Excessive proliferation of keratinocytes and resistance to apoptosis lead to thickening of the epidermis. Plasmacytoid dendritic cells are involved in the occurrence of psoriasis mainly by secreting interferon-alpha (IFN-α). IFN-α is a glycoprotein with antiviral, antitumor, and immunomodulatory effects, but its role in psoriasis remains unclear. In this investigation, a mild psoriatic phenotype was observed in mice upon topical application of IFN-α cream, and the inflammation was exacerbated when combined with imiquimod (IMQ). Immunohistochemical analyses demonstrated that IFN-α induces psoriatic inflammation in mice by stimulating phosphorylation of forkhead box O3, consistent with the involvement of this protein in cell proliferation, apoptosis, and inflammation. Our results suggested that topical IFN-α caused psoriatic inflammation and that the psoriatic inflammation was exacerbated by the combination of IFN-α and IMQ, possibly due to the dysfunction of forkhead box O3.
Introduction
Psoriasis, a common chronic inflammatory skin disease, is characterized by hyperproliferation, incomplete differentiation of epidermal keratinocytes, infiltration of dermal inflammatory cells, and neovascularization. Among immune cells, the role of plasmacytoid dendritic cells as key pathogenic contributors in psoriasis is well-established, and their targeted elimination significantly impedes the progression of psoriasis (Glitzner et al., 2014). The overproduction and excess secretion of interferon-alpha (IFN-α) by plasmacytoid dendritic cells is a key driver of psoriatic inflammation (Nestle et al., 2005).
IFN-α represents a key effector molecule of plasmacytoid dendritic cells in the innate immune response, and it is also a necessary for the initiation of psoriasis. IFN-α, which is a glycoprotein, is also produced by monocytes and lymphocytes after viral infection (Gu et al., 2022). Accordingly, IFN-α has been adapted for use as a drug in the treatment of a variety of diseases, due to its activities in resisting viral infection and interfering with cell proliferation and differentiation. Emerging studies have also shown that IFN-α has immunomodulatory effects, such as its involvement in the pathogenesis of systemic sclerosis and primary Sjogren's syndrome, in addition to its roles in psoriasis (Trutschel et al., 2022).
The importance of generation of IFN-α by plasmacytoid dendritic cells in the early stages of psoriasis has been demonstrated in multiple ways. For example, the levels of expression of IFN-α and of gene products targeted by IFN-α-stimulated signaling have been shown to be significantly increased in the skin samples of patients with psoriasis (Khosravi-Hafshejani et al., 2022). In addition, patients with chronic viral hepatitis or cancer who are treated with human recombinant IFN-α are at risk of developing de novo psoriasis or experiencing an exacerbation of existing psoriasis (Afshar et al., 2013; Tas and Atsu, 2016).
Conversely, blocking of IFN-α signaling has been shown to inhibit the activation and proliferation of T cells, thereby relieving psoriasiform inflammation in murine models (Gui et al., 2016; Nestle et al., 2005). Given the potential role of IFN-α in psoriasis, we hypothesized that topical administration of IFN-α may induce psoriasis-like inflammation and may aggravate inflammation caused by imiquimod (IMQ) application in mice.
In the present study, we confirmed that topical administration of IFN-α induces multiple molecular and histological responses that characterize psoriatic lesions, such as increased expression of psoriasis-related keratins and cytokines. The production of these keratins and cytokines were more pronounced when combined with IMQ. Furthermore, we also determined that IFN-α causes excessive proliferation of epidermal keratinocytes by enhancing the phosphorylation of FOXO3. Thus, we determined that the innate immune factor IFN-α primes an inflammatory response in the skin that has many features of psoriasis.
Materials and Methods
Preparation of creams
Powdered IFN-α (2 mg; MedChemExpress, NJ) was dissolved in ultrapure water. The mixture was subjected to ultrasonic treatment until the drug was dissolved and the solution was transparent. The IFN-α solution was added into 12.5 g matrix cream (Jinbei Chemical Co., LTD, Zhengzhou, China) or 12.5 g 5% IMQ cream (Sichuan Mingxin Pharmaceuticals Co., China), stirred at high speed with a mini high-shear dispersing emulsifier (FLUKO, Shanghai, China) and cooled at room temperature to yield IFN-α and mixed IFN-α and IMQ creams, both containing 2 × 105 U/g IFN-α.
Mice
Animal experiments were conducted using 8-week-old BALB/c female mice (weight: 20–25 g; SPF-grade) purchased from the Medical Animal Center of Xi'an Jiaotong University (Xi'an, China). This study was approved by the Institutional Animal Care and Research Advisory Committee of the institution.
