
Abstract
Intestinal damage and secondary bacterial translocation are caused by the inflammatory response induced by sepsis. Tongfu Lifei (TLF) decoction has a protective effect on sepsis-related gastrointestinal function injury. However, the relation between gut microbiota, immune barrier, and sepsis under the treatment of TLF have not been well clarified yet. Here, rats were subjected to cecal ligation and puncture (CLP) to create a sepsis model. Subsequently, the TLF decoction was given to CLP rats by gavage, fecal microbiota transplantation (FMT), and antibiotic were used as positive control. TLF suppressed the inflammatory response and improved the pathological changes in the intestines of CLP rats. Besides, TLF promoted the balance of the percentage of the Th17 and Treg cells. Intestinal barrier function was also improved by TLF through enhancing ZO-1, and Occludin and Claudin 1 expression, preventing the secondary translocation of other gut microbiota. TLF dramatically boosted the gut microbiota's alpha- and beta-diversity in CLP rats. Moreover, it increased the relative abundance of anti-inflammatory gut microbiota and changed the progress of the glucose metabolism. In short, TLF regulated the gut microbiota to balance the ratio of Th17/Treg cells, reducing the inflammation in serum and intestinal mucosal injury in rats.
Introduction
A
In recent years, the doctrine of intestinal bacterial/bacterial endotoxin lipopolysaccharide (LPS) translocation has received increasing attention in the mechanism of sepsis (Panpetch et al., 2020; Wang et al., 2019). Patients with sepsis exhibit a dysfunctional intestinal microbiota, which increases the intestinal epithelial cell barrier's permeability (Shen et al., 2014). A large number of bacteria and LPS enter the blood circulation via the portal system and the mesenteric lymphatic system, leading to the uncontrolled regulation of the inflammatory and immune response of the organism (Huang and Chen, 2016).
Regulation of the Th17/Treg cells balance may have an important role in the protection of the intestinal mucosal immune barrier (Shi et al., 2022). It has been reported that when the intestinal microecology is disturbed, the gut microbiota co-metabolizes with the host and its co-metabolite stimulation can trigger the organism to react and immune response, leading to the production of pro-inflammatory cytokines and causing a tilt of the Th17/Treg balance toward Th17 cells (Hansson, 2012). Therefore, it is very important to study the mechanism of sepsis and drug development from the intestinal mucosal immune barrier (Vancamelbeke and Vermeire, 2017).
Sepsis can be classified as a “warm disease” based on the early manifestations of sepsis characterized by exogenous fever which is consistent with modern medical research that confirms that the displacement of bacteria and toxins in the gastrointestinal tract due to intestinal failure is a major reason for severe sepsis and MODS (Kernbauer et al., 2015).
Tongfu Lifei (TLF) is a kind of traditional Chinese medicine, which exerts protective treatment of sepsis in clinical practice (Lv et al., 2018). In terms of therapeutic effects, it has been shown that high mobility group box-1 and interleukin-10 (IL-10) levels can be considerably reduced and elevated by TLF decoction colon dialysis combined with fiberoptic bronchoscopy alveolar lavage, which inhibits the release of pro-inflammatory molecules, suppressing excessive inflammatory responses (Yang, 2018). The treatment that cleaning the bowels through TLF can improve gastrointestinal dysfunction and promote the recovery of patients. The mechanism is that it can effectively protect the intestinal mucosa, decreasing intestinal permeability, and reducing inflammatory mediators. Our group found that TLF decoction can protect the intestinal barrier damage with sepsis, improve intestinal mucosal villi damage through upregulating ZO-1/occludin/claudin-1 expression (Chen et al., 2020).
Therefore, based on the above work, we hypothesized that TLF can regulate the balance of gut microbiota in sepsis, intervening in the state of intestine-bacteria immune interaction. Subsequently, it adjusts the balance of Th17/Treg immune axis, the key target of intestine-bacteria immune interaction, restoring the immune response in the intestine, and achieving the protective effect on the intestinal mucosal immune barrier protection of sepsis.
Materials and Methods
Ethics
The animal experiments of the study were permitted by the Animal Experimentation Ethics Committee of Zhejiang Eyong Pharmaceutical Research and Development Center (SYXK (Zhe) 2021-0033) and adhered to the guidelines of the China Council on Animal Care and Use.
Animals and experiments
Thirty Sprague-Dawley rats, male and female for half, weighing 250–300 g, were supplied from Zhejiang Weitong Lihua Laboratory Animal Technology Co., LTD., Animal Production License No. SCXK (Zhe) 2019-0001. They lived at 24°C–26°C and relative humidity of 45%–60%. The animal research was ratified by the Ethics Committee of the Animal Center of Zhejiang Eyong Pharmaceutical Research and Development Center. Animal use license number: SYXK (Zhe) 2021-0033. The rats were fed adaptively for 7 days before undergoing cecal ligation and puncture (CLP) surgery and were fasted for 6 h before the experiment.
Establishment of sepsis model
According to the previous study (Chen et al., 2021), rats received CLP as the sepsis model. They were anesthetized with isoflurane, afterward, a median incision was made in the abdominal wall, and the cecum was gently pulled out to protect the mesenteric blood vessels. To fill the cecal ends and divide the mesenteric surface blood vessels, the feces at the top end of the cecum were gently pressed. A sterile needle was used to puncture the cecum wall at the midpoint between the ligation site and the apex of the cecum to create perforation. A sterile No. 4 silk thread was ligated at the midpoint between the blind valve and the cecum. The cecum was gently squeezed to allow the extrusion of a little content so that the perforation was unobvious. After the extruded content was wiped out, the cecum was pushed back into the abdominal cavity, and layers of sutures were applied. After the operation, saline was used for fluid resuscitation (37°C, 5 mL/100 g) and buprenorphine (0.05 mg/kg) was used for pain relief.
Rats in sham group were treated with abdominal wall opening without CLP, and then the abdominal cavity was sutured.
TFL decoction preparation
Mirabilite, Magnolia officinalis, Forsythia, Scutellaria, Almond, Bletilla, and Panax notoginseng each for 10 g, and Rhubarb 20 g. After these medicines were immersed in water for 2 h, the water was decocted and concentrated. After disinfection, the decoction is refrigerated for later use.
Fecal filtrate preparation
The same batch of healthy rats were selected as fecal donors for the fecal microbial transplantation experiment. Under aseptic conditions, feces were collected from donor rats every day under laminar flow hood. Feces from rats in the donor group were pooled, and 100 mg feces were dissolved in 1 mL sterile normal saline. The solution was vigorously mixed using a bench swirl for 10 s, and the supernatant was collected by centrifugation and used as donor graft material for fecal microbial graft experiments.
Group administration
Each group contains 6 rats. All drugs were prepared by dissolving in distilled water. Sham group rats were given normal saline 10 mL/kg by gavage. Model group rats were given CLP, and normal saline 10 mL/kg was given intragastric administration. CLP+TLF group: 7.2 g/kg TLF decoction was given to CLP rats by gavage. CLP+fecal microbial transplantation (FMT) group: each CLP rat was given fresh diluted fecal filtrate 10 mL/kg by gavage. CLP+antibiotic group: antibiotics were given to CLP rats by gavage (4 mixtures of ampicillin, neomycin sulfate and metronidazole for 1 g/L vancomycin for 0.5 g/L), dissolved in water at this concentration, and administered through drinking water. They were all treated once a day for a week.
Sample collection
One week after gavage, the fresh feces were collected, and then the rats were anesthetized with isoflurane. Two milliliters blood from the abdominal aorta. The serum was separated after centrifugation and stored at −20°C away from light. The fecal sample was stored in a sterile centrifuge tube and about 2–3 mL of each tube was then transferred into the collection tube, followed by being stored in the refrigerator at −80°C. Ultimately, the rats were euthanized by CO2 inhalation, and the spleens were removed. Lymphocytes were separated from the spleens and centrifuged. The single-cell suspension of the spleen was detected by flow cytometry (FCM). About 2 cm of diseased colon tissue was cut at 1.5 cm of the upper anus, and then dissected longitudinally for cleaning with sterile saline, and 0.5 cm of intestinal tissue was cut for staining.
Enzyme-linked immunosorbent assay measurement
The serum levels of tissue IL-1β (MM-0047R1), tumor necrosis factor (TNF)-α (MM-0180R1), and IL-6 (MM-0190R1) were tested with ELISA kits from Meimian (Yancheng, China) following the manufacturer's instructions.
Hematoxylin-eosin staining
The intestinal tissues were quickly removed and fixed with 4% paraformaldehyde. The intestinal tissues were dehydrated by gradient ethanol and xylene, and then immersed in wax. The tissues were cut into 5 μm slices and affixed to the antipeeling slides. The slices were baked at 60°C for 12 h, dewaxed and hydrated by xylene and gradient ethanol, and stained with hematoxylin and eosin (H&E) staining (G1005; SERVICEBIO, Wuhan, China). At last, ethanol with low to high concentrations was added to dehydrate. Following the vitrification by xylene, the slices were sealed with neutral balsam and were observed under the microscope (Nikon Eclipse E100).
Inflammatory cell infiltration scoring system was applied according to the previous study (Zhang et al., 2021), specifically no inflammation or lesion, score 0. Minimal inflammatory cell infiltration, lesion limited to the submucosa, score 1. Mild inflammatory cell infiltration, lesion extending to the muscular layer with crypt disruption, score 2. Significant inflammatory cell infiltration, lesion involving the muscular layer with evident crypt destruction, score 3. Extensive inflammatory cell infiltration, lesion reaching the serosal layer, with destruction of crypts and epithelium, score 4.
Western blot
Pure protein was extracted from the intestinal tissues, and the protein concentration was measured by the bicinchoninic acid method. After the loading buffer was added, the protein was boiled to achieve denaturation. The total protein was separated by electrophoresis, and the corresponding protein was transferred to the polyvinylidene difluoride membrane. The nonspecific antigen was blocked with 5% milk, and the protein on the membrane was incubated with the target antibody against Occludin (DF7504; Affinity, Cincinnati, OH), Claudin 1(AF0127; Affinity), ZO-1(21773-1-AP; proteintech, Wuhan, China), and β-actin (AF7018; Affinity). After incubation at 4°C overnight, it was incubated with secondary antibodies (Anti-rabbit IgG, HRP-linked Antibody, CST, 7074). Unbound antibodies were washed and captured in an enhanced chemiluminescence luminescence imager.
Quantitative real-time polymerase chain reaction
Pure inferior vena tissue RNA was obtained via the extraction using Trizol (B511311; Sangon Biotech, Shanghai, China) and transcribed into cDNA with a reverse transcription kit (CW2569; Jiangsu Cowin Biotech, Taizhou, China). Primers, DEPC, cDNA, and SYBR Green (RR820A; Takara, Shiga, Japan) were added to prepare the corresponding system for amplification products in the PCR instrument. The sequences of the primer are listed (Table 1). The fold changes of mRNA were calculated using the 2−ΔΔCT method.
Primer Sequence
Flow cytometry
The spleen and intestine tissues were mechanically separated, and then the red blood cell lysis buffer was used for lysis. Single cell suspensions were centrifuged for 5 min at 1,200 rpm after being passed through nylon membranes. For use in a FCM test, the suspension was resuspended in RPMI 1,640 containing 10% FBS. 20 μL CD3-APC (AR00305; MultiSciences Biotech, Hangzhou, China), 20 μL CD4-BV421 (743088; BD Biosciences Pharmingen, San Diego, CA), and 20 μL IL-17A-PE (AM0I1704; MultiSciences Biotech, China) was added to each tube (cell density at 106), followed by the rinse in Hank's buffer and incubation in a refrigerator at 4°C for 15 min in darkness. Twenty microliters Foxp3-PE (320011; Biolegend, San Diego, CA) was then added before the FCM assay. For the FCM assay, the quantity of CD4+IL-17+ Th17 cells was identified. Anti-CD4, anti-CD3, and anti-Foxp3 antibodies were coupled with fluorescein isothiocyanate, allophycocyanin, and phycoerythrin to stain lymphocytes. Using the FCM assay, the number of CD4+, CD3+, and Foxp3+ Treg cells was selected.