The mice were randomly divided into 4 groups: a control group, in which the mice were treated with the matrix cream; an IFN-α group, in which the mice were treated with IFN-α cream; an IMQ group, in which the mice were treated with 5% IMQ cream; and an IFN-α+IMQ group, in which the mice were treated with the cream containing both IFN-α and IMQ. A 2 × 2 cm area on the back of each animal was shaved 2 days before modeling. The mice were administered 62.5 mg cream/day on the back for 7 days. Morphological changes in skin, and body weight were recorded daily. Erythema, scaling, and thickness were assessed based on the psoriasis area and severity index scoring system: 0, none; 1, slight; 2, moderate; 3, severe; and 4, very striking (Peng et al., 2018). On day 8, the mice were sacrificed to collect skin samples.
Immunohistochemical and histological evaluation
Immunohistochemistry was performed according to a standard protocol. Skin tissue was fixed with 4% paraformaldehyde, embedded in paraffin, and sectioned for later use. Following a 1 h incubation with 2.5% normal goat serum in phosphate-buffered saline, the sections were first incubated with the primary antibody overnight at 4°C and then with the secondary antibody for 30 min at room temperature. All sections were imaged with a Nano Zoomer 2.0 HT slide scanner (Hamamatsu Photonics, Hamamtsu, Japan). The 2–5 fields were randomly selected from each section and quantified using Image Pro-Plus 6.0 software. The epidermal thickness was measured using hematoxylin and eosin sections.
The primary antibodies and dilutions used were as follows. Anti-Ki67 (1:200, ab16667; Abcam), Anti-Keratin 1 (1:1,000, ab185628; Abcam), Anti-Keratin 10 (1:1,000, ab76318; Abcam), Anti-Keratin 17 (1:100, ab109725; Abcam), Anti-Loricrin (1:100, ab85679; Abcam), Anti-CD3 (1:100, ab16669; Abcam), Anti-Iba-1 (1:1,000, ab178846; Abcam), Anti-Ly6G (1:200, GB11229-100; Servicebio), Anti-FOXO3 (1:100, 10849-1-AP; Proteintech), Anti-Phospho-FOXO3a (Ser253) (1:100, AF1783; Beyotime).
Enzyme-linked immunosorbent assay
Enzyme-linked immunosorbent assay (ELISA) was used to detected the expression of inflammatory factors. Fresh tissue was ground in RIPA lysis buffer (Beyotime, Shanghai, China) to obtain protein lysates. All lysate samples were normalized to the mass of the original tissue. Levels of interleukin (IL)-17A, IL-22, IL-23, or TNF-α in the lysate supernatants were quantified with corresponding ELISA kits (Thermo Fisher Scientific, Waltham, MA) according to the manufacturer's instructions. IFN-α was detected using an ELISA kit purchased from Westang Biotech (Shanghai, China).
Western blotting
Fresh tissues or cell cultures were ground using a homogenizer in RIPA lysis buffer containing protease and phosphatase inhibitors (Beyotime). Protein concentrations were determined using the BCA Protein Assay Kit (Beyotime). Western blotting was performed according to a standard procedure. In brief, equal amounts of total protein were separated on a polyacrylamide gel and transferred to a polyvinylidene difluoride membrane (Merck Millipore, Burlington, MA). The membranes were blocked with 5% nonfat milk, incubated with primary antibodies at 4°C overnight, and then incubated with the secondary antibody. The signals were detected using an ECL kit (Biosharp, Hefei, China). The band intensities were quantified using ImageJ software (National Institutes of Health, Bethesda, MD). The primary antibodies and dilutions used were as follows. Anti-Keratin 1 (1:1,000, ab185628; Abcam), Anti-Keratin 10 (1:1,000, ab76318; Abcam), Anti-Keratin 17 (1:1,000, ab109725; Abcam), Anti-Loricrin (1:1,000, ab85679; Abcam), total protein levels were normalized with rabbit anti-β-actin (1:2,000, 81115-1-RR; Proteintech).
Reverse transcription quantitative PCR
Detailed protocols for reverse transcription quantitative PCR is provided in Supplementary Data S1. The primers used are provided in Supplementary Table S1.
Statistics analysis
Data are presented as the mean ± standard error of mean and were statistically analyzed using GraphPad Prism 8.3 (GraphPad Software, San Diego). Multigroup comparisons were performed using one-way analysis of variance. P of <0.05 was considered to indicate statistical significance.
Results
Topical IFN-α administration confers a psoriatic appearance in a mouse model
To determine whether topical IFN-α application induces skin inflammation along with structural features of psoriasis, IFN-α cream, IMQ cream, and a mixture of the 2 creams were separately applied to the shaved back skin of BALB/c mice for 7 consecutive days. After IFN-α treatment for 3 to 4 days, small scales and mild erythema appeared on the skin, whereas treatment with the IMQ cream alone or the mixed cream treatment resulted in severe erythema, larger scales, and a firmer skin texture (Fig. 1A). Erythema and scales continued to increase until the end of the experiment, accompanied by the shedding of scales. The severity of skin lesions (erythema, scaling, and thickness) was scored on days 0, 2, 4, and 7 based on the psoriasis area and severity index score.

IFN-α induces skin inflammation and aggravates IMQ-induced psoriatic inflammation. BALB/c mice were treated daily with IFN-α cream or IMQ cream or a cream containing a mixture of IFN-α and IMQ on a shaved portion of the back skin.
Compared with the control mice, IFN-α-treated mice exhibited greater erythema, scales, and skin thickening, and the mean psoriasis area and severity index score were statistically higher (Fig. 1B–E). Although the erythema of the skin of mice treated with a mixed cream containing both IFN-α and IMQ was worse than that of the mice treated with IMQ cream (Fig. 1B), there was no significant difference in scaling or thickness (Fig. 1C, D), leading to a lack of statistical difference in psoriasis area and severity index scores (Fig. 1E). Histopathological analyses showed that IFN-α increased the epidermal thickness of mice, and the IFN-α and IMQ combination further increased the epidermal thickness, mainly reflected in thick scales (Fig. 1F, G).
IFN-α administration influences the differentiation of keratinocytes
Because HE staining suggested that the IFN-α-induced increase in epidermal thickness was caused by excessive proliferation of keratinocytes, the expression of Ki67 was examined. The results showed an increased number of Ki67-positive keratinocytes in the skin of mice treated with IFN-α, and more proliferating cells were observed in the skin of mice treated with both IFN-α and IMQ than in those treated with IMQ alone (Fig. 2A, B).
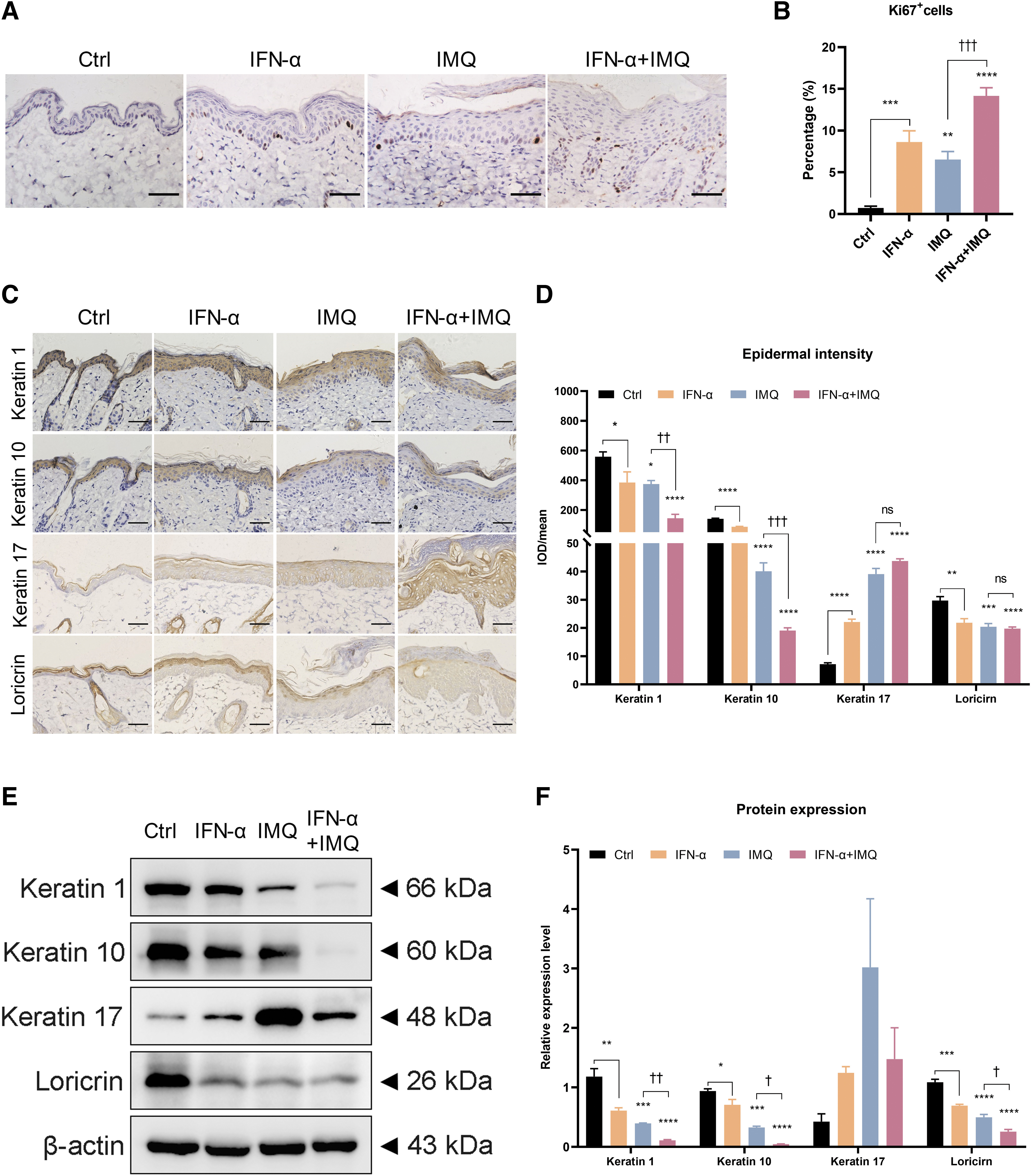
IFN-α results in proliferation and abnormal differentiation of keratinocytes.