16S rDNA sequencing and analysis
Each fecal group's whole genomic DNA was extracted, and target sequences (the 16S rDNA V3-V4 region) were amplified by PCR. To create a sequencing library, the products were then cleaned, measured, and homogenized. For building libraries, a library quality assessment was done, and qualified libraries were sequenced on Illumina HiSeq 2500 (Illumina, San Diego, CA). Base-calling analysis was used to turn the original picture data files obtained during high-throughput sequencing into sequenced reads. The results were stored in FASTQ format file for further bioinformatics analysis. The Alpha diversity level of each group was evaluated according to the distribution of Amplicon Sequence Variants/Operational Taxonomic Units in different groups. Each group's distance matrix was constructed and using a number of unsupervised sequencing and clustering techniques along with associated statistical testing techniques, the differences in beta diversity and species abundance composition between various groups were determined. Based on the results of gene sequencing, the metabolic function of the sample flora was predicted, and the species composition of the specific pathway was obtained.
Statistical analysis
The statistical analysis was carried out using SPSS software (16.0; IBM, Armonk, NY). One-way analysis of variance was used to analyze the data under normal distribution in this study. Dunnett's T3 test or independent sample t-test was used if the distribution was normal, but the variance was not uniform. If it did not conform to the normal distribution, the Kruskal–Wallis H test was used. P < 0.05 was designated as statistically significant.
Results
TLF improved the pathological changes of intestinal mucosa in CLP rats
As can be seen from Fig. 1, the intestinal tissue structure of the sham group is complete, and the hierarchy is clear. Compared with the sham group, the CLP group showed a large number of inflammatory cell infiltration, and the lesions invaded the serosa layer. CLP+TLF group and CLP+FMT group had a small amount of inflammatory cell infiltration, and few intestinal villi were destroyed. The pathological damage of intestinal tissue in rats of the CLP+antibiotic group is lighter than the CLP group. In addition, the semiquantitative H&E score in the CLP group significantly increased compared to the control group (P < 0.01). Compared to the CLP group, H&E scores in the CLP+TLF group, CLP+FMT group, and CLP+antibiotic group significantly decreased (P < 0.01). Compared to the CLP+TLF group, H&E score in the CLP+antibiotic group significantly decreased (P < 0.01), and compared to the CLP+FMT group, H&E score in the CLP+antibiotic group significantly decreased (P < 0.05).
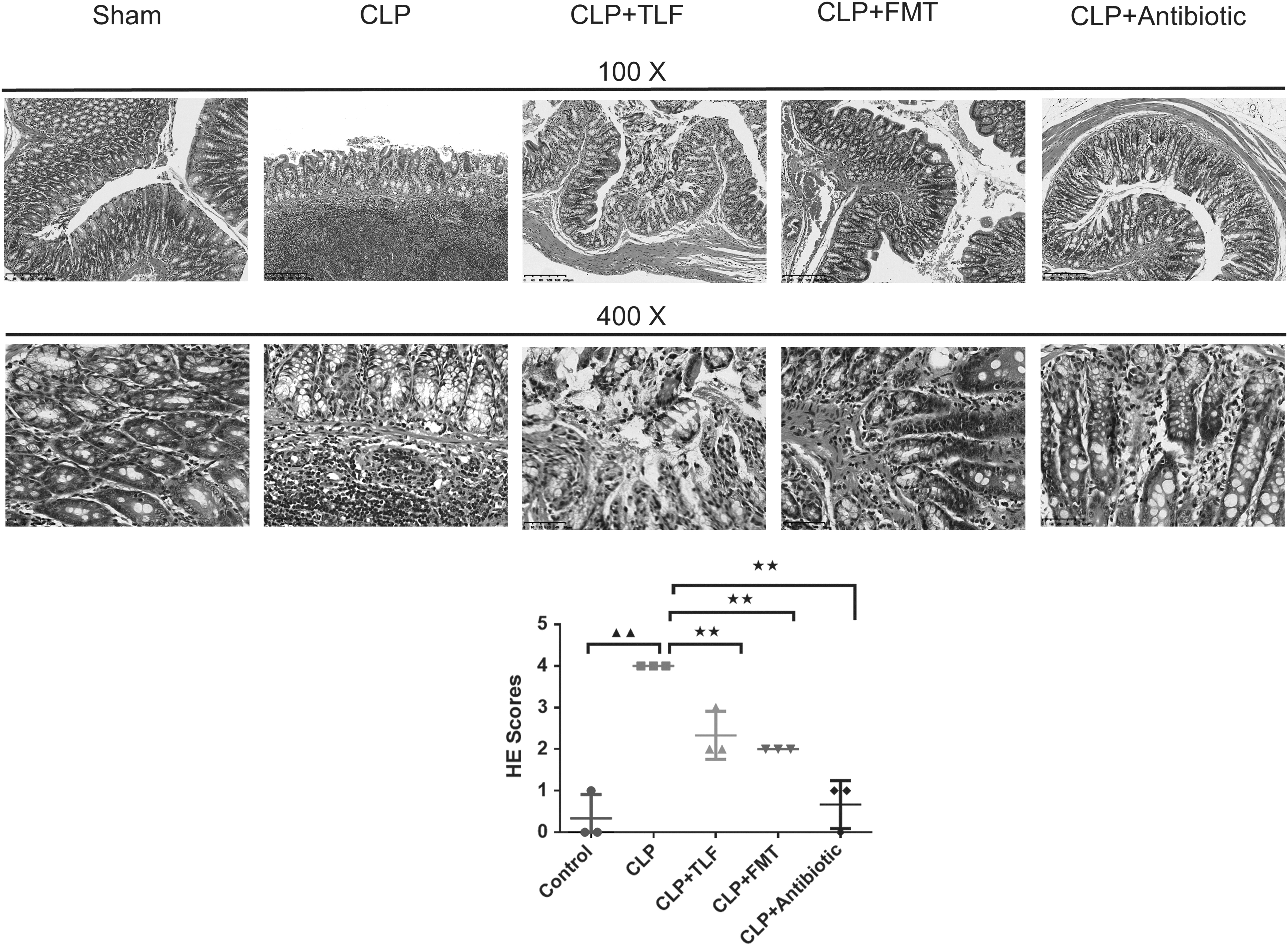
TLF decoction improved the pathological changes of intestinal mucosa in CLP rats. The histomorphology of intestinal tissue in rats was observed by H&E staining and evaluated by the H&E score (magnification × 100, 400) ▲▲ P < 0.01 versus sham group. ★★ P < 0.01 versus CLP group. CLP+TLF: cecal ligation and perforation rats with the treatment of TLF decoction, CLP+FMT: cecal ligation and perforation rats with the treatment of fecal microbial transplantation, CLP+Antibiotic: cecal ligation and perforation rats with the treatment of antibiotic. CLP, cecal ligation and puncture; FMT, fecal microbiota transplantation; H&E, hematoxylin and eosin; TLF, Tongfu Lifei.
TLF exerted an anti-inflammatory effect on the CLP rats
In Figure 2, ELISA kits were used to test the content of inflammatory factors. Compared with the sham group, the contents of TNF-α, IL-1β, and IL-6 in the serum of rats in the CLP group were significantly increased (P < 0.01). With the interference of TLF, FMT, and antibiotic, respectively, the levels of TNF-α, IL-1β, and IL-6 in the serum of rats were lower than CLP rats (P < 0.05 or P < 0.01).
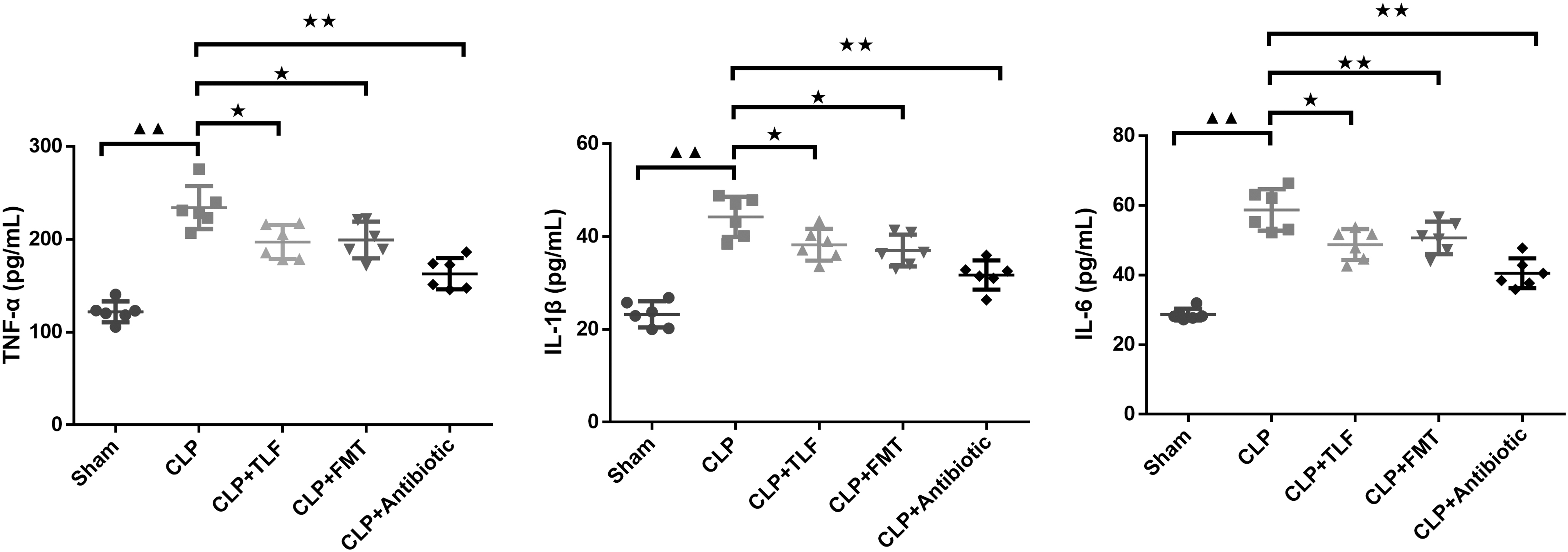
TLF exerted anti-inflammatory effect in serum of the CLP rats. ELISA was used to test TNF-α, IL-1β, and IL-6 levels in each group, n = 6 in each group; ▲▲ P < 0.01 versus sham group. ★ P < 0.05, ★★ P < 0.01 versus CLP group. CLP: cecal ligation and perforation rats, CLP+TLF: cecal ligation and perforation rats with the treatment of TLF decoction, CLP+FMT: cecal ligation and perforation rats with the treatment of fecal microbial transplantation, CLP+Antibiotic: cecal ligation and perforation rats with the treatment of antibiotic. ELISA, enzyme-linked immunosorbent assay; IL, interleukin TNF, tumor necrosis factor.
The effect of TLF on the intestinal barrier of the intestinal in CLP rats
To explore the effect of TLF on the intestinal barrier, quantitative real-time polymerase chain reaction (qRT-PCR) and western blot were used to observe the intestinal barrier-related factors. In Fig. 3A, the relative mRNA expressions of ZO-1, Claudin 1, and Occludin in CLP rats were decreased compared to the sham group. With the interference of TLF, FMT, and antibiotic, respectively, the expressions of intestinal barrier-related factors were strengthened compared to the CLP group (P < 0.01). As expected, in Fig. 3B, the results of western blot that the expression trend of ZO-1, Claudin 1, and Occludin demonstrated the same way as qRT-PCR observed (P < 0.05 or P < 0.01).

TLF enhanced the expression of ZO-1, Claudin 1 and Occludin proteins associated with intestinal mucosa in CLP rats.
TLF promotes the balance of the Th17/Treg cells in the lymphocyte of the CLP rats
To clarify whether TLF will impact the balance of immune cells in CLP rats, we used the FCM to observe the percentage of the Th17/Treg cells in spleen and intestine. In Fig. 4A, compared with the sham group, Th17 and Treg cells of the CLP group were significantly increased which were imbalanced (P < 0.01). Compared with CLP group, the number of Th17 cells in splenic cells of CLP+TLF group, CLP+FMT group, and CLP+antibiotic group was decreased (P < 0.05 or P < 0.01), while the number of Treg cells was enhanced (P < 0.05 or P < 0.01), contributing to the balance situation in Th17/Treg cells in the lymphocyte. In Fig. 4B, compared to the sham group, the number of Th17 cells in intestinal cells of the CLP group significantly increased (P < 0.01), and Treg cells significantly decreased (P < 0.01). Compared to the CLP group, the number of Th17 cells in intestinal cells of the CLP+TLF group, CLP+FMT group, and CLP+antibiotic group significantly decreased (P < 0.01), while the number of Treg cells significantly increased (P < 0.01).