Psoriatic skin lesions are characterized by scaling, which is a manifestation of parakeratosis caused by altered epidermal differentiation (Ren and Xia, 2022). In addition, while keratin 1, keratin 10, and loricrin, are distributed in the suprabasal layer in normal epidermis, the levels of these proteins are known to be significantly reduced in psoriatic skin (Donetti et al., 2012; Wang et al., 2022). Therefore, the effects of the treatments on the expression of psoriasis-related keratins were investigated. Immunohistochemical analyses demonstrated that IFN-α treatment correlated with decreased expression of keratin 1, keratin 10, and loricrin, compared with the control skin.
Moreover, the combination of IFN-α and IMQ was more effective in reducing these keratins than was IMQ alone (Fig. 2C, D). The levels of keratin 17, a marker of excessive keratinocyte proliferation in psoriasis, were increased by IFN-α treatment, and the combination of IFN-α and IMQ showed a synergistic effect on this protein (Fig. 2C, D). The results of Western blotting analyses were consistent with those of immunohistochemistry (Fig. 2E, F). Collectively, these results suggested that IFN-α caused excessive proliferation and abnormal differentiation of epidermal keratinocytes.
IFN-α administration increases the infiltration of inflammatory cells
A major histopathological feature of psoriasis is inflammatory cells infiltration, especially involving T lymphocytes, macrophages, and neutrophils. Cytokines secreted by these inflammatory cells stimulate keratinocyte proliferation and promote angiogenesis, further aggravating psoriasis (Liu et al., 2021; Ren and Xia, 2022). In this study, the infiltration of immune cells in the skin on day 7 of treatment was investigated through immunohistochemistry. Markers of activated macrophages (Iba-1), T cells (CD3), and neutrophils (Ly-6G) were used to detect relevant immune cells.
Compared with the control cream, treatment with IFN-α led to significantly increased numbers of CD3+ T cells but had little effect on the presence of macrophages or neutrophils (Fig. 3A–D). The distributions of CD3+ T cells and Iba-1-positive activated macrophages were similar in the skin of mice treated with the IMQ cream or the mixed cream, and the levels of these cells in these treatment groups were higher than that in the control group (Fig. 3A–C). Neutrophil accumulation under the stratum corneum was observed in the skin of both IMQ-treated mice and mice treated with the mixed cream, but the number of neutrophils in the mixed cream group was higher than that in the IMQ group (Fig. 3A, D).