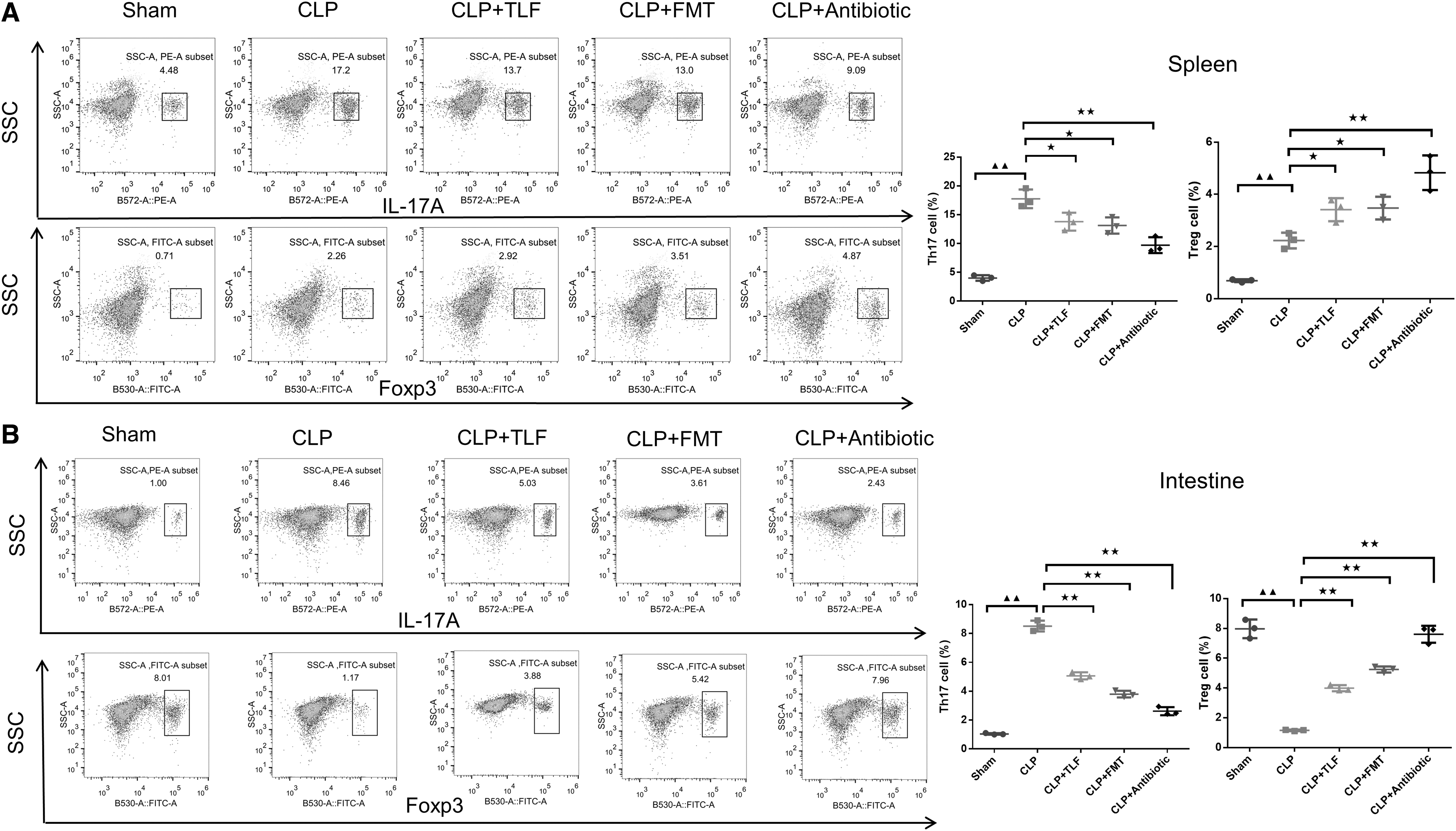
TLF promote the balance of the Th17/Treg cells in the lymphocyte of the CLP rats.
The effect of TLF on the diversity of gut microbiota in CLP rats
A gut analysis was done on CLP rats to investigate the effect of CLP and TLF interactions on the gut microbiota. According to the findings in Fig. 5A, the CLP group's gut microbiota's alpha diversity was lower than that of the Sham group (P < 0.01) in terms of the alpha diversity indices (Chao1, Simpson, and Shannon). Naturally, the TLF group had higher alpha diversity indices. Antibiotic therapy worsened the loss of alpha diversity in CLP rats. We also looked at the beta diversity of the gut microbiota (variation of microbial composition in groups) in Fig. 5B, C. It's interesting to note that the CLP group's gut microbiome differed from that of the other four groups, and that of the CLP+TLF group was more like that of the sham group than the CLP group.

The effect of TLF on the diversity of gut microbiota in CLP rats.
The moderating effect of TLF on gut microbiota in CLP rats
The gut microbiota showed abundance differences in all groups in the phylum and genus level. The top 20 phyla and genera with the highest average ASV/OTU frequency in the groups were displayed, respectively (Fig. 6A, B). Importantly, TLF upregulated the relative abundance of Oscillospira, while Blautia was decreased in the TLF than the CLP group, and paraprevotella was decreased in CLP rats compared to the sham group (Fig. 6C, P < 0.05 or P < 0.01).