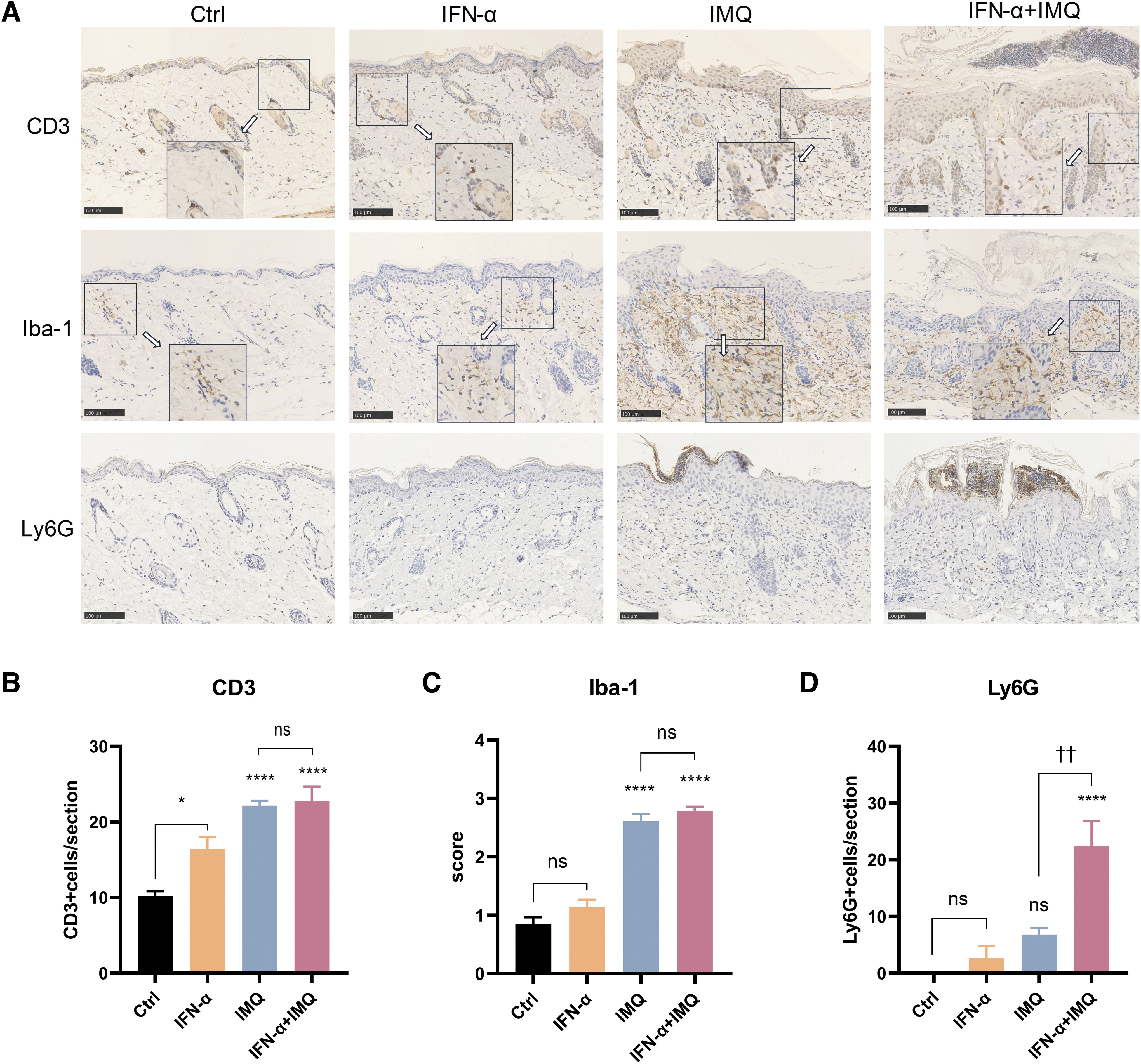
IFN-α treatment induces the accumulation of inflammatory cells.
IFN-α administration enhances the expression of psoriatic cytokines
T cell-mediated immune responses are central to the pathogenesis of psoriasis and involve various cytokines, including IL-17, IL-22, and IL-23 (Gu et al., 2022; Ren and Xia, 2022). Therefore, we quantified the levels of T cell-associated cytokines in tissue lysates using and ELISA-based strategy to determine whether IFN-α could induce the accumulation of inflammatory factors in psoriatic skin. The protein level of IFN-α in the supernatant of skin tissue lysates from IFN-α-treated mice was significantly higher than that observed for the control group (Fig. 4A). There was no significant difference in IFN-α levels in the IMQ group and the combined treatment group (Fig. 4A).

IFN-α treatment leads to the release of inflammatory factors.
Compared with the control group, the protein levels of IL-17A, IL-22, and IL-23 were not significantly changed in the IFN-α group, whereas TNF-α expression was significantly increased (Fig. 4B–E). Although IL-17A, IL-22, IL-23, and TNF-α levels were increased in the skin of mice treated with both IFN-α and IMQ compared with IMQ treatment alone, these differences were not statistically significant.
IFN-α administration exerts its proinflammatory effect through FOXO3
Four members of the forkhead box O family (FOXO) of proteins, specifically FOXO1, FOXO3, FOXO4, and FOXO6, play important roles in inflammation, cell proliferation, tumor formation, and longevity (Santos et al., 2023). Specifically, abnormal proliferation of keratinocytes in psoriasis has been shown to be associated with downregulation of FOXO1 (Aldabbas et al., 2023; Liu et al., 2011). Therefore, we examined the effects of the treatments on the expression levels of the 4 FOXO types in mouse skin tissues, first at the transcriptional level. Here, the expression of mRNA encoding all 4 types of foxo was found to be significantly decreased in IFN-α treated skin tissues (Supplementary Fig. S1). In addition, combined treatment with IFN-α and IMQ led to a significantly stronger decrease of the expression of foxo mRNA than did treatment with IMQ alone (Supplementary Fig. S1B).
Because we observed that IFN-α upregulated the level of inflammatory factors and Keratin 17 (Fig. 4), we speculated that IFN-α might upregulate inflammation and cell proliferation through its effects on FOXO3. Therefore, the expression of FOXO3 and phosphorylated FOXO3 proteins were examined by immunohistochemistry. We observed that phosphorylated FOXO3 was mainly distributed in the cytoplasm, and more of the protein was present in cells from IFN-α-treated mice compared with the control group, although the increase did not rise to the level of statistical significance (Fig. 5A, C). Compared with treatment with IMQ alone, treatment with the combination of IFN-α and IMQ resulted in more pronounced expression of phosphorylated-FOXO3 (Fig. 5A, C).
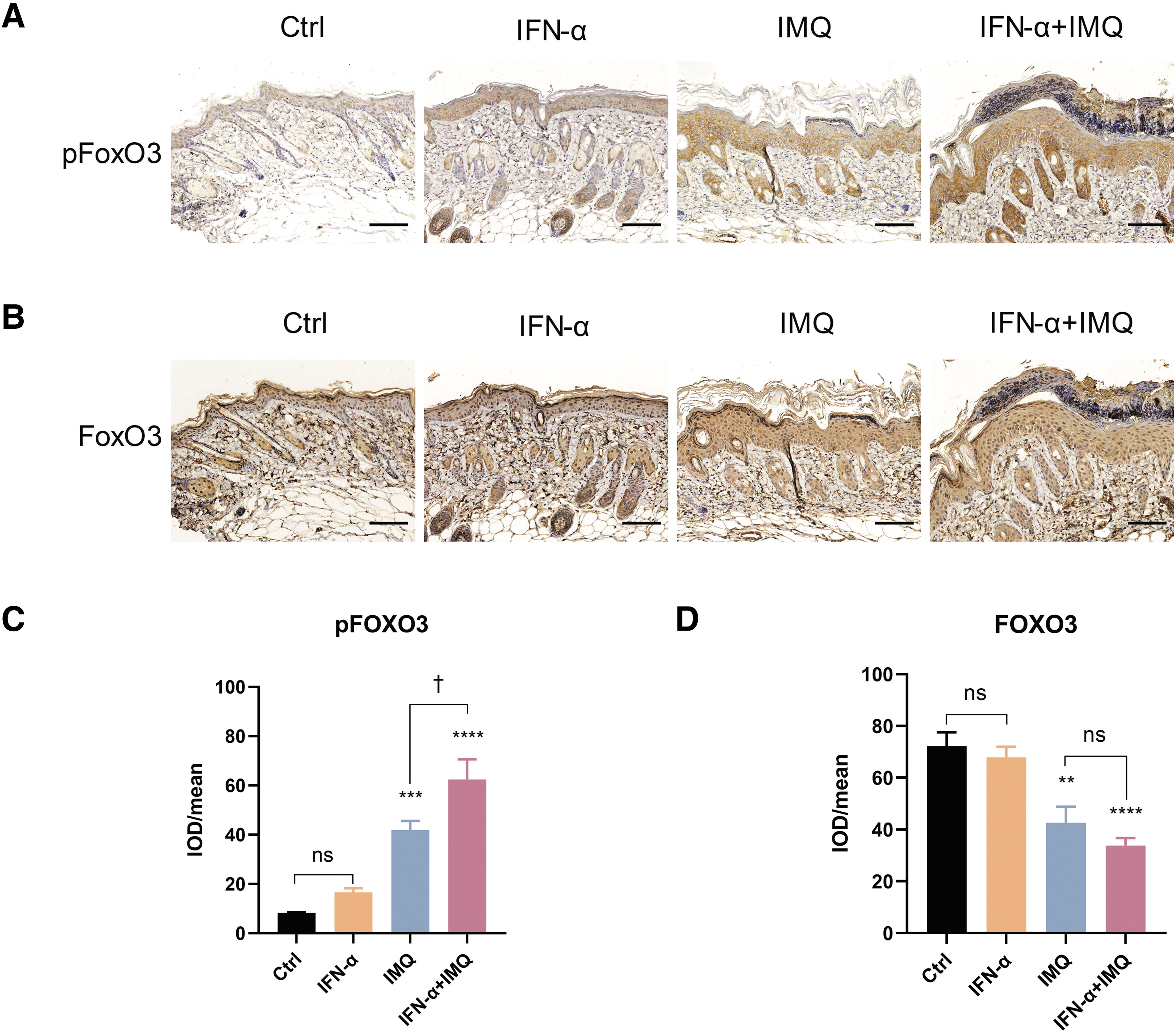
Increased expression of FOXO3 and phosphorylated FOXO 3 in mouse dorsal skin.