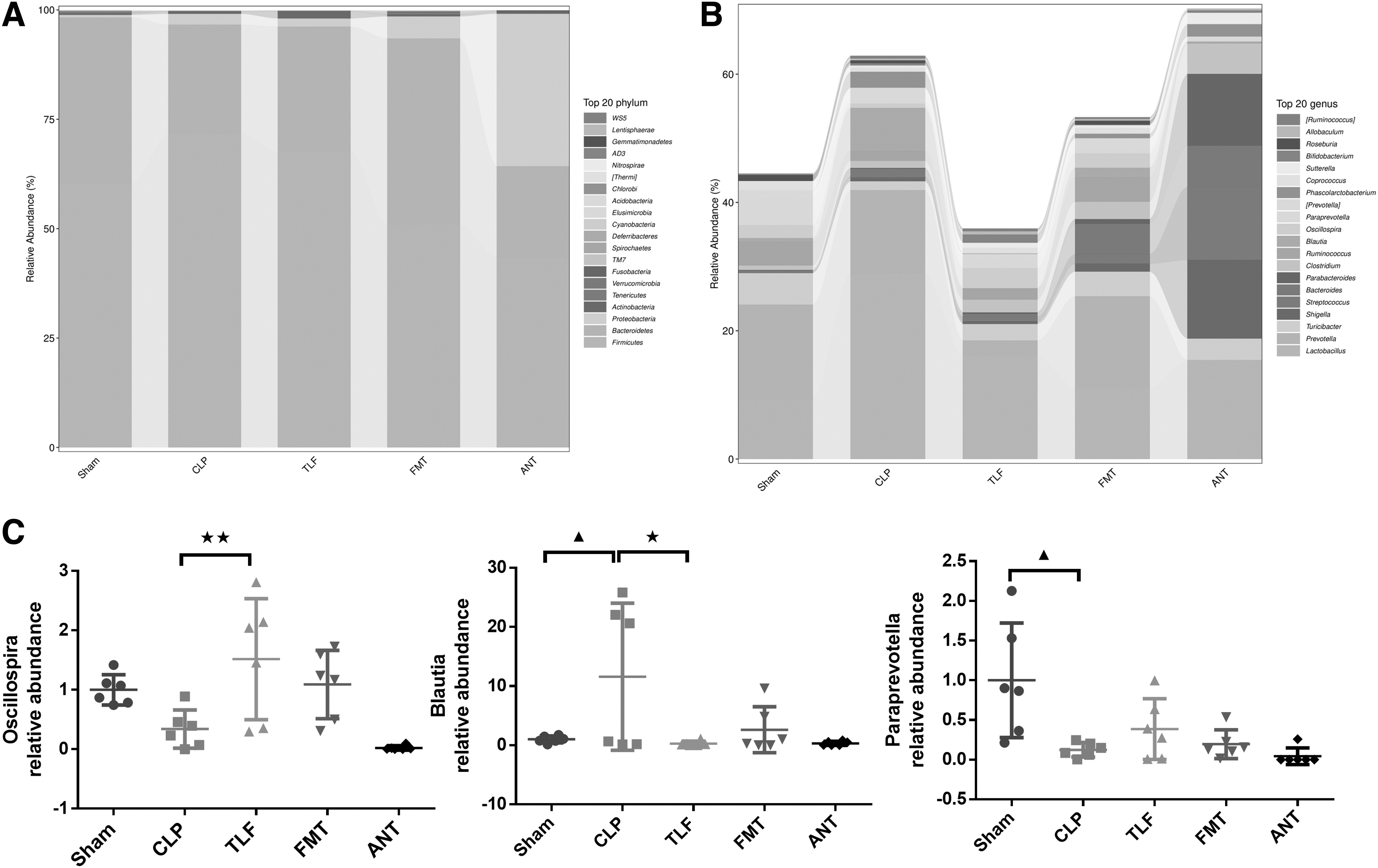
The beneficial effect of TLF on gut microbiota in CLP rats.
We further used the LEfSe to analyze the degree of influence of significantly different species in groups. In Fig. 7A, B, we can see different significant species of gut microbiota in each group. In the TLF group, the LDA scores of the Clostridia and Clostridiales Oscillospira were high. The colored solid spot showed that it influenced the variation of the gut microbiota, especially with Clostridia, Clostridiales, and Lachnospiraceae.
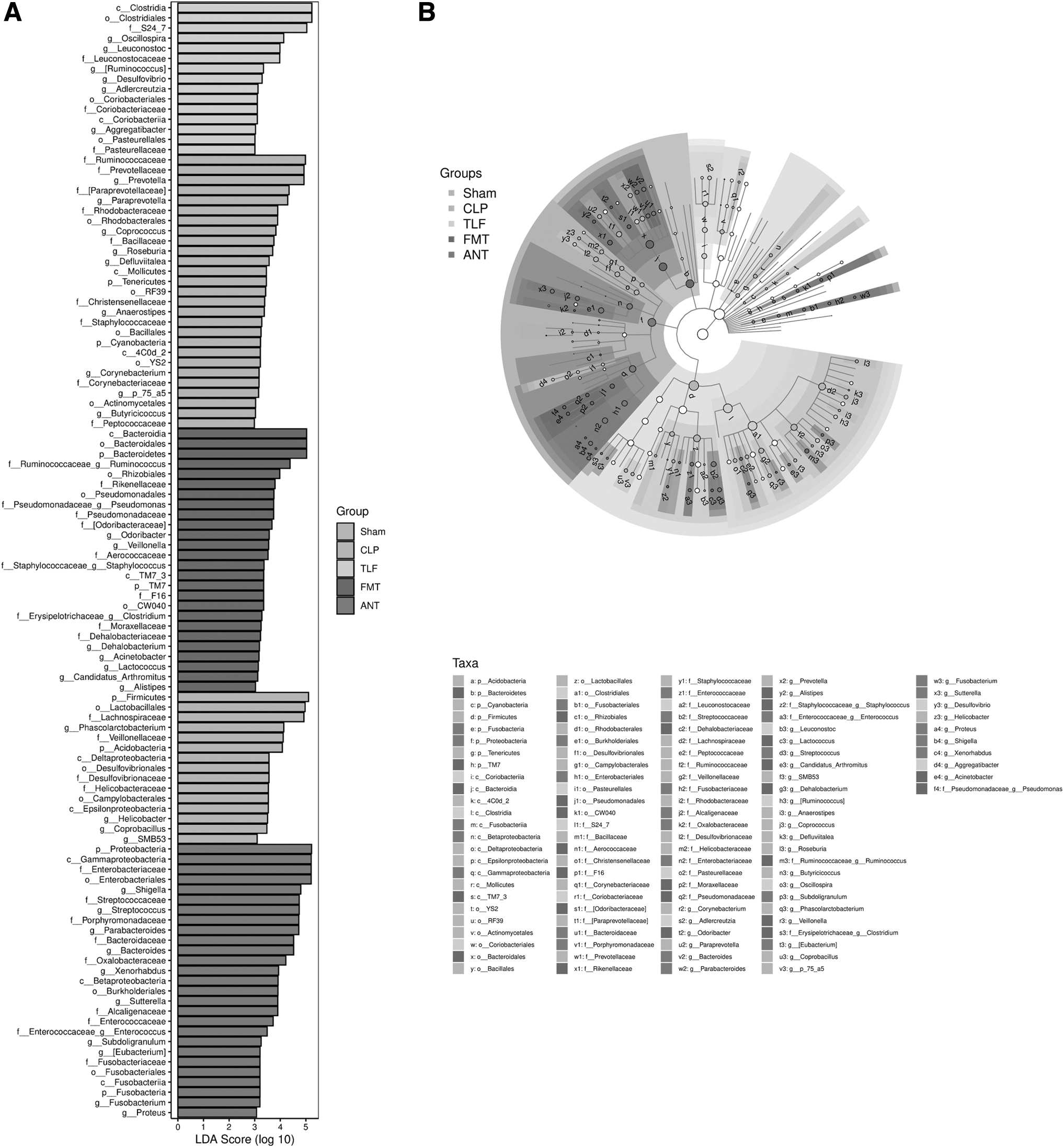
The variation analysis of gut microbiota after the TLF in CLP rats.
In Figure 8A, B, we can see that from both the glycolysis and glycogen biosynthesis process, the treatment of the TLF restored the relative abundance of the gut microbiota.
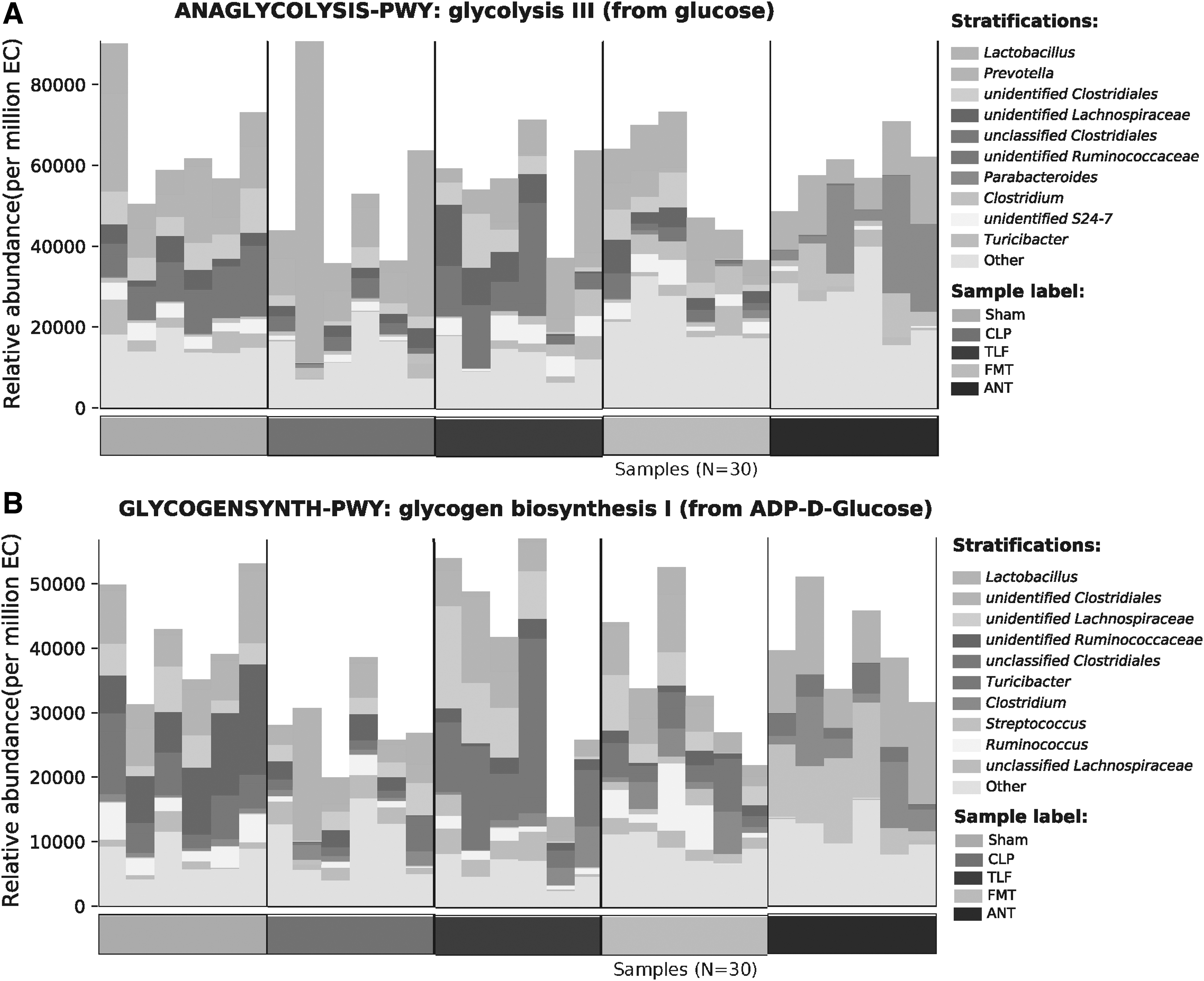
The beneficial effect of TLF on glycometabolism in CLP rats.
Discussion
Sepsis is a dangerous disease, which can lead to organ dysfunction. The gastrointestinal system has been shown to have a significant role in the pathophysiology of sepsis (Wang et al., 2019). In an infectious and inflammatory situation, it is possible that with high permeability of intestinal bacteria, endotoxin and a variety of metabolites from the intestinal transfer to the location of the previous sterile, aggravating inflammation and immune response leading to serious sepsis (Cavaillon, 2018; Hu et al., 2019). It has been reported that an intact intestinal barrier can reduce inflammation and immune response caused by sepsis (Adelman et al., 2020; Ma et al., 2020). Meanwhile, through previous studies, TLF can play a role in reducing sepsis. As a result, in this study, we further explored whether TLF decoction attenuates sepsis-related intestinal mucosal injury by regulating Th17/Treg balance in modulating the gut microbiota of rats.