The FOXO3 was mainly localized in the nucleus, and there was no significant difference between the control group and the IFN-α group (Fig. 5B, D). The staining representing nuclear-localized FOXO3 was weaker in the IMQ group and the mixed cream group. Moreover, FOXO3 staining was also observed in the cytoplasm.
Discussion
This is the first study to investigate the role of IFN-α in psoriasis by applying IFN-α alone or in combination with IMQ on the depilated skin of mice. We demonstrated that IFN-α alone induced an inflammatory response that was observed at multiple levels and that mice cotreated with IFN-α and IMQ exhibited a more severe inflammation. Macroscopically, the skin of mice in the IFN-α group had light red plaques with small scales, while the erythema in the combined treatment group was mainly dark red with more adherent scales that were not readily shed. The combination treatment was found to lead to increased proliferation of Ki67-positive keratinocytes and stronger immune invasion, in particular neutrophils; these changes were accompanied by significantly decreased levels of keratin 1, keratin 10, and loricrin. The expression of TNF-α, as detected by ELISA, was notably increased.
In short, our results suggest that IFN-α exacerbates IMQ-induced psoriatic inflammation by interfering with keratinization and by promoting inflammatory cell infiltration and cytokine expression. We also determined that the proinflammatory and proproliferative effects of IFN-α may be related to FOXO3 dysfunction.
The main purpose of our experiment was to observe whether IFN-α can act on the skin to cause psoriasis-like inflammation and the effect on IL-17/IL-23; therefore, we used the most widely used mouse model of psoriasis (Gangwar et al., 2022), in which IMQ treatment mimics the local inflammation of human psoriasis in an IL-17/IL-23-dependent manner (van der Fits et al., 2009). Another benefit of this model was that topical application reduced the impact of systemic inflammation and facilitated the observations of the effect of IFN-α on the skin. As expected, we found that IFN-α induced inflammatory responses and aggravated IMQ-induced psoriasis-like inflammation. We also recognize, however, that psoriasis is a systemic disease. Thus, in future experiments, we will investigate the role of the systemic use of IFN-α, for example through intraperitoneal or intravenous injection, in a mouse model of psoriasis.
Type I IFNs signaling pathways are activated in psoriasis and in IMQ-induced models (Nakao et al., 2020; Saiga et al., 2022; Zhang, 2019), as demonstrated in our experiments. Moreover, IFN-α can induce de novo psoriasis or an exacerbation of existing psoriasis during the treatment of nonpsoriatic diseases (Afshar et al., 2013; Li et al., 2022; Tas and Atsu, 2016). Consistent with these findings, in our study, we found that topical IFN-α application also induced skin inflammation and aggravated psoriatic dermatitis in model mice. Activated plasmacytoid dendritic cells, which product IFN-α, are among the factors driving psoriatic inflammation. IMQ induces plasmacytoid dendritic cells to produce proinflammatory mediators such as IFN-α through TLR7 (Ueyama et al., 2014). IFN-α strengthens the expression of the IL-22 receptor and CXC motif chemokine ligand 16 in epidermal keratinocytes to promote neutrophil aggregation (Sa et al., 2007; Steffen et al., 2018; Tohyama et al., 2012).