The relationship between sepsis, dysregulated potassium levels (hypokalemia or hyperkalemia), and gut microbiota is multifaceted (Suri et al., 1997). Sepsis can disrupt potassium balance, leading to either hypokalemia or hyperkalemia (Shih et al., 2012). Factors contributing to hypokalemia in sepsis include diuretic use, increased renal potassium loss, and hormonal shifts causing potassium to move into cells. Hyperkalemia in sepsis may result from tissue breakdown, metabolic acidosis, and impaired renal function. While gut microbiota influences the body's metabolism and electrolyte balance, its direct impact on potassium levels in sepsis is less clear and requires further research. The complex interplay of immune responses, hormonal changes, and renal function in sepsis affects both potassium balance and gut microbiota composition. It is commonly acknowledged that TNF-α, IL-1β, and IL-6 are the major inflammatory biomarkers in sepsis (Deng et al., 2022). With less inflammatory cell infiltration, fewer TNF-α, IL-1β, and IL-6 content in serum and less intestinal tissue destruction under the treatment of the TLF in our study, we concluded that TLF exerted a powerful protective effect on the sepsis-induced inflammation and tissue injury. In addition, TLF decoction has been researched in sepsis with the protection of the intestinal barrier. According to previous studies (Chen et al., 2021; Chen et al., 2020), it was observed that TLF protected the intestinal barrier of sepsis in rats through regulating ZO-1/Occludin/claudin-1, and TLF, to some extent, engaging in the immune response to alleviate the intestinal barrier injury caused by the sepsis.
It is believed that the balance of the Th17/Treg is critical to the immune system in sepsis disease. Sepsis patients had significantly higher Treg cells and lower Th17 cells in comparison to nonsepsis (Gupta et al., 2016). On the contrary, the results we concluded that the ratio of Th17/Treg in CLP model was higher than sham group, and after the treatment of TLF, FMT, antibiotic, respectively, the ratio of Th17/Treg was reduced than CLP model which was the same as Cheng's study (Cheng et al., 2022). The apparent inconsistency can be explained through time-dependent changes, differences in local versus systemic immune response, and impact of experimental conditions factors. TLF contributed to the upregulation of the expression of ZO-1/Occludin/Claudin-1 which maintained the integrity of the intestinal barrier and suppressed the leaky gut. The balance percentage of the Th17/Treg and the less inflammatory factors produced indicate that TLF may prevent the process of sepsis that less gut microbiota transferred to the sterile places, causing the less inflammatory and immune response.
A study confirmed that in patients intestinal dysbiosis and sepsis are related (Liu et al., 2019). Ye (Ye et al., 2021) in their research found that gut microbiota changes are brought on by chlorogenic acid enhanced metabolic endotoxemia. It is believed that TLF affects sepsis by modulating the microbiome at least in part. Our study showed that TLF dramatically boosted the alpha- and beta-diversity of gut microbiota in CLP rats based on 16s rDNA gene sequencing. A study (Jiao et al., 2018) showed that Oscillospira, which turns polysaccharides into short-chain fatty acids (SCFAs) and promotes insulin resistance and systemic inflammation, was found in greater abundance in obese rodents. Liang (Liang et al., 2022) in their study mentioned and revealed that metformin treatment enhanced the relative abundance of bacteria that offer intestinal cells energy, maintain the gut barrier, and keep LPS from crossing the intestinal wall, including Bifidobacterium, Muribaculaceae, Parabacteroides-distasonis, and Alloprevitella.
Consistent with their study, in our study, we observed that the treatment of the TLF increased the relative abundance of Oscillospira which exerted an anti-inflammatory effect on sepsis. Besides, the relative abundance of Bifidobacterium was strengthened compared with the CLP group. These bacteria produce SCFA, which provides energy to intestinal cells and maintains the gut barrier. Blautia prevents pathogens colonization by producing bacteriocins that exhibit anti-inflammatory properties and maintain glucose homeostasis via upregulating the production of Treg cells and SCFA (Liu et al., 2021; Palmieri et al., 2021). However, in our study, Blautia had a great decrease after the treatment of the TLF that may be related to the balance of Th17/Treg cells.
To further explore whether the gut microbiota species participate in the metabolic process, we specifically focus on glycometabolism. Since studies confirmed that abnormal glucose metabolism during sepsis is related to metabolic hormones and inflammatory cytokines. A large number of cytokines IL-6 and TNF-a can affect glucose metabolism by stimulating the secretion of hyperglycemia, cortisol, and other hormones in the body (Owolabi et al., 2021); on the other hand, they can also affect glucose metabolism through direct effects. In Masataka Nakamura's study (Nakamura et al., 2012), they revealed that high blood IL-6 level was correlated with hyperglycemia in septic patients. In our research, we found that from both glycolysis and glycogen biosynthesis processes, the treatment of the TLF restored the relative abundance of the gut microbiota and ameliorated the inflammation.
In this study, we found that TLF reduced the expression of serum inflammatory factors, restored the impaired intestinal tissues and intestinal barrier, increased the diversity of gut microbiota, and balanced the ratio of Th17/Treg cells, thus exerting an ameliorating effect on the sepsis rats. However, deeper studies are needed to explore the related bacteria in alleviating the sepsis disease, and the study results need to be confirmed by other similar studies.
Conclusion
To sum up, TLF regulated the gut microbiota to balance the ratio of Th17/Treg cells, reducing the inflammation response and intestinal mucosal injury in sepsis rats. This study provided the basis for further clinical and experimental research on TLF's treatment of sepsis.
Footnotes
Authors' Contributions
Conceptualization: H.C.; Funding acquisition: H.C.; Data curation: H.C., Z.Y., Z.Q., X.H., J.G.; Roles/Writing—original draft: H.C., J.G.; Writing—review and editing: J.G.; Final approval of the version to be published: All authors. Agreement to be accountable for all aspects of the work to ensure questions related to the accuracy and integrity of the work are appropriately investigated and resolved: H.C., Z.Y., Z.Q., X.H., J.G.
Data Availability Statement
Data will be made available on request.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the “Hangzhou Science and Technology Development Plan Project” (grant numbers 20201203B181).