It also induces the maturation of myeloid dendritic cells, accelerates T cell proliferation, and stimulates T cell polarization into type 1 T helper or type 17 T helper cells (Ganguly et al., 2009). These activities account for the aggravation of psoriatic inflammation by IFN-α in mice treated with IMQ.
Epidermal renewal is an evolutionarily conserved process in which proliferative basal cells develop into anuclear keratinocytes. Under normal conditions, this process takes ∼13 days (Ogawa et al., 2018); however, this process occurs in only 48 h in psoriasis, indicating that keratinocytes in psoriatic lesions proliferate rapidly and abnormally produce keratins (Ogawa et al., 2018). Under normal conditions, the distribution of differentiation-specific keratins 1, keratin 10, and loricrin gradually increases above the spinous layer (Donetti et al., 2012). Conversely, psoriatic keratinocytes are abnormally differentiated and overproliferated, and the distribution of keratin 1, keratin 10, and loricrin above the spinous layer tends to be decreased, whereas that of keratin 17 gradually increases (Wang et al., 2022).
In the present set of experiments, treatment with IFN-α alone led to the formation of mild psoriatic lesions, and aberrant keratin expression was observed. The IFN-α and IMQ combination further strengthened the changes in keratins. These results indicate that the IFN-α and IMQ combination may produce a psoriatic mouse model that involves more severe abnormal keratin expression, which may be useful for some in vivo studies of psoriasis.
The immune response mediated by T cells, especially Th17 cells, is central to the pathogenesis of psoriasis. IL-17A, IL-22, IL-23, and TNF-α are the main pathogenic factors of psoriasis. In our study, IFN-α alone did not cause an increase in the protein levels of IL-17, IL-22, and IL-23. Its combination with IMQ produced a synergistic effect on these proteins, although the differences observed were not statistically significant. The relatively mild changes we detected may be a result of the timing of the single observation. In a study by van der Fits et al. (2009) the expression of IL-17 and IL-22 reached their highest levels on day 3 after IMQ treatment and then decreased rapidly. Here, levels were investigated on day 7 after initiation of IMQ treatment; subsequent experiments will involve the determination of these effects over a longer span of time.
FOXO3 belongs to the O subclass of the forkhead transcription factor family (Santos et al., 2023). The function of FOXO3 depends on its phosphorylation status and subcellular localization. The sites Ser253, Ser315, and Thr32 of FOXO3 can be phosphorylated by protein kinase B/c-AKT, resulting in the translocation of FOXO3 to the cytoplasm, where it is sequestered via interactions with 14-3-3. This phosphorylation thus reduces the level of FOXO3 in the nucleus and inhibits its transcriptional activity (Cao et al., 2023), resulting in inflammation and proliferation (Orea-Soufi et al., 2022). Previous studies have found that FOXO3 correlates with TNF-α in inflammation and proliferation (Brandstetter et al., 2019; Feng et al., 2021; Liu et al., 2017; Min et al., 2015). In our experiment, we observed that IFN-α promoted the inflammatory response and upregulated the expression of Ki67 and keratin 17.
Interestingly, we also found that IFN-α treatment increased levels of phosphorylated FOXO3 in the cytoplasm and decreased the levels of FOXO3 in the nucleus. Therefore, we propose that the development of psoriasis upon treatment with IFN-α involve the phosphorylation of FOXO3.
Conclusions
Our data suggested that IFN-α plays a critical role in the development of psoriasis, including in the abnormal differentiation and proliferation of keratinocytes and in the release of inflammatory factors. In addition, IFN-α aggravates psoriatic inflammation in mice when coadministered with IMQ, which may provide a useful new option for in vivo psoriasis research.
Footnotes
Acknowledgments
Thanks to the second affiliated hospital of Xi'an Jiaotong University Research Center that provided experimental platform and technical support.
Authors' Contributions
H.G.: conceptualization, data curation, formal analysis, investigation, methodology, validation, visualization, writing-original draft, and writing-review and editing. X.W.: methodology, supervision, and writing-review and editing. M.L.: formal analysis, investigation, and methodology. Y.W.: methodology and supervision. K.R.: investigation and formal analysis. Y.Z.: investigation and methodology. W.L.: investigation and resource. G.H.: funding acquisition and resources. W.Z.: conceptualization, project administration, and writing-review and editing. Y.X.: conceptualization, data curation, project administration, and writing-review and editing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Natural Science Foundation of Shaanxi Province (No. 2022JQ-976).
Supplementary Material
Supplementary Data S1
Supplementary Figure S1
Supplementary Table S